Association entre les polymorphismes et
l’expression du gène de la leptine et la qualité de la
viande et de la carcasse chez l’agneau lourd
Mémoire
Vincent Brodeur
Maîtrise en sciences animales
Maître ès sciences (M. Sc.)
Québec, Canada
iii
L’étude faisant l’objet de ce mémoire avait pour but d’évaluer les effets des caractéristiques moléculaires du gène de la leptine ovine, une hormone impliquée dans le contrôle des réserves énergétiques de l’organisme et de la satiété, sur les caractères liés à la qualité de la viande et de la carcasse dans une population d’agneaux lourds provenant du Québec. La population à l’étude était composée de 128 individus (½mâles, ½femelles) provenant de deux génotypes : ½Suffolk ¼Romanov ¼Dorset (xSU) et ½Arcott Canadien ¼Romanov ¼Dorset (xCD).
Le gène LEP a été séquencé. Le gène a ensuite été séquencé pour chaque individu et ses parents, entre l’exon 2 et l’exon 3, afin d’y chercher des variations dans un segment d’ADN de 2328 paires de bases. Vingt-trois polymorphismes ont été recensés; de ceux-ci, 12 ont été étudiés et ont montré des effets sur la qualité de la viande et de la carcasse.
L’expression du gène LEP a été mesurée dans le tissu adipeux sous cutané, à l’aide de la méthode du PCR en temps réel. Des corrélations significatives ont été établies entre l’activité du gène et des variations de paramètres liés à la couleur de la viande chez les agneaux mâles xCD. L’épaisseur du muscle mesurée aux ultrasons chez ce même les animaux de ce même croisement était positivement liée à l’activité du gène.
Des corrélations ont aussi été établies entre la concentration de la leptine dans le sang, l’expression du gène LEP et quelques caractères liés au rendement en maigre du muscle longissimus dorsi de même qu’à la couleur de la viande.
Les résultats du projet offrent des pistes pouvant mener au développement de marqueurs génétiques qui pourraient aider dans la sélection des ovins reproducteurs, dans le but d’améliorer la qualité de la carcasse et de la viande des agneaux produits au Québec.
iv
Abstract
The purpose of this study was to evaluate the effects of the molecular characteristics of the ovine leptin gene (LEP) on characteristics related to meat and carcass quality in a lamb population from the Province of Quebec. The studied population consisted of 128 individuals (½ males, ½ females) from 2 crossbreeds : ½Suffolk ¼Romanov x¼ Dorset (xSU) and ½Arcott Canadian ¼Romanov ¼Dorset (xCD).
Sequencing of the LEP gene was performed. The gene was then sequenced for each individual and its parents, between exon 2 and exon 3, in order to search for variations within a 2328 base pairs DNA strand. Twenty-three polymorphisms were identified; 12 have been the subject of further studies and have shown interesting effects on meat and carcass quality.
Expression of the LEP gene was measured in subcutaneous adipose tissue using the real-time PCR method. Significant correlations were established between gene activity and variations in meat color parameters in male lambs of the xCD crossbreed. The thickness of the muscle measured with ultrasound in this same crossbreed was positively related to the activity of the LEP gene.
Correlations were also established between leptin concentration in blood, LEP gene expression, and a few characteristics related to lean yield of the longissimus dorsi muscle as well as the color of the meat.
The study of the sheep leptin gene, as part of this project, offers some avenues that could lead to the development of genetic markers that could help in the selection of breeding sheep to improve the meat and carcass quality of lamb produced in Québec.
v
Table des matières
Résumé ... iii
Abstract ... iv
Table des matières ... v
Liste des tableaux ... vi
Liste des figures ... vii
Liste des abréviations ... viii
Remerciements ... xi
Avant-propos ... xiii
Chapitre 1 : Introduction ... 1
Chapitre 2 : Revue des travaux antérieurs ... 4
2.1. Régulation du poids corporel ... 4
2.2. Le tissu adipeux ... 9
2.3. La leptine : généralités ... 13
2.3.1. Travaux préliminaires ... 13
2.3.2. Caractérisation ... 15
2.3.3. Sites d’expression de la leptine ... 20
2.3.4. Le récepteur de la leptine et ses sites d’expression ... 20
2.3.5. Fonctions physiologiques de la leptine ... 26
2.3.5.1 Régulation de la prise alimentaire et métabolisme lipidique ... 28
2.3.5.2 Métabolisme et croissance musculaire ... 30
2.3.5.3 Reproduction ... 33
2.3.5.4 Hématopoïèse et immunité cellulaire ... 38
2.3.5.5 Angiogenèse ... 40
2.3.5.6 Métabolisme osseux ... 43
2.4 Caractérisation de la leptine ovine ... 47
2.5 Polymorphismes du gène de la leptine chez différentes espèces d’intérêt agronomique : effets physiologiques ... 55
2.5.1. Le bovin ... 55
2.5.2. Le porc ... 64
2.5.3. Le mouton ... 68
2.6 Hypothèse et objectifs ... 71
Chapitre 3 : Article scientifique ... 72
3.1 Résumé de l’article ... 72
3.2 Article scientifique ... 73
Leptin molecular characteristics affect carcass and meat quality in crossbred lambs ... 73
Abstract ... 74
Introduction ... 74
Material and Methods ... 76
Results and Discussion ... 81
Implications ... 89
Chapitre 4 : Discussion ... 97
Chapitre 5 : Conclusions générales ... 105
Liste des ouvrages cités ... 108
vi
Liste des tableaux
Revue des travaux antérieurs
Tableau 2.1 : Polymorphisme C305T du gène de la leptine bovine présent dans l’exon 2 du gène ... 58 Tableau 2.2 : Polymorphismes identifiés dans le promoteur du gène de la leptine
bovine ... 63 Tableau 2.3 : Polymorphismes du gène de la leptine porcine ... 65 Tableau 2.4 : Polymorphismes du gène de la leptine ovine ... 70 Article scientifique
Table 3.1 : Characteristics of lambs used in experimentation ... 91 Table 3.2 : Allelic frequencies of ovine leptin gene single nucleotide polymorphisms
... 92 Table 3.3 : Additional leptin gene polymorphisms ... 92 Table 3.4 : Effect of G1958A polymorphism of ovine leptin gene on carcass
composition, growth, meat quality traits and leptin concentration in blood
(associated with xSU) ... 93 Table 3.6 : Correlations between concentration of leptin in blood and carcass
composition, growth, and quality traits of meat in crossbreed lambs. ... 95 Table 3.7 : Correlations between expression of LEP gene and carcass composition,
vii
Liste des figures
Figure 2.1 : Éléments clés de l’équilique énergétique de l’organisme. ... 5
Figure 2.2 : Hypothèse du point défini ou Set Point Hypothesis. ... 8
Figure 2.3 : Facteurs impliqués dans la régulation de la lipolyse et la lipogenèse. .. 11
Figure 2.4 : Expériences de Coleman en circulation croisée impliquant des souris normales (+/+), des souris obèses (ob/ob) et des souris diabétiques (db/db). . 14
Figure 2.5 : Structure tridimensionnelle de la leptine. ... 17
Figure 2.6 : Séquences de l’exon 2 et de l’exon 3 du gène de la leptine. ... 19
Figure 2.7 : Le récepteur de la leptine sous ses six isoformes. ... 22
Figure 2.8 : Voies de signalisation de la leptine à partir de son récepteur. ... 24
Figure 2.9 : Caractère pléïotropique de la leptine. ... 27
Figure 2.10 : Voie métabolique contrôlant l’oxydation des acides gras. ... 32
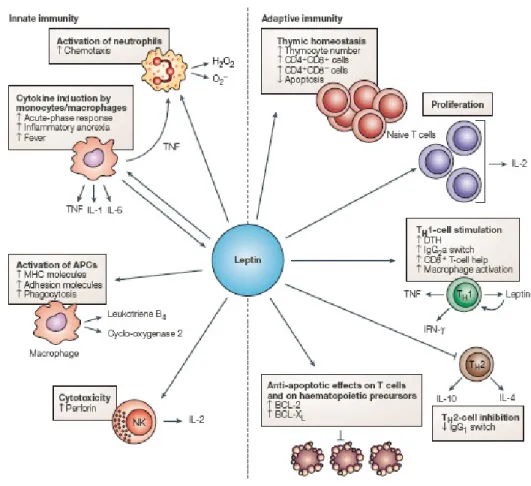
Figure 2.11 : Effets de la leptine sur l’immunité innée (innate) et l’immunité acquise (adaptive). ... 41
Figure 2.12 : Effet angiogénique de la leptine. ... 42
Figure 2.13 : Mécanisme de la résorption osseuse contrôlé par le système nerveux sympathique... 46
viii
Liste des abréviations
α alpha
β beta
δ epsilon
γ gamma
µ micro
ANOVA Analyse de variance
ACC Acétyl-CoA Carboxylase
ADN Acide désoxyribonucléique
ADNc Acide désoxyribonucléique complémentaire
AG Acide gras
AMPK Adenosine monophosphate-activated protein kinase
Ang2 Angiopoïétine 2
ARN Acide ribonucléique
ARNm Acide ribonucléique messager
Asn Asparagine
ASP Protéine stimulant l’acylation
Asp Acide aspartique
ASRA Assurance stabilisation des revenus agricole
BW Poids corporel
CART Cocaine amphetamine regulated transcript
CD Croisement Arcott canadien
CEBP Protéine CEBP ou CCAAT-enhancer-binding proteins
CPT1 Carnitine palmitoyltransferase 1
COX2 Cyclo-oxygénase 2
CS Citrate synthase
db Diabétique
EDTA Acide éthylène diamine tétraacétique
EtOH Alcool éthylique
FAQ Financière agricole du Québec
FGF Facteur de croissance des fibroblastes
ix
GH Hormone de croissance
GLP-1 Glucagon like-peptide 1
GNRH Hormone de libération des gonadotrophines
Hal Gène Halothane
HPG Hypothalamo-hypophyso-gonadique
HSL Lipase hormonosensible
ICV Intracérébroventriculaire
IgG Immunoglobuline G
IRS-1 Substrat du récepteur à l’insuline de type 1
JAK/STAT Janus kinase/Protéine Signal Transducers and
Activators of Transcription
LD Muscle longissimus dorsi
LEP Gène de la leptine
LEPR Gène du récepteur de la leptine
LH Hormone lutéinisante
LPL Lipoprotéine lipase
M Mole
MAP Protéine mitogène activée
MOPS Acide 3-(N-morpholino)propane sulfonique
NK Cellules tueuses
NPY Neuropeptide Y
Ob Obèse
pb (fr) ou bp (an) Paire de bases
PAI-1 Inhibiteur de l'activateur du plasminogène 1
PCR Réaction de polymérisation en chaîne
PCK Protéine kinase C
PI3-K Phosphoinositide 3-kinase
PPAR Récepteur activé par les proliférateurs de peroxysomes
RFLP Polymorphisme de longueur de fragment de restriction
RIA Dosage immuno-isotopique
RN Gène du rendement Napole
SHP-2 Protéine tyrosine phosphatase SHP-2
SNC Système nerveux central
x
SOCS Suppresseur de la signalisation des cytokines
SU Croisement Suffolk
TG Triglycérides
TNF- α Facteur de nécrose tumorale
TRL Lipoprotéine-triacylglycerol
VEGF Facteur de croissance endothélial vasculaire
ad libitum à volonté
de novo nouvellement synthétisé
in vitro observé au laboratoire, en dehors de l'organisme in vivo dans l’organisme vivant
xi
Remerciements
Je tiens à remercier quelques personnes qui m’ont aidé dans le cadre de ce projet de recherche, parfois un peu, parfois beaucoup… qui ont été présentes dans les moments les plus amusants, mais aussi quand plus rien n’allait.
Premièrement, ma conjointe des 17 dernières années, Joëlle, qui, malgré le long délai entre la fin de mes expériences en laboratoire et le dépôt de ce mémoire, n’a jamais cessé de m’encourager dans mes projets. Elle m’a toujours appuyé, encore plus lorsque j’ai entrepris de terminer la rédaction de ce document en novembre 2016.
Un merci spécial à mes parents Francine et Carol, qui m’ont aidé à faire les études de mon choix, loin de la maison, et qui m’ont soutenu pendant les 7 années que j’ai passées à Québec. Merci de m’avoir aidé comme vous l’avez fait. Peu d’étudiants ont cette chance.
Merci à François Castonguay, Marie-France Palin et Claude Gariépy pour les discussions que nous avons eues au cours des années et qui m’ont permis d’avancer mes travaux, mais aussi de redémarrer la rédaction de mon mémoire quand j’ai annoncé que je désirais terminer ce qui avait été commencé.
Merci aux producteurs d’agneaux qui ont fourni les animaux destinés, en partie, à mes expérimentations.
Merci aux membres du personnel de l’abattoir de Luceville pour leur collaboration lors de l’abattage des animaux et la récolte des tissus.
J’aimerais aussi remercier ces collègues au laboratoire et au travail qui m’ont tous, à leur manière, permis d’apprendre à leur contact, que ce soit au niveau technique, scientifique ou humain.
Mireille Thériault et Catherine Boivin pour leur aide dans la partie agronomique et technique du projet de recherche, que ce soit lors des récoltes d’échantillons en bergerie ou lors de l’abattage des animaux à Luceville.
xii
Marie-Christine Fortin pour son aide dans les manipulations à effectuer à l’abattoir, pour la récolte des tissus devant servir à mes analyses.
Dan Lacroix et France Bégin, travaillant à l’époque chez TGN Biotech, pour m’avoir accueilli dans votre laboratoire et m’avoir conseillé sur les méthodes à utiliser dans mes diverses manipulations.
Serge McGraw, Christian Vigneault, Karine Tremblay, et Isabelle Dufort pour leur aide lors de mon apprentissage au laboratoire.
Athanasia Gomatos, pour son aide dans la révision de la qualité de l’écriture en anglais du chapitre 3.
Finalement, merci à mon directeur de maîtrise, François Pothier. Au cours des 15 dernières années, il a d’abord agit comme enseignant, puis comme mentor et comme guide dans mes études, mais est aussi devenu un ami. Je ne crois pas que beaucoup d’étudiants à la maîtrise peuvent affirmer avoir été jouer au badminton de manière régulière avec leur directeur au cours de leur maîtrise et avoir continué à collaborer en enseignant pendant presque 10 ans dans un cours universitaire après avoir quitté le campus pour débuter leur carrière. Merci pour tout ce que j’ai appris et développé comme compétences à tes côtés, pour tes connaissances pour tes qualités humaines, ta patience…
xiii
Avant-propos
L’article scientifique faisant l’objet du chapitre 3 a été rédigé dans l’optique d’une publication dans Canadian Journal of Animal Science, mais n’a pas été publié à ce jour. L’auteur du présent mémoire est l’auteur principal de l’article, qui en est à sa première version publiée dans un document.
Les coauteurs sont François Pothier, à titre de directeur des travaux de recherche, Marie-France Palin, à titre de codirectrice, François Castonguay et Claude Gariépy, chercheurs associés au projet de recherche, de même que Mireille Thériault, assistante de recherche ayant dirigé la phase expérimentale à la ferme.
1
Chapitre 1 : Introduction
Au cours des 20 dernières années, le secteur ovin québécois a connu des transformations considérables. Entre 1994 et 2004, le nombre d’animaux du cheptel de la province a considérablement augmenté, le nombre de brebis enregistrées au programme de stabilisation du revenu de la Financière agricole du Québec (FAQ) passant de 68 000 brebis en 1994 à 179 317 en 2004 (Chesnais et al., 2002; Financière Agricole du Québec, 2005). Il en fut de même pour le nombre d’entreprises ovines, leur nombre passant de 393 à 871 en 2004 (adhérents à la FAQ) (Financière Agricole du Québec, 2016). Puis, à la fin des années 2000, le portrait du secteur a considérablement changé, la Financière Agricole du Québec ayant entamé, en 2009, une réforme du mode de paiement de l’Assurance Stabilisation des Revenus Agricoles (ASRA) aux producteurs (Financière Agricole du Québec, 2016). Le nombre d’entreprises adhérant à l’ASRA a alors chuté pour se trouver, en 2015, à 489 entreprises (152 507 agneaux et brebis assurés) (Financière Agricole du Québec, 2016).
Le mode de compensation de l’ASRA, autrefois basé sur le nombre de brebis en inventaire, est maintenant basé sur le nombre d’agneaux et de kilogrammes d’agneau produits par les entreprises. Ce mode de compensation a provoqué la diminution du nombre de brebis et du nombre d’entreprises ovines sur le territoire québécois. En contrepartie, le changement amorcé en 2009 incite les producteurs à être plus efficaces et à produire plus. Cette situation, combinée au paiement au producteur en fonction de la qualité de la carcasse, force les producteurs à être plus efficaces dans leurs opérations et à chercher à améliorer la génétique de leur troupeau, tant pour la prolificité des animaux, que pour la qualité de la viande et de la carcasse. Ce dernier point a d’ailleurs été identifié comme une priorité par le secteur dans les dernières années (Zins Beauchesne et associés, 2010; Forest Lavoie Conseil, 2015).
L’augmentation de la production de viande d’agneau lourd répond à la demande des consommateurs québécois, ceux-ci désirant une viande produite au Québec et de qualité supérieure, reconnue pour son goût raffiné. Cependant, les acheteurs veulent aussi un agneau jeune et présentant des caractéristiques homogènes, un haut
2
rendement en viande maigre et une proportion de gras équilibrée dans les découpes (pas trop gras, pas trop maigre). Et c’est là un problème pour l’industrie ovine québécoise : les agneaux présentent bien souvent une qualité et des caractéristiques hétérogènes, particulièrement en ce qui concerne la quantité de gras déposée qui est souvent trop élevée (Dumoulin, 2009).
Dans un contexte où le paiement aux producteurs est basé sur un système de classification des carcasses tenant compte de l’épaisseur de gras et le développement musculaire, ceux-ci devront assurer une plus grande homogénéité des produits offerts afin de rentabiliser au maximum leurs efforts et leurs investissements. Idéalement, l’agneau devra être jeune, présenter une grande proportion de viande et une faible proportion de gras afin d’assurer le goût fin tant recherché par le consommateur. Ainsi, afin de rester concurrentiels vis-à-vis des joueurs mondiaux de l’industrie tels que la Nouvelle-Zélande ou l’Australie ou vis-à-vis des concurrents au plan national (Ontario et Ouest canadien), un effort important de sélection génétique s’impose afin d’uniformiser la qualité du produit offert.
Dans cette optique, l’utilisation de marqueurs moléculaires pourrait apporter un élément de réponse important et rapide par rapport à l’utilisation d’indices de potentiel génétique pour la sélection d’individus reproducteurs. Bien que l'utilisation d'indices de potentiel génétique permette un gain génétique relativement rapide pour les caractères liés à la qualité de la viande et de la carcasse, le recours aux marqueurs moléculaires indiquant des variances du code génétique liées à certains phénotypes désirables permettrait, en comparaison, de faire des avancées appréciables plus rapides dans la sélection d’individus reproducteurs donnant une descendance de meilleure qualité.
De nombreux marqueurs génétiques ont été identifiés et utilisés chez des espèces
d’élevage (gènes RN- et Hal chez le porc, par exemple) afin d’améliorer les caractères
reliés à la qualité de la carcasse. Chez les ovins, le gène callipyge a déjà été identifié comme améliorateur des performances liées à la qualité de la carcasse (œil de longe, gigot, adiposité) chez les animaux porteurs de l’allèle favorable provenant du père (Cockett et al., 1996; Cockett et al., 2005).
3
Chez des espèces d’intérêt agronomique, des recherches ont mis en lumière les effets sur la déposition de gras d’une hormone, la leptine, et de polymorphismes présents sur le gène codant pour celle-ci (Ob, LEP), exprimé en très grande partie dans les tissus adipeux (Zhang et al., 1994; Macajova et al., 2004; Wylie, 2011). La leptine agit sur la déposition de gras dans la carcasse et sur la qualité de celle-ci. Chez le porc et le bovin en particulier, certaines mutations portées par le gène de la leptine ont été associées à la déposition de gras dorsal, à la conversion alimentaire et au gain de poids quotidien (Martel-Kennes et al., 2001; Nkrumah et al., 2004; Schenkel et al., 2005).
Dans ce contexte, le gène de la leptine pourrait être un excellent candidat pour identifier les individus présentant potentiellement une meilleure qualité de carcasse chez les ovins. Des avancées en ce sens ont été répertoriées ces dernières années, entre autres dans le cadre de la première phase de ce projet de recherche portant sur l’étude du gène Ob chez des agneaux de race pure Dorset et Suffolk (Boucher et al., 2006). Des effets de polymorphismes de ce même gène ont aussi été documentés pour d’autres caractères de production, tels que le taux de conversion alimentaire (lait) (Jonas et al., 2016) et le poids corporel lors de l’engraissement, mesuré à chaque période de 3 mois d’âge (Shojaei et al., 2010).
4
Chapitre 2 : Revue des travaux antérieurs
2.1. Régulation du poids corporel
Chez les mammifères, la régulation du poids corporel est un phénomène complexe ayant évolué à travers les âges pour permettre aux diverses espèces de s’adapter et de survivre dans des milieux où les conditions n’étaient pas toujours clémentes. Ainsi, la capacité d’entreposer efficacement de l’énergie sous forme de triglycérides a permis et permet toujours aux animaux confrontés à des périodes de jeûne prolongées ou à de fortes demandes énergétiques, comme lors de la lactation, ou à des climats hostiles, de survivre.
Cependant, dans le contexte agroalimentaire actuel, les matières grasses ont mauvaise presse et toute l’industrie agroalimentaire doit s’adapter. Les productions animales, particulièrement celles dont les élevages sont destinés à la production de viande, n’échappent pas à l’exigence des consommateurs qui impose la réduction de la quantité de gras présente dans les aliments. Les producteurs doivent, par conséquent, diminuer la masse de tissu adipeux présent dans les produits carnés.
Ainsi, d’un point de vue agronomique, il est important de comprendre les mécanismes de l’accumulation d’énergie chez les animaux d’élevage, particulièrement dans un contexte de production animale rémunérée selon la qualité de la carcasse, comme c’est le cas pour l’agneau au Québec. La compréhension du contrôle du poids corporel et de l’accumulation de gras des animaux revêt une importance capitale dans le cadre d’une économie de marché, la conversion de la ration alimentaire en produit commercialisable et désiré par le consommateur étant l’un des déterminants de l’efficacité des entreprises d’élevage.
Chez les mammifères, la régulation du poids corporel est un phénomène complexe dépendant d’interactions entre plusieurs facteurs dont la génétique et l’environnement (Jequier, 2002), tous influant de manière plus ou moins importante sur la balance
5
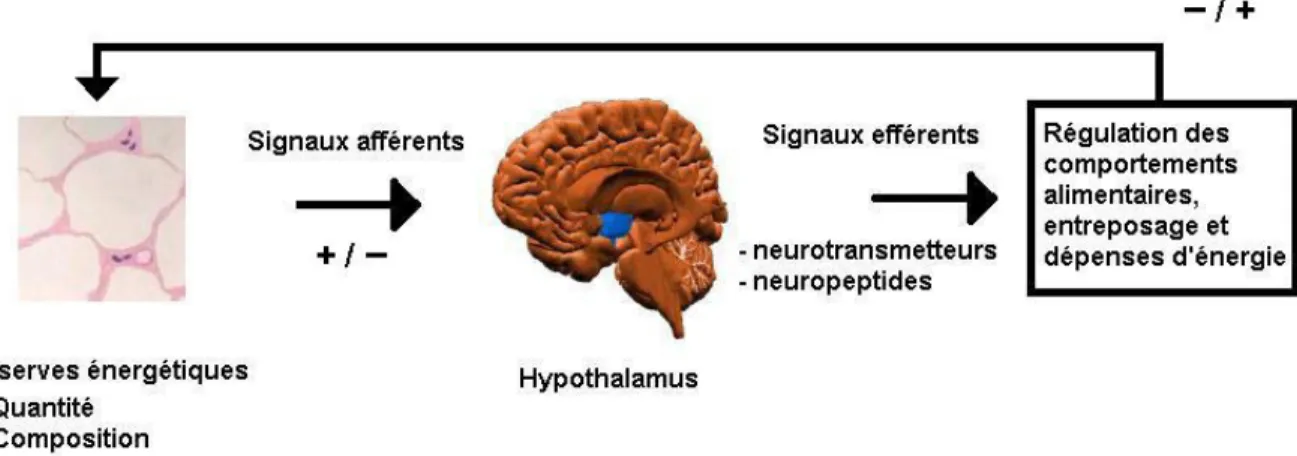
énergétique de l’organisme. La figure 2.1 illustre les principaux éléments impliqués dans
la
Figure 2.1 : Éléments clés de l’équilique énergétique de l’organisme.
6
balance énergétique, soit la prise alimentaire (entrée d’énergie), le métabolisme basal de l’organisme, la thermogénèse et l’activité physique (dépenses d’énergie) (Spiegelman et Flier, 2001). On peut aussi inclure la croissance et la production laitière dans les paramètres de dépense énergétique.
Un léger déséquilibre entre la prise alimentaire et la dépense énergétique peut donc résulter, à plus ou moins long terme, en une accumulation excessive des réserves énergétiques, donc en une augmentation de la quantité de triglycérides accumulés dans les tissus adipeux.
Les chercheurs tentent depuis longtemps de comprendre les mécanismes de la régulation du poids corporel chez les mammifères, permettant la conservation d’un état de chair stable dans le temps. Déjà en 1953, Kennedy, dans son hypothèse lipostatique, proposait un mécanisme de contrôle du poids corporel. Celui-ci suggérait que des signaux inhibiteurs, générés en proportion de la quantité de réserves adipeuses de l’organisme, agissaient au niveau du cerveau, plus précisément sur l’hypothalamus, afin d’inhiber la prise alimentaire et conserver un poids stable de l’organisme. Ainsi, lorsque l’organisme subit un déficit énergétique (diminution de ses réserves adipeuses), le niveau des signaux inhibiteurs diminuerait et la prise alimentaire augmenterait jusqu’à ce que la situation soit rétablie (Kennedy, 1953). Cependant, l’hypothèse émise par Kennedy n’expliquait pas le contrôle de la prise alimentaire à court terme (Schwartz et
al., 2000). C’est Gibbs et ses collaborateurs, en 1973, qui ont proposé que des facteurs
de satiété, comprenant des peptides sécrétés par le tractus digestif, étaient produits pendant la prise alimentaire et menaient à la cessation du repas (Gibbs et al., 1973). Ceux-ci fournissent au cerveau l’information concernant l’état de satiété à court terme. Plus tard, l’hypothèse du point défini ou set point hypothesis a été proposée afin d’expliquer la régulation à long terme de la balance énergétique de l’organisme. Celle-ci suggère que la prise alimentaire et la dépense énergétique sont coordonnées par les centres végétatifs du système nerveux central (SNC) afin de maintenir un équilibre relativement stable de la quantité d’énergie accumulée dans l’organisme et, par conséquent, le poids de celui-ci (Friedman et Leibel, 1992). Le statut des réserves énergétiques, selon cette hypothèse, serait sous la surveillance constante du SNC et plus particulièrement de l’hypothalamus. La moindre déviation par rapport à l’équilibre établi serait ajustée par l’intermédiaire de la régulation de la prise alimentaire et de la
7
dépense énergétique de l’organisme. Ces ajustements se feraient par l’intermédiaire de signaux afférents (provenant des réserves) vers l’hypothalamus, signalant tout changement des réserves énergétiques (quantitatif ou qualitatif). Par la suite, le système nerveux dicterait la réponse appropriée à la situation (régulation de l’entreposage et de la dépense énergétique, comportements d’ingestion), comme illustré à la figure 2.2.
8
Figure 2.2 : Hypothèse du point défini ou Set Point Hypothesis.
9
Les recherches subséquentes ont montré qu’il existe bien des mécanismes complexes permettant la communication entre les réserves énergétiques de l’organisme, le SNC et les autres tissus et, par conséquent le contrôle de l’homéostasie énergétique de l’organisme. Un nombre grandissant de molécules impliquées dans ces mécanismes ont été découvertes, illustrant la grande complexité du processus du contrôle des réserves énergétiques (Schwartz et al., 2000; Morton et al., 2006). La complexité de ce système est d’autant plus grande que la synthèse des facteurs produits par l’organisme lui-même est influencée non seulement par le statut énergétique et la prise alimentaire, mais aussi par des facteurs environnementaux, la génétique et le caractère propre de l’individu.
2.2. Le tissu adipeux
2.2.1. Physiologie du tissu adipeux
Le tissu adipeux est un tissu conjonctif lâche composé en majeure partie de cellules de trois types, soit les adipocytes remplis de lipides (2/3 des cellules), les préadipocytes et les cellules endothéliales. Les adipocytes proviennent de la différenciation des préadipocytes. La formation du tissu adipeux commence vers le milieu ou la fin du développement embryonnaire et se poursuit tout au long de la vie, en fonction des conditions environnementales, spécialement en fonction du statut nutritionnel (Feve, 2005). La différentciation des cellules du tissu adipeux est régulée de manière très fine. Plusieurs facteurs de transcription jouent un rôle dans la différentiation des adipocytes, mais PPAR-γ et δ de même que C/EBPα, β et δ apparaissent comme étant les régulateurs clés du processus (Feve, 2005; Luquet et al., 2005; Tang et Lane, 2012). De plus, nombre de facteurs physiologiques sont impliqués dans ce processus (hormones, peptides, nutriments). Les glucocorticoïdes, les acides gras et l’insuline, entre autres, jouent un rôle important dans la différentiation des préadipocytes en adipocytes.
En plus de ces cellules, on retrouve dans le tissu adipeux des fibres nerveuses de même que des macrophages et des vaisseaux sanguins. Les fibres nerveuses permettent au tissu de communiquer avec le reste du système nerveux alors que les vaisseaux sanguins permettent l’échange de nutriments et de signaux avec les autres organes. La composition du tissu adipeux varie substantiellement en fonction de son emplacement dans l’organisme, conférant à certains dépôts graisseux une activité métabolique plus ou moins élevée (tissus adipeux blancs vs tissus adipeux bruns) et,
10
par conséquent, une capacité variable à produire des molécules actives. À titre d’exemple, le gras intra-abdominal est plus innervé et contient plus de vaisseaux sanguins que le gras sous-cutané, indiquant une activité métabolique plus importante (Hauner, 2004).
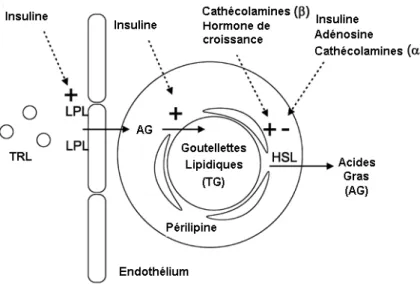
Le tissu adipeux évolue dans le temps, augmentant ou diminuant ses réserves de triglycérides par le biais, respectivement, de la lipogenèse et de la lipolyse. La lipogenèse se produit lors de l’hydrolyse de lipoprotéine-triacylglycerol (TRL) circulante par la lipoprotéine lipase (LPL) en acides gras qui seront estérifiés par les adipocytes (figure 2.3). L’activité de la LPL, une enzyme exprimée dans le tissu adipeux, et l’estérification des acides gras sont stimulées par l’insuline (Frayn et al., 2003). La lipogenèse se produit aussi de novo, à partir de substrats non-lipidiques. Ce processus a été observé chez les rongeurs, mais serait d’importance mineure chez l’humain (Daval
et al., 2006).
La lipolyse se produit lors de la mobilisation des réserves énergétiques et requiert l’action de l’enzyme lipase hormonosensible (HSL). Cette enzyme est phosphorylée afin d’être activée et transloquée du cytosol de l’adipocyte vers la gouttelette lipidique. La périlipine entourant la gouttelette est phosphorylée et retirée afin de laisser l’HSL accéder à celle-ci et réaliser la lipolyse (Frayn et al., 2003). Ce processus est stimulé par les catécholamines β et l’hormone de croissance (GH) (figure 2.3).
11
Figure 2.3 : Facteurs impliqués dans la régulation de la lipolyse et la lipogenèse. TRL : lipoprotéine-triacylglycérol; LPL : Lipoprotéine lipase; HSL : Lipase hormonosensible; TG : triglycérides
12
2.2.2. Le tissu adipeux : un tissu dynamique
Jusqu’à tout récemment, le tissu adipeux était considéré comme un réservoir de triglycérides pour le corps et le rôle qui lui était attribué était plutôt passif. Sa fonction consistait essentiellement à accumuler et relâcher des réserves énergétiques, selon la balance énergétique de l’organisme. Cependant, la découverte, il y a environ 20 ans, de la leptine et du fait que le tissu adipeux est un site majeur du métabolisme des stéroïdes sexuels et d’adipocytokines a complètement changé la donne (Hauner, 2004; Otero et
al., 2005). Les données s’accumulant, il est aujourd’hui évident que le tissu adipeux
n’est pas un tissu de cellules passives, mais plutôt un organe multifonctionnel produisant de nombreuses molécules pouvant agir de manière endocrine, paracrine ou autocrine (Coelho et al., 2013). Il est maintenant acquis que le tissu adipeux est au coeur de la communication entre sa propre entité, le système nerveux et les autres organes. Plus de 100 molécules produites et sécrétées par le tissu adipeux ont été décrites, mais toutes n’ont pas une fonction connue (Hauner, 2004).
Les adipocytes sécrètent non seulement une grande variété de protéines, mais aussi des prostaglandines, des hormones stéroïdiennes et possiblement d’autres composés contribuant à la communication avec l’organisme et le contrôle de la balance énergétique. Plusieurs molécules sont impliquées dans le contrôle du métabolisme des lipides. Parmi celles-ci, figurent les cytokines (leptine, TNF-α, adinopectine), l’interleukine-6 (IL-6), la résistine, le PAI-1, l’adipsine, l’ASP, la LPL ainsi que tous les composants du système rénine-angiotensine (Frayn et al., 2003). Ces molécules agissent sur le métabolisme lipidique soit de manière endocrine, paracrine ou une combinaison des deux et à des moments différents, certaines produisant leurs effets sur la différentiation des préadipocytes alors que d’autres agissent sur la lipolyse.
Les adipocytes ne seraient pas les seules cellules du tissu adipeux à sécréter des molécules actives. Il s’avère de plus en plus évident que les préadipocytes et les cellules endothéliales seraient en mesure de sécréter quelques facteurs, contribuant à la grande diversité de produits observés dans ce tissu (Hauner, 2004). Par exemple, les préadipocytes seraient le site presque exclusif de la production des œstrogènes chez les femmes ménopausées, une arômatase, qui en contrôle la production, y étant exprimée (Cleland et al., 1985).
13
2.3. La leptine : généralités
2.3.1. Travaux préliminaires
Les premiers travaux laissant penser qu’il existait une molécule agissant comme facteur de satiété datent des années ‘70 alors que Coleman effectua au laboratoire Jackson des expériences de parabiose (circulation croisée) sur des rongeurs obèses (ob/ob) et des rongeurs diabétiques (db/db) de souche C57BL/6J (Coleman, 1978). Les souris obèses (ob/ob) possèdent un surplus de poids important et présentent un comportement d’hyperphagie, une glycémie sanguine et une concentration d’insuline plasmatique anormalement élevées. Les rongeurs diabétiques, pour leur part, présentent aussi un phénotype montrant un surplus de poids, un comportement hyperphagique et un diabète important lié à une glycémie fortement élevée (Coleman, 1978). Comme les phénotypes des animaux étaient, à toute fin pratique, identiques dans un même environnement, il était permis de croire que les deux gènes impliqués occupaient une fonction dans le même système (Coleman, 1978).
Les expériences de parabiose menées par Coleman sont résumées à la figure 2.4. Lors du croisement de la circulation sanguine d’une souris normale (+/+) avec celle d’une souris db/db, la souris normale voit son insulinémie diminuer fortement de même que son taux de glucose sanguin. L’union des deux rongeurs résulte en la mort du sujet normal suite à un jeûne prolongé. Le phénotype du rongeur diabétique reste inchangé.
14
Figure 2.4 : Expériences de Coleman en circulation croisée impliquant des souris normales (+/+), des souris obèses (ob/ob) et des souris diabétiques (db/db).
Adaptée de Coleman, 1978 +/+ ↓ insulinémie
↓glucose sanguin mort par inanition db/db ↑ poids corporel
↑ masse de tissu adipeux
ob/ob ↓ poids corporel
↓ masse de tissu adipeux ↓ insulinémie
↓ glucose sanguin mort par inanition db/db ↑ poids corporel
↑ masse de tissu adipeux
+/+ aucun changement ob/ob ↓ prise alimentaire
↓ insulinémie ↓ glucose sanguin
15
Cela démontre l’existence d’un facteur de satiété puissant, mais auquel la souris diabétique ne peut répondre. Ce fait aidant, l’union de deux rongeurs db/db ne mène pas au décès des animaux, ceux-ci étant inaptes à répondre au facteur de satiété même s’il est présent dans la circulation (Coleman, 1978).
Pour ce qui est de l’union de souris db/db à des souris ob/ob, seul le rongeur diabétique (db/db) survit et ne montre pas de changement dans son phénotype. Cela indique donc que les souris obèses, tout comme les souris normales, sont aptes à répondre au facteur de satiété et qu’une surdose de ce facteur inhibe la prise alimentaire et peut mener ultimement à la mort (Coleman, 1978).
Finalement, l’union d’un rongeur ob/ob à un rongeur +/+ mène à un retour à la normale du phénotype de l’animal obèse et n’entraîne la mort d’aucun des deux sujets. L’appétit n’étant pas inhibé chez la souris normale, il est permis de penser que la souris obèse ne produit pas ou produit en trop faible quantité le facteur de satiété supposé. Cependant, l’inverse étant observé, il est évident que la souris normale produit le facteur de satiété (Coleman, 1978).
Globalement, les expériences de Coleman ont mis en évidence l’existence d’une molécule dans la circulation sanguine apte à contrôler le comportement alimentaire de l’animal. Ces travaux montrent, par le fait même, que les souris ob/ob sont inaptes à produire le facteur de satiété en question alors que les souris db/db sont inaptes à y répondre, expliquant leur phénotype. Aujourd’hui, il est reconnu que la molécule servant de facteur de satiété dans ces expériences est la leptine et que la mutation observée chez la souris ob/ob mène à la production d’une protéine tronquée inactive alors que chez la souris db/db, le récepteur est incomplet et, de ce fait, inactif.
2.3.2. Caractérisation
En réponse aux questionnements soulevés par les expériences de Coleman à la fin des années 70, Zhang et ses collaborateurs ont, en 1994, identifié le gène muté responsable du phénotype d’obésité observé chez les souris ob/ob. Ainsi, le gène de l’obésité (Ob) fut localisé et identifié par clonage positionnel chez les souris ob/ob présentant un phénotype d’obésité morbide et d’infertilité (Zhang et al., 1994). Ces souris sont porteuses d’une mutation autosomale récessive. Le gène est présent sur le
16
chromosome 6 chez la souris et encode un ARNm de 4,5 kb. Chez l’humain, le gène est situé sur le chromosome 7; sa longueur totale est d’environ 20 kb et encode un ARNm d’environ 3,5 kb. Le gène comporte trois exons dont seuls les deux derniers sont codants (Isse et al., 1995).
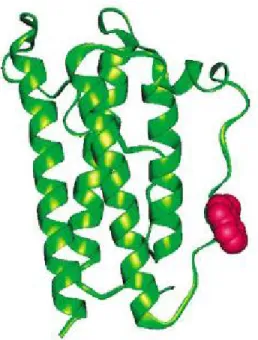
Le produit de la transcription de ce gène est une hormone, la leptine, du grec leptos (mince), faisant référence à son effet anti-obésité (Hegyi et al., 2004). Cette hormone protéinique de 16 kD, par sa structure tridimensionnelle à quatre hélices groupées formant un domaine hydrophobe (figure 2.5), est associée à la famille des cytokines de type 1 (Zhang et al., 1997). Sa séquence primaire se compose de 167 acides aminés. Une séquence N-terminale de 21 acides aminés (issue du début de l’exon 2) constitue le signal peptide qui sera éventuellement éliminé suite à la translocation du peptide dans l’appareil de Golgi pour ainsi donner le peptide actif de 146 acides aminés. Cette transformation précède le passage de la leptine dans le sang (Macajova et al., 2004). Dans le sang, la concentration de leptine est influencée par plusieurs facteurs dont, entre autres, la prise alimentaire, la composition de la ration, le sexe, la température ambiante, l’exercice, le sommeil et la balance énergétique récente. Cette influence se fait surtout sentir par l’intermédiaire de l’insuline, le taux d’entrée du glucose dans les adipocytes et par des régulateurs du système nerveux sympathique (SNS) (Frayn et al., 2003).
17
Figure 2.5 : Structure tridimensionnelle de la leptine.
18
Chez les souris obèses, la protéine s’avère être tronquée à partir de l’acide aminé 105 (Zhang et al., 1994; Grasso et al., 1997). Or, des travaux suggèrent que la partie active de la protéine se trouverait, partiellement, entre les acides aminés 106 et 140, la séquence se situant entre les résidus 116 et 130 ayant démontré la plus forte activité lorsqu’injectée à des rongeurs obèses (Grasso et al., 1997).
Initialement, la leptine a été identifiée comme un facteur de satiété permettant le contrôle des réserves énergétiques corporelles et de la prise alimentaire. En effet, sa concentration dans le sang est positivement reliée aux réserves lipidiques de l’organisme (Ahima et Osei, 2004). La concentration sanguine de leptine indique à l’organisme l’état de satiété et l’état de manque d’énergie alimentaire.
Au fil des ans cependant, la leptine fut associée à plusieurs autres processus physiologiques dénotant son caractère pléiotropique. Des recherches ont mis en lumière son rôle dans le contrôle non seulement du flux lipidique, mais aussi de processus tels que le contrôle de la reproduction, l’angiogenèse, l’immunité et le métabolisme osseux. Aujourd’hui, le gène de la leptine a été cloné chez plusieurs espèces domestiques d’intérêt agronomique, dont le porc, le mouton, et le bovin (Bidwell et al., 1997; Dyer et
al., 1997a; Ji et al., 1998). Le gène de la leptine a été séquencé aussi chez le poulet
(Taouis et al., 1998), mais les résultats de cette étude font encore l’objet de débats dans le milieu scientifique, plusieurs équipes de recherche n’ayant pas été en mesure de confirmer ces résultats (Pitel et al., 2010). Une publication récente suggère que le gène de la leptine, chez le poulet, pourrait avoir disparu au cours de l’évolution, laissant le gène du récepteur orphelin (Pitel et al., 2010).
L’analyse des séquences, fait ressortir une forte homologie inter-espèces chez les mammifères (>85 %), laissant présager un rôle similaire d’une espèce à l’autre, d’où l’intérêt de son étude chez les ovins (figure 2.6).
19
Figure 2.6 : Séquences de l’exon 2 et de l’exon 3 du gène de la leptine.
La forte homologie de la séquence du gène entre les différentes espèces (OA : mouton ; SS : porc ; BT : vache ; MM : souris ; HS : humain) laisse présager un rôle similaire d’une à l’autre. Une forte homologie est observée pour tout l’exon 3.
20
2.3.3. Sites d’expression de la leptine
La leptine est principalement produite par les adipocytes blancs. Cependant, le gène est aussi exprimé, mais de manière beaucoup moins importante, dans les tissus adipeux bruns, le fœtus, le placenta, l’épithélium de la glande mammaire, les muscles squelettiques, l’hypophyse, le tractus gastro-intestinal et les fibroblastes (Ahima et Osei, 2004; Kokta et al., 2004; Chilliard et al., 2005). Chez les ruminants, le gène de la leptine est exprimé dans la paroi le rumen, l’abomasum et le duodénum (Chilliard et al., 2005). Il a aussi été montré chez des fœtus de souris que la leptine est exprimée dans certains organes dont le cœur, les os, les cartilages et les follicules pileux (Hoggard et al., 1998). Aquila et ses collaborateurs (2005) ont mis en évidence la sécrétion de leptine par les spermatozoïdes humains, ce qui ouvre la porte à de nouvelles considérations face au rôle de la leptine dans la fertilité masculine. Des recherches subséquentes n’ont pu démontrer d’effets de la leptine sur la motilité et la capacitation des spermatozoïdes (Li
et al., 2009), mais ont permis d’observer un effet négatif d’un traitement à la leptine sur
la concentration et la morphologie de ces cellules chez le rat (Almabhouh et al., 2015). Chez le poulet, le site où l’ARNm de la leptine est le plus exprimé semble être le foie, organe majeur dans la synthèse du gras chez les oiseaux (Macajova et al., 2004). Cependant, aucune synthèse de la leptine n’a été détectée dans cet organe chez les mammifères. La synthèse de la leptine a aussi été observée dans le foie de poussins au stade embryonnaire, de même que dans le sac vitellin (Ashwell et al., 1999).
2.3.4. Le récepteur de la leptine et ses sites d’expression
Le gène du récepteur de la leptine (LEPR) a été identifié chez la souris par clonage d’expression en 1995 (Tartaglia et al., 1995). Depuis, il a été identifié chez l’homme et plusieurs espèces domestiques. Par homologie de séquences, le récepteur de la leptine a été classé dans la famille des récepteurs des cytokines de classe I (Tartaglia et al., 1995), groupe de récepteurs comportant un domaine homologue aux récepteurs des cytokines dans la partie extracellulaire (Ahima et Osei, 2004).
21
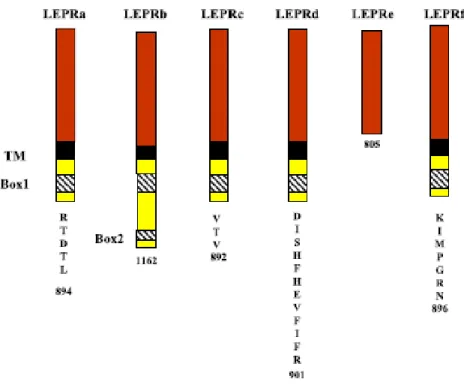
À ce jour, six isoformes du récepteur (a, b, c, d, e et f) ont été identifiées (Ahima et Osei, 2004; Zieba et al., 2005). Elles sont obtenues par épissage alternatif de l’extrémité de l’ARNm encodant la protéine. Toutes possèdent un domaine extracellulaire de 840 acides aminés identique à l’extrémité N-terminale et liant la leptine, mais diffèrent au niveau de l’extrémité C-terminale, soit les domaines transmembranaire et intracellulaire. Cinq des six isoformes (a, b, c, d et f) possèdent un domaine transmembranaire. De celles-ci, seule la forme LEPRb, la forme longue du récepteur, possède un domaine intracellulaire complet présentant tous les motifs protéiques nécessaires à l’activation maximale du système de signalisation « Janus tyrosine kinase/Signal Transduction and Activator of Transcription » (JAK/STAT), soit les motifs Box 1 et Box 2 (Ahima et Osei, 2004; Hegyi et al., 2004; Zieba et al., 2005). Les différentes isoformes du récepteur sont de longueurs différentes et présentent donc des séquences terminales d’acides aminés variées (figure 2.7).
22
Figure 2.7 : Le récepteur de la leptine sous ses six isoformes.
Les deux parties supérieures représentent respectivement le domaine
extracellulaire (rouge) et le domaine transmembranaire (TM, noir). Le domaine intracellulaire comporte deux motifs importants (Box 1 et Box 2) impliqués dans l’activation du signal transmis par la voie JAK/STAT. Box 2 est présent uniquement chez LEPRb, la forme longue et active du récepteur.
23
Bien que le système JAK/STAT soit considéré comme étant le mécanisme principal de transmission du signal activé par le LEPR (Hegyi et al., 2004), il n’en reste pas moins que d’autres mécanismes tels que les MAP Kinases, PI3-K, SHP-2 et IRS-1 (figure 2.8) sont aussi impliqués dans la transmission du signal du récepteur de la leptine (Niswender et al., 2001), par des mécanismes cellulaires variés. Par exemple, la voie de signalisation des MAPK serait impliquée dans le contrôle des effets antiapoptotiques de la leptine sur les cellules immunitaires (Matarese et al., 2005) et les cellules adipeuses (Hegyi et al., 2004), tandis que la voie de signalisation PI3-K serait sollicitée dans la régulation de l’absorption du glucose par le muscle (Matarese et al., 2005).
24
Figure 2.8 : Voies de signalisation de la leptine à partir de son récepteur.
Le système de signalisation JAK/STAT est la principale voie de signalisation du récepteur de la leptine. Après la liaison de la leptine à son récepteur, JAK2 est activé et phosphorylé puis, phosphoryle à son tour le domaine
intracellulaire du récepteur. Les sites phosphorylés serviront de points d’encrage à STAT3, un substrat de JAK 2. STAT3 sera dimérisé avant d’entrer dans le noyau de la cellule pour activer SOCS3 et d’autres gènes. SOCS3 agit comme signal de retour négatif inhibant le signal. D’autres voies alternatives sont aussi possibles, entre autres, les voies MAPK, PI3-K, IRS-1 et SHP-2.
25
Le site d’expression majeur de la forme longue du gène LEPR se situe au niveau de l’hypothalamus, au centre du contrôle de la satiété, où le récepteur est exprimé en grand nombre. Cependant, la forme longue du récepteur est aussi exprimée, en moins forte proportion, dans les tissus adipeux, les cellules endothéliales, le pancréas, les cellules souches hématopoïétiques, le foie, le cœur, les poumons, les testicules, les ovaires, tous les types de cellules régissant l’immunité, acquise ou innée, les ostéoblastes et les reins (Lee et al., 1996; Reseland et al., 2001; La Cava et Matarese, 2004).
Le rôle joué par les isoformes courtes du récepteur demeure encore aujourd’hui incertain. Même si elles (a, c, d et f) présentent un domaine intracellulaire incomplet, elles présentent tout de même la première série d’acides aminés nécessaire à la reconnaissance de JAK et, par conséquent, à l’activation de certaines voies de signalisation (Hegyi et al., 2004). Une équipe de recherche a émis l’hypothèse que la fonction principale des formes courtes du récepteur consisterait en un rôle dans l’internalisation et la dégradation de la leptine (Uotani et al., 1999). Elles seraient aussi impliquées dans le transport et l’élimination de la leptine du système de même que dans le transport de la leptine au travers de la barrière hémato-céphalique (Hegyi et al., 2004). L’isoforme « e » du récepteur de la leptine ne présente quant à elle ni domaine transmembranaire ni domaine intracellulaire et circule dans le sang en tant que récepteur soluble de la leptine (Tartaglia, 1997). Son rôle se résumerait à réguler la quantité de leptine libre, et par conséquent active, dans la circulation (Hegyi et al., 2004) Le signal induit par la leptine est inhibé par SOCS-3 (« Supressor Of Cytokine Signalling-3 »), une protéine membre d’une famille de facteurs inhibant la cascade de signalisation JAK/STAT (Ahima et Osei, 2004). La production des protéines de cette famille est induite par les cytokines, dont fait partie la leptine, et inhibe par une boucle de rétroaction négative la voie de signalisation du récepteur de la leptine.
26
2.3.5. Fonctions physiologiques de la leptine
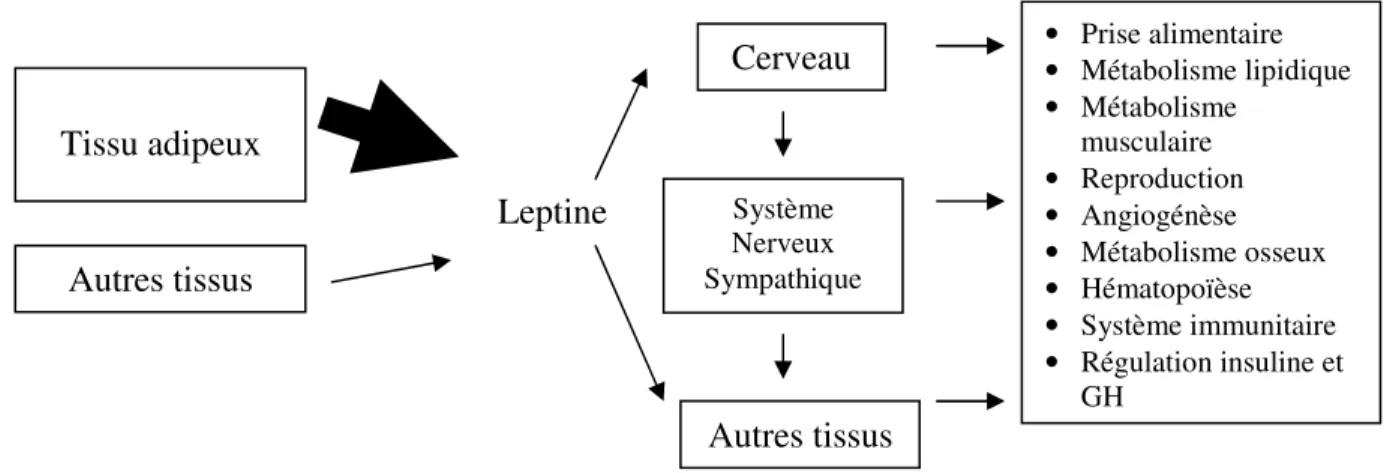
La leptine, depuis sa découverte en 1994, a fait l’objet de nombreux travaux. Ceux-ci ont rapidement révélé ses multiples fonctions, beaucoup plus nombreuses que ce qui était initialement perçu. Comme mentionné précédemment, les transcrits du gène de la leptine et de son récepteur ont été localisés dans plusieurs tissus autres que le tissu adipeux, ce qui a mené à la découverte de différents rôles de l’hormone. Parmi ceux-ci, notons les os, le placenta et les muscles, dénotant le caractère pléïotropique de la leptine (figure 2.9).
27
Figure 2.9 : Caractère pléïotropique de la leptine.
Adaptée de Chilliard et al., 1999 Tissu adipeux Autres tissus Leptine Cerveau Autres tissus Système Nerveux Sympathique • Prise alimentaire • Métabolisme lipidique • Métabolisme musculaire • Reproduction • Angiogénèse • Métabolisme osseux • Hématopoïèse • Système immunitaire • Régulation insuline et GH
28
2.3.5.1 Régulation de la prise alimentaire et métabolisme lipidique
La leptine a initialement été identifiée comme une hormone agissant au niveau du cerveau pour inhiber la prise alimentaire et augmenter la dépense énergétique, permettant ainsi la diminution de la masse adipeuse de l’organisme après injection de l’hormone chez des rongeurs obèses (ob/ob) (Campfield et al., 1995; Halaas et al., 1995; Pelleymounter et al., 1995). Plusieurs études ultérieures ont révélé le rôle joué par la leptine dans l’organisme quant à son effet sur le contrôle de la prise alimentaire et l’adiposité (Ahima et Osei, 2004).
Prise alimentaire
Chez les souris obèses ob/ob, une diminution significative de la prise alimentaire a été observée après injection intrapéritonéale de leptine recombinante alors que chez les souris normales, cette réduction était observée, mais de moindre importance (Campfield
et al., 1995; Halaas et al., 1995; Pelleymounter et al., 1995). L’injection de leptine mène,
par conséquent, à la perte de poids de l’animal par la diminution de sa masse adipeuse. Un phénomène semblable a été observé chez le porc (Barb et al., 1998).
L’effet de la leptine dans le mécanisme de contrôle de la prise alimentaire est antagoniste à celui d’un autre peptide agissant sur la prise alimentaire, le neuropeptide Y (NPY). Ce peptide produit par l’hypothalamus induit fortement l’appétit et diminue la dépense d’énergie. La leptine a pour effet de diminuer l’expression du gène codant pour NPY (Stephens et al., 1995; Ahima et al., 1996; Schwartz et al., 1996a; Schwartz et al., 1996b). À titre d’exemple, chez les souris ob/ob, la leptine étant inactive, le neuropeptide Y est surexprimé, provoquant l’hyperphagie et par conséquent, un phénotype d’obésité. Chez les souris db/db, le récepteur de la leptine étant inactif, la leptine ne peut exercer son effet, ce qui résulte aussi en une surexpression de NPY et un phénotype d’obésité. Lors de l’injection de la leptine, l’expression du gène codant pour NPY est fortement diminuée chez les souris ob/ob, ce qui entraîne une diminution de la prise alimentaire et une perte de poids marquée. Cependant, le même phénomène n’est pas observé chez les souris db/db (Schwartz et al., 1996a; Dyer et al., 1997b). Ces différentes études montrent l’importance de la leptine dans la régulation de l’expression de NPY et, par
29
conséquent, de la prise alimentaire. Inversement, NPY exerce une régulation positive sur l’expression de la leptine.
Chez l’agneau, une injection de NPY dans le tissu adipeux augmente l’expression périphérique du gène de la leptine, probablement par l’intermédiaire du récepteur NPY-Y1 (Dyer et al., 1997b). Cependant, NPY ne serait pas le seul peptide impliqué dans la régulation positive de la prise alimentaire et n’est pas essentiel au contrôle de la prise alimentaire par la leptine. Les souris transgéniques dont le gène NPY est inactivé montrent une prise alimentaire normale et répondent tout de même à un traitement à la leptine qui réduit leur prise alimentaire et amène une perte de poids (Erickson et al., 1996).
En plus de son action sur NPY, un peptide anabolique, la leptine exerce son action sur d’autres neuropeptides, ceux-ci ayant plutôt une action catabolique (Cameron et al., 2003). En fait, la leptine semble interagir avec la plupart des neuropeptides connus pour leur implication dans la régulation de la balance énergétique et de la prise alimentaire. Le glucagon-like peptide 1 (GLP-1), l’hormone de libération de la corticotrophine, la pro-opiomélanocortine, le « Cocaine Amphetamine Regulated Transcript » (CART), l’orexine et la galanine sont quelques-uns de ces neuropeptides (Macajova et al., 2004).
Métabolisme lipidique
De manière générale, lors de l’augmentation de la concentration plasmatique de leptine, une diminution de la prise alimentaire est observées, mais aussi une diminution de la disponibilité des nutriments pour les adipocytes, de même que des changements hormonaux provoquant une diminution de la lipogenèse et une augmentation de la lipolyse (Chilliard et al., 1999). La leptine inhibe la lipogenèse chez les adipocytes porcins. Cet effet serait dû à une inhibition de l’oxydation du glucose et de la lipogenèse. De plus, elle stimulerait l’oxydation des acides gras tout en inhibant leur estérification (Ramsay, 2003a). Une augmentation semblable de la lipolyse a été remarquée chez la souris ob/ob lorsque soumise à une injection de leptine recombinante (Fruhbeck et al., 1998). La leptine exerce un effet sur différents gènes clés du métabolisme lipidique, augmentant l’expression des gènes de protéines thermogéniques et d’enzymes lipolytiques (notamment de la lipase hormonosensible (HSL), des protéines de
30
découplage 1 et 2 et la lipoprotéine lipase) et diminuant l’activité de l’enzyme acide gras synthase (Sarmiento et al., 1997).
La leptine agirait aussi en inhibant l’effet lipogénique de l’insuline. Des études in vitro ont montré que l’hormone agissait au niveau du pancréas en inhibant directement la sécrétion d’insuline (Emilsson et al., 1997) et en atténuant l’activité de celle-ci (Cohen et
al., 1996). De plus, la leptine agirait directement sur les adipocytes en empêchant la
liaison de l’insuline comme cela fût observé in vitro sur des adipocytes de rat (Muller et
al., 1997; Walder et al., 1997).
2.3.5.2 Métabolisme et croissance musculaire
Il a été observé que la leptine avait quelques effets notables sur le métabolisme et la croissance musculaire. En effet, chez des cellules musculaires de poulet mises en culture, la leptine a un effet favorisant la prolifération cellulaire de même que la synthèse protéique (Lamosova et Zeman, 2001). L’action de la leptine observée dans ces procédés cellulaires s’est avérée dose-dépendante et elle variait en fonction de l’âge des embryons.
Chez le bovin, la leptine agit au niveau de l’hypophyse antérieure pour moduler la sécrétion de l’hormone de croissance (GH). L’effet de la leptine sur la sécrétion de GH est cependant tributaire de l’état nutritionnel dans lequel se trouve l’animal (Zieba et al., 2003b). Un effet semblable a été noté chez le porc (Barb et al., 1998) et le mouton (Henry et al., 2001). Or, l’hormone de croissance est d’une importance capitale pour la croissance musculaire, augmentant l’accumulation protéique au détriment des dépôts adipeux (Cassar-Malek et al., 1998). Cette relation entre la leptine et la GH pourrait s’avérer très intéressante à explorer du point de vue agronomique afin de produire des animaux présentant un meilleur rendement en viande maigre.
De plus, une augmentation de l’absorption du glucose par les muscles squelettiques a été observée suite à l’injection de leptine dans l’hypothalamus (Minokoshi et al., 1999). Une augmentation de l’expression du gène de la leptine et de sa concentration protéique dans le muscle squelettique suite à l’hyperglycémie et l’hyperlipidémie, montre le lien
31
entre l’état nutritionnel de l’organisme et l’expression du gène de la leptine dans le muscle, comme dans le tissu adipeux (Wang et al., 1998).
Chez la souris, le rat et l’humain, la leptine augmente directement la production de CO2
dans les muscles squelettiques (Ceddia, 2005). De plus, la leptine stimule l’activité de la glycolyse et du cycle de Krebs dans le muscle soleus, deux mécanismes impliqués dans le métabolisme du glucose et des lipides au niveau de la mitochondrie. Comme mentionné précédemment, l’effet de la leptine sur le métabolisme du glucose dans les cellules musculaires est régulé par la voie de signalisation PI3-K. Il en est de même pour l’effet thermogénique de l’hormone, effet pouvant influer sur le métabolisme des acides gras et du glucose (Ceddia, 2005). Il a aussi été observé que la leptine augmente l’intensité de l’oxydation des acides gras dans le muscle (Muoio et al., 1997). L’hormone agit sur l’activation de l’AMPK en augmentant son expression ce qui entraîne une chaîne d’inhibitions menant à une augmentation de l’oxydation des acides gras (figure 2.10) (Minokoshi et al., 2002; Steinberg et al., 2003).
32
Figure 2.10 : Voie métabolique contrôlant l’oxydation des acides gras.
La leptine stimule l’AMPK, provoquant par le fait même la phosphorylation de l’acéthyl-CoA Carboxylase (ACC). Cela mène à une inhibition du malonyl-CoA qui entraîne la levée de l’inhibition de la carnitine palmitoyltransferase 1 (CPT1), résultant ainsi en l’augmentation de l’oxydation des acides gras dans le muscle squelettique.
33
La perte de poids provoquée par l’administration de leptine amène une diminution spécifique de la masse adipeuse tout en préservant le muscle. En comparaison, la perte de poids provoquée par la sous-nutrition amène la perte simultanée des deux types de tissus (Chilliard et al., 1999). Chez le porc, un traitement avec de la leptine entraîne un effet protecteur contre la dégradation des protéines musculaires. Cet effet protecteur serait dû au fait que l’hormone permet préférentiellement l’utilisation d’autres sources d’énergie que les protéines, notamment les acides gras (Ramsay, 2003b)
Finalement, il a été rapporté que des polymorphismes d’ADN présents sur le gène de la leptine, le niveau d’expression de celui-ci de même que la concentration sanguine de l’hormone ont un effet notable sur certains paramètres de qualité de la carcasse et de la viande chez les ovins (Boucher et al., 2006).
Ces effets se font sentir, entre autres, sur le pourcentage de muscle dans la carcasse, la couleur et la teinte du muscle et sur le pourcentage d’eau contenue dans le muscle (Boucher, 2005; Boucher et al., 2006). Ces effets seront discutés plus en détail dans une section ultérieure.
2.3.5.3 Reproduction
Il a été mis en évidence, lors des premiers travaux avec des souris ob/ob et db/db, que les rongeurs présentant un phénotype d’obésité morbide étaient infertiles. Des expériences subséquentes ont également permis l’observation du retour de la fertilité des rongeurs après injections de leptine (Chehab et al., 1996). Ces études laissaient présager un rôle de la leptine dans la reproduction.
Puberté
À la puberté, l’individu acquiert la capacité de se reproduire. Or, des études ont montré qu’une nutrition inadéquate retarde le déclenchement de cette phase du développement et empêche une cyclicité normale chez la femelle (Houseknecht et al., 1998). La faible adiposité, donc la faible quantité de réserves énergétiques, semble empêcher l’initiation du cycle reproducteur. La concentration de leptine dans le sang étant fortement corrélée à la masse adipeuse déposée dans l’organisme (Macajova et al., 2004), celle-ci est donc présente en plus faible quantité chez les individus maigres. Il semble donc que
34
l’organisme ait besoin de l’atteinte d’un seuil de la concentration de leptine pour initier le développement du système reproducteur et l’établissement de la cyclicité chez la femelle à la puberté (Houseknecht et al., 1998).
C’est d’ailleurs à ce stade du développement qu’est observée une augmentation marquée de la concentration sérique de leptine chez la femme, stimulée par les œstrogènes. Le phénomène est plutôt inverse chez l’homme alors qu’une diminution de la concentration sérique de leptine est provoquée par les androgènes (Moschos et al., 2002; Tena-Sempere et Barreiro, 2002). Chez les rongeurs, par contre, le phénomène est inversé, le mâle présentant une concentration sérique de leptine plus élevée que la femelle. Les raisons de cette différence interespèce sont encore mal comprises (Ahima et Osei, 2004).
Des expériences menées chez des souris de 21 jours traitées avec des injections répétées de leptine ont montré une initiation de la puberté plus hâtive. (Ahima et al., 1997; Chehab et al., 1997). Chez ces animaux, l’ouverture du vagin s’est effectuée quelques jours plus tôt que prévu (1-4 jours) et le poids des organes du tractus reproducteur femelle s’est avéré plus élevé (Chehab et al., 1997). La leptine permet le déclenchement de la puberté de souris dont le développement avait été retardé par une sous-alimentation (Zieba et al., 2005) et chez des souris transgéniques surexprimant la leptine à raison de 12X le taux de base (Moschos et al., 2002). L’effet de l’hormone serait quantitatif chez les animaux normaux, c’est-à-dire qu’un seuil doit être atteint pour que l’effet soit observé. Un essai chez des souris transgéniques surexprimant 5X le taux basal de leptine ne s’est soldé par aucun effet sur le déclenchement de la puberté. Il existe une différence de 10X entre le niveau basal de leptine et le pic atteint à la puberté (Chehab et al., 2002).
Il est cependant important de noter que certaines études ont montré que la leptine n’induisait pas une maturation sexuelle plus hâtive chez les rongeurs bien nourris et prévenait partiellement les effets de la malnutrition sur le déclenchement de la puberté (Zieba et al., 2005). Suite aux résultats contradictoires obtenus concernant l’effet de la leptine sur le déclenchement de la puberté, il a été suggéré que l’hormone serait l’un des multiples facteurs nécessaires au processus. Son effet seul ne serait, par conséquent, pas suffisant pour initier la maturité sexuelle (Cheung et al., 2001).
35
Cycle œstral
Le cycle œstral est un processus physiologique comportant des changements dramatiques des concentrations sériques des gonadotrophines, des stéroïdes sexuels et d’autres hormones connues pour interagir avec la leptine (Moschos et al., 2002). Dans cette optique, il est possible que les variations dans la concentration sanguine de leptine au cours des différentes phases du cycle œstral soient liées aux variations des concentrations sériques des différentes hormones impliquées dans le cycle reproducteur chez la femelle. À cet effet, il a été noté que la concentration de leptine atteignait son zénith à la phase lutéale chez l’humain alors que son point le plus bas était atteint au début de la phase folliculaire du cycle ovarien (Hardie et al., 1997).
Ainsi, il a été mis en évidence que la leptine était en mesure de moduler l’activité de l’axe hypothalamo-hypophyso-gonadique (HPG) en modulant de façon positive la fréquence de relâchement de la GnRH et ce, de manière dose-dépendante (Moschos et
al., 2002). De plus, il a été montré in vitro que la leptine, en plus de son effet direct sur la
sécrétion de GnRH au niveau de l’hypothalamus, avait un effet direct sur la sécrétion de LH au niveau de l’hypophyse (Zieba et al., 2005). Des expériences menées in vivo chez le mouton (Nagatani et al., 2000; Henry et al., 2001), le porc (Barb et al., 2004) et la vache (Amstalden et al., 2000; Amstalden et al., 2002; Zieba et al., 2003a) ont permis d’illustrer cet effet de la leptine sur la sécrétion de LH.
Ceci dit, l’espèce, l’état d’avancement de la maturité sexuelle et le statut nutritionnel des animaux sont des éléments déterminants importants de l’effet qu’aura la leptine sur l’axe HPG, particulièrement chez les ruminants, tels que le mouton (Zieba et al., 2005). Pour ce qui est de la FSH, il a été rapporté que la leptine avait aussi un effet positif sur sa sécrétion, mais à une échelle moindre (Moschos et al., 2002).
En ce qui concerne la progestérone et les œstrogènes, des travaux supplémentaires devront être effectués afin d’éclaircir leur lien avec la leptine compte tenu des résultats divergents des différentes études publiées. Plusieurs articles rapportent des corrélations positives entre la concentration de leptine et les concentrations de progestérone et d’œstrogènes lors de la phase lutéale principalement chez l’humain et le mouton (Hardie
et al., 1997; Paolisso et al., 1999; Cella et al., 2000; Kendall et al., 2004). Cependant,
d’autres publications font état d’une absence de lien entre la leptine et les deux types de molécules, chez l’humain (Quinton et al., 1999; Ludwig et al., 2000).