Effets de l'épandage de boues d'alun sur certains paramètres chimiques du sol et sur la croissance de l'orge.
Texte intégral
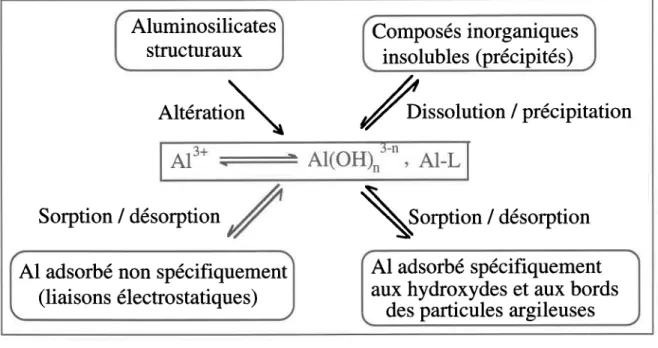
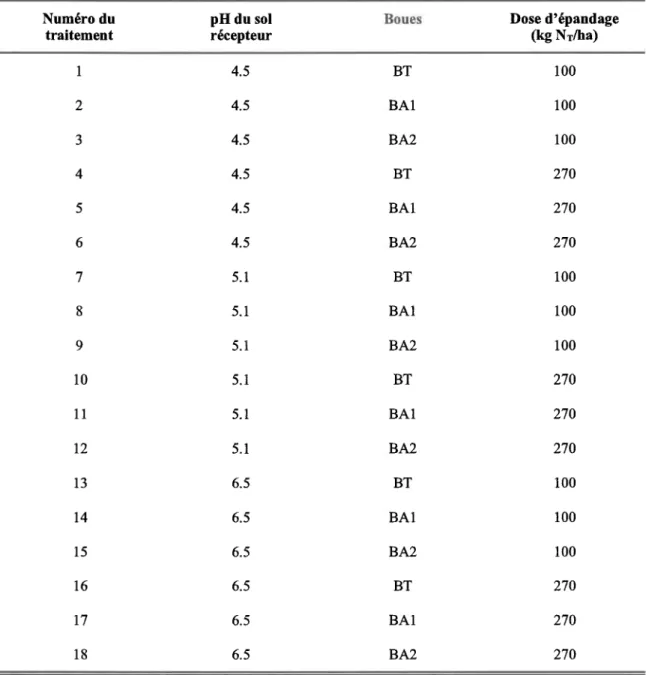
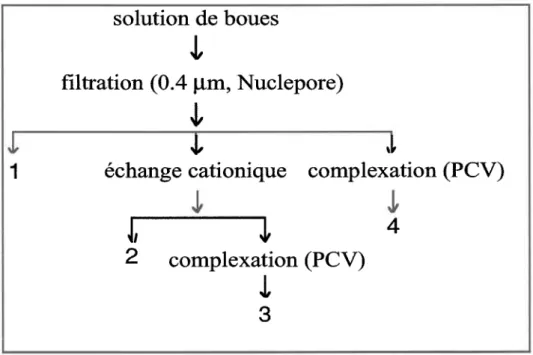
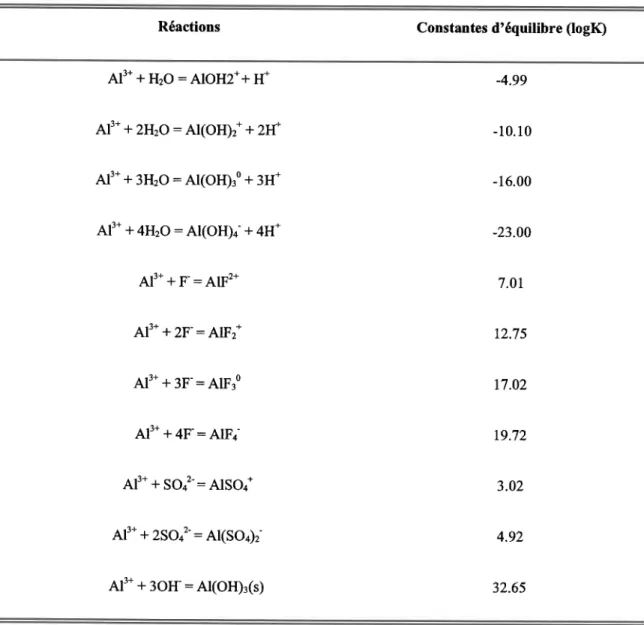
Figure




Documents relatifs
Concerning science, two points need to be better exemplified (in a future edition, if any), i) the place of soil animals in the genesis of the different kind
La gamme de dilution doit être approfondie vers les rapports d'extraction plus élevés (3/1, 4/1).Ne sachant pas si l'effet d'aération (réutiliser le même
Tant que la phase calcaire n’est pas en excès, l’augmentation du pH est reliée aux quantités de carbonate de calcium apportées par une relation qui pour être applicable
L’objectif de notre étude est de délimiter la gamme de pH du sol dans laquelle le test Vicia-micronoyaux peut être réalisé et d’observer l’effet de cette variable sur
Le suivi in situ de la dynamique de l’azote est en accord avec les observations de labo- ratoire sur la grande rapidité de la minéralisation des eaux résiduaires; un sup-
Une quantification des entérobactéries et des résidus d'antibiotique a été réalisée sur les fumiers à la sortie du bâtiment d'élevage et dans le sol pendant 2 mois
Le présent travail dont le thème est «Influence de la nature du sol et de la topographie sur la distribution spatiale de Julbernardia seretii (De Wild) Troupin et de
Sur la colonne 2 sans boue, dés le niveau 2 les quantités d’éléments cumulées exportées diminuent, et ce phénomène est accentué au niveau 3 (graphiques 36(Ni), 38(Ca),
