DYNAMIQUE DE LA PESSIÈRE A MOUSSES AU NORD DU
LAC SAINT-JEAN (QUÉBEC)
Mémoire
Mathieu Frégeau
Maîtrise en biologie
Maître ès sciences (M.Sc.)
Québec, Canada
© Mathieu Frégeau, 2013
iii
Résumé
L’objectif de comprendre la dynamique forestière des peuplements de la pessière à mousses au nord du lac Saint-Jean (Québec). L’intervalle des feux moyen a été mesuré dans des stations représentatives d’une chronoséquence forestière à l’aide de la datation 14C de charbons de bois des sols forestiers. La structure, la date d’établissement des peuplements et la composition botanique des charbons montrent que les forêts étudiées évoluent sous une dynamique de récurrence. Les espèces bien adaptées au feu colonisent les régions ou les feux sont fréquents, tandis que les espèces peu adaptées au feu sont confinées aux stations où les feux sont rares. L’analyse des datations et de la composition des charbons de bois à l’échelle régionale indique que les feux sont présents dans la région depuis au moins 8000 ans et que le régime des feux et la composition des peuplements est demeurée stable au cours des 5000 dernières années.
v
Table des matières
Résumé ... iii
Table des matières ... v
Liste des tableaux ... vii
Liste des figures ... ix
Remerciements ... xi
Introduction ... 1
Objectifs ... 6
Méthodes ... 8
Région d’échantillonnage ... 8
Sélection des stations ... 9
Échantillonnage de la végétation et description des stations ... 11
Échantillonnage des charbons de bois ... 13
Structure des peuplements ... 14
Historique des feux ... 15
Détection de feux régionaux ... 18
Résultats ... 19
Caractéristiques des stations échantillonnées ... 19
Datation des peuplements ... 28
Composition botanique des charbons ... 32
Abondance et composition botanique des charbons de bois au cours du temps ... 35
Estimation de l’erreur induite ... 39
Historique des feux ... 40
Discussion ... 49
Conclusion ... 53
Bibliographie ... 57
vii
Liste des tableaux
Tableau 1 Critères de sélection des stations, nombre de stations recensées et nombre de stations
sélectionnés dans chacune des catégories. PG : pin gris; ÉPN : épinette noire. ... 11
Tableau 2 Caractéristiques générales des stations échantillonnées. ... 19
Tableau 3 Description générale des sols. ... 20
Tableau 4 Fréquence relative (%) des principales espèces d’éricacées, d’herbacées, de mousses et de
lichens des stations échantillonnées. ... 21
Tableau 5 Nombre total de tiges, nombre de tiges >10 cm (DHP), surface terrière (S.T.) et surface terrière
relative (S.T. Rel.) des individus de la strate arborescente des stations échantillonnées. ... 22
Tableau 6 Taux de croissance en hauteur (cm/année) entre la base des arbres et 1,3 m au-dessus du sol
chez les pin gris et les épinettes noires des stations PGEN, ENPG1 et ENPG2. Un test de t a été effectué afin de vérifier si une différence significative existe entre le taux de croissance en hauteur des deux espèces. ... 26
Tableau 7 Taux de croissance en hauteur (cm/année) des tiges de pin gris et d'épinette noire des stations
PGEN, ENPG1, ENPG2. Un test de t a été effectué afin de vérifier si une différence significative existe entre le taux de croissance des deux espèces. ... 26
Tableau 8 Année d’établissement du pin gris et de l'épinette noire dans les stations PGS, PGEN, ENPG1,
ENPG2. ... 28
Tableau 9 Datation des feux les plus récents survenus dans chaque station. ... 29 Tableau 10 Nombre de macrorestes ligneux d’épinette et de pin dans les stations ENPG1, ENPG2 et PGEN.
Le chiffre entre parenthèses correspond au nombre de restes ligneux sans souche. ... 31
Tableau 11 Composition botanique des charbons de surface des stations échantillonnées. ... 33 Tableau 12 Composition botanique des charbons du sol minéral des stations échantillonnées. ... 34
ix
Liste des figures
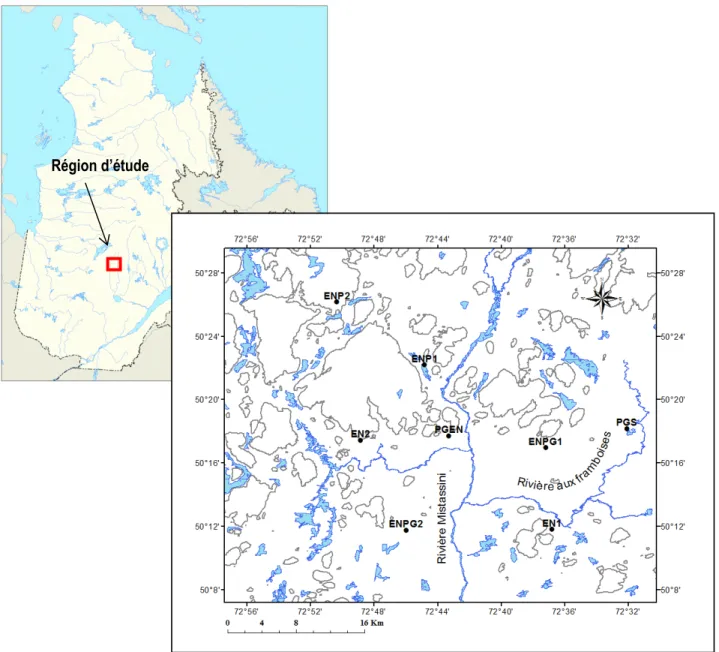
Figure 1 Zones de végétation du Québec. Modifié de Payette et Bouchard (2001). ... 1 Figure 2 Localisation de la région d’étude et des stations. En bleu : les principaux cours d’eau de la région. La
courbe de niveau de 500 m est représentée par une ligne grise (MRNF 2000). ... 8
Figure 3 Nombre de tiges/ha de plus 10 cm au DHP de Pinus banksiana (en gris) et de Picea mariana (en
noir) dans chaque station. ... 23
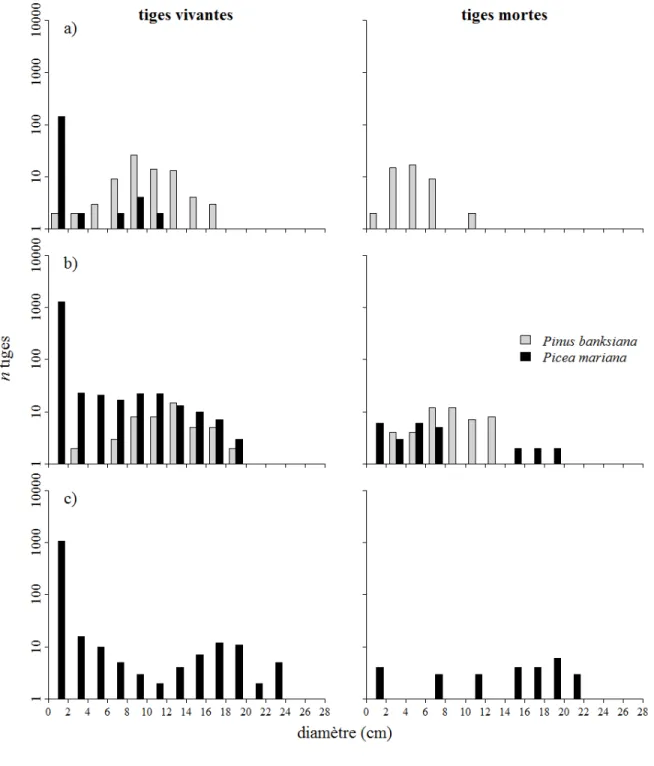
Figure 4 Structure de taille (DHP) des tiges vivantes et mortes du pin gris (en gris) et de l’épinette noire (en
noire) dans les stations PGS (a), ENPG2 (b) et EN2 (c). Les tiges sont regroupées en classes de 2 cm. Le nombre de tiges est additionné de 1 pour permettre la représentation logarithmique. ... 24
Figure 5 Hauteur des tiges de pin gris (ligne hachurée) et d’épinette noire (ligne continue) en fonction du
temps dans les stations PGEN (a), ENPG2 (b) et ENPG1(c). ... 27
Figure 6 Fréquence des tiges de Picea mariana (en noir), de Pinus banksiana (en gris) et d’Abies balsamea
(en blanc) en fonction du temps chez les stations échantillonnées. ... 30
Figure 7 Nombre total de charbons de bois datés dans toutes les stations en fonction du temps (classe de 150
ans). (Total), en surface (Surface) et enfouis dans le sol minéral (Minéral). ... 36
Figure 8 Résidus de la régression log-linéaire de l'abondance des charbons de bois de moins de 5000 ans BP
étal. en fonction du temps (classe de 150 ans ... 37
Figure 9 Nombre de charbons (surface et minéral) de Picea sp. (en noir), de Pinus banksiana (en gris) et
d’Abies balsamea (en blanc) en fonction du temps. Les dates sont regroupées en classes de 500 ans. ... 38
Figure 10 Distribution de probabilité cumulée des dates 14C des charbons (surface et minéral) de chacune des stations. Les symboles + correspondent à des évènements de feu recensés à l’aide des dates 14C des charbons. ... 42
Figure 11 Historique des feux couvrant les 8000 dernières années établi à l’aide des charbons de bois de
surface et du sol minéral. Colonne de gauche : Distribution cumulée des feux, nombre de feux (n) et période de feu (P). Colonne du centre : Nombre d’intervalle en fonction de leur durée par classe de 100 ans et intervalle de feu moyen (Im). Colonne de droite : Courbe d’accumulation, nombre de feux estimés (ne) et intervalle de feux moyen estimé (Ime). ... 43
Figure 12 Historique des feux couvrant les 1500 dernières années établi à l’aide des charbons de bois de
surface et du sol minéral. Colonne de gauche : Distribution cumulée des feux, nombre de feux (n) et période de feu (P). Colonne du centre : Nombre d’intervalle en fonction de leur durée par classe de 100 ans et intervalle de feu moyen (Im). Colonne de droite : Courbe d’accumulation, nombre de feux estimés (ne) et intervalle de feux moyen estimé (Ime). ... 44
x
Figure 13 Intervalle de feu moyen calculé à partir de l'historique complet (a) et de l'historique des 1500
dernières années BP étal.. (b). En gris foncé : intervalle de feu moyen mesuré, en gris pâle : intervalle de feu moyen estimé. ... 45
Figure 14 Superficie approximative des grands feux ayant affecté plusieurs stations. Le nombre associé à
l’identification de la station correspond à la date d’un feu en années BP étal. Les contours des grands feux (ligne noire) ont été tracés à l’aide de la localisation des dates des feux. En bleu : les principaux cours d’eau de la région. La courbe de niveau de 500 m est représentée par une ligne grise (MRNF 2005). ... 47
xi
Remerciements
J’aimerais tout d’abord remercier mon directeur de maîtrise, Serge Payette. Il a su me transmettre sa passion pour l’écologie, sa rigueur et son esprit scientifique tout au long des diverses étapes de ma maîtrise. Les connaissances et les méthodes que j’ai acquises auprès de lui me seront utiles durant l’ensemble de mon parcours professionnel.
Je remercie aussi Pierre-Luc Couillard, Mélanie Veilleux Nolin, Marianne White, Mélanie Jean, Alexandre Truchon Savard, Vanessa Pilon, Francis St-Amour en tant que membre et ex-membre du laboratoire de Serge Payette. L’esprit d’entraide et la bonne humeur qui règnent dans ce laboratoire ont su forger des souvenirs inoubliables de cette période de ma vie. J’aimerais aussi remercier les membres du LaBoudreau dont la collaboration avec notre laboratoire a été particulièrement enrichissante. Mention spéciale au locataire du local 4052, Marie-Pier Denis, Marianne White et Mélanie Jean, pour leur appui et les nombreuses conversations animées qui ont comblé certaines fins d’après-midi.
Je remercie aussi toute ma famille et mes amis, qui m’ont appuyé lors de ce fastidieux parcours qu’est la maîtrise. J’aimerais remercier particulièrement ma compagne, Jacinthe Bédard Lacroix, pour son appui et son enthousiasme qui m’ont beaucoup aidé à traverser les hauts et les bas de ce projet.
Je remercie Marianne Ricard, Catherine Barthe et Karine Champagne d'avoir supporté les longues heures de terrain et de travaux en laboratoire que je leur infligeais. Je remercie aussi Ann Delwaide pour son aide lors des travaux de laboratoire.
Je remercie également le MRN, notamment Pierre-Luc Couillard, Pierre Grondin et Jocelyn Gosslin et Produits forestiers Résolu (Serge Gosselin, François Levesque) pour leur soutien technique et leurs
propositions sur les orientations données au projet.
Enfin, je remercie le CRSNG, le FQRNT, le programme de bourses en milieu de pratique BMP innovation, la CRÉ du Saguenay-Lac-Saint-Jean et le MRN pour leur soutien financier.
1
Introduction
La forêt boréale nord-américaine, caractérisée par la dominance des conifères, est divisée en trois zones selon la densité des arbres : la forêt fermée, la forêt ouverte et la toundra forestière (Rowe 1972, Hare et Ritchie 1972, Payette 1992, Payette et Bouchard 2001). La pessière à mousses, une zone de végétation
Figure 1 Zones de végétation du Québec. Modifié de Payette et Bouchard (2001). Érablière
Forêt mixte
Forêt coniférienne fermée
Forêt coniférienne ouverte
Sapinière Pessière à mousses Pessière à lichens Toundra forestière Toundra Toundra arbustive Toundra herbacée
2
dominée par l’épinette noire (Picea mariana (Mill.) B.S.P.), se situe entre le 49 °N et le 52 °N, de la baie de James jusqu’à l’est du Labrador (Saucier et al. 2009) (Figure 1). Le feu est la principale perturbation de la pessière à mousses, lequel exerce une grande influence sur la composition et la structure de la forêt (Payette et al. 1989, Payette 1992). Les épidémies d’insectes (Morin 1998, Bouchard et Pothier 2010) et les chablis (Bouchard et al. 2009) affectent aussi cette forêt, bien qu'ils aient un effet moindre que les feux.
L’influence des feux sur le paysage forestier peut être décrite grâce au concept de régime des feux qui comprend la fréquence, l’étendue et la sévérité des feux (Heinselman 1981, Agee 1996, Payette 2010). La fréquence des feux est le nombre de feux survenus en un point donné au cours d’une période de temps donnée. L’intervalle de feu moyen est la durée moyenne en année entre tous les feux survenus en un point donné. La période de rotation correspond au temps nécessaire pour que la surface d’une région donnée brûle au moins une fois dans son entièreté (Heinselman 1973). L’étendue d’un feu comprend la superficie et la forme de la surface brûlée. La sévérité d’un feu décrit l’impact écologique des feux sur la station et le peuplement, par exemple le taux de mortalité des arbres ou la quantité de matière organique brûlée.
Le climat est le facteur écologique ayant la plus grande influence sur le régime des feux (Agee 1996). Les précipitations et la température sont parmi les principaux facteurs déterminants de la fréquence des feux dans une région donnée (Turner et Romme 1994, Romme 2005). Une température élevée et de faibles précipitations assèchent les matériaux combustibles (bois au sol, matière organique, litière) et favorisent l’allumage et la propagation des feux (Van Wagner 1983, Renkin et Despain 1992). L’influence du climat peut-être constaté en comparant la période de rotation des feux entre des régions au climat différent. Avec moins de 900 mm de précipitation par année, le régime des feux des régions de la baie de James et de l’Abitibi se caractérise par une période de rotation élevée d’environ 100 à 150 ans (Cogbill 1985, Payette et al. 1989, Bergeron et al. 2001, Parisien et Sirois 2003). À l’opposé, des précipitations annuelles allant de 1000 à 1100 mm dans l’est du Québec favorisent une plus longue période de rotation, pouvant dépasser les 500 ans (Foster 1983, Bouchard et al. 2008).
Le concept de dynamique forestière décrit les changements directionnels et cumulatifs de la structure et de la composition d’un peuplement forestier au cours du temps (Drury et Nisbet 1973, Finegan 1984, Pickett et al. 1987). La séquence des espèces est déterminée par la modification graduelle de l’humidité, de la luminosité, de la disponibilité des nutriments et des interactions entre les espèces (Odum 1969, Connell et Slatyer 1977). La trajectoire d’une dynamique forestière est en partie tributaire du type sol et de la topographie (Urban et al. 1987, Turner et al. 2001). La disponibilité en eau et en nutriments, qui est liée en partie au type
3
de sol et à la topographie, détermine quelles espèces sont capables de croître dans un milieu et donc susceptibles de participer à la succession (Oliver et Larson 1996, Bridge et Johnson 2000). Le régime des feux influence la composition spécifique du paysage en fonction de la capacité des espèces forestières à résister au feu ou pour coloniser rapidement la station après le feu (Heinselman 1981, Rowe 1983, Turner et al. 2001). Le type de dynamique est aussi tributaire du temps écoulé entre les perturbations (Oliver 1980, Romme et Knight 1981). Par exemple, dans les biomes où le temps écoulé moyen entre les perturbations est court, la séquence successionnelle pourra rarement atteindre les stades de fin de succession. Ainsi, les peuplements âgés et matures seront rares dans le paysage.
Le régime de feux influence la structure d’âge des peuplements dans une région donnée (Whittaker et Levin 1977, Turner et al. 2001). La mosaïque forestière dans une région donnée comprend de jeunes et de vieux peuplements régénérés après feu ainsi que des peuplements non affectés par les feux. L’action récurrente des feux sur la forêt crée une diversité de peuplements d’âge et de composition variables.
Trois types de dynamique sont observés en forêt boréale, c’est-à-dire la dynamique de récurrence, la dynamique de succession et la dynamique d’états alternatifs stables. Chaque type est associé à une position topographique et à un régime de feu particulier. La dynamique de récurrence se caractérise par l’établissement rapide après feu d’un peuplement de composition spécifique semblable au peuplement qui a subi la perturbation (Frelich 2002, Johnstone et al. 2004, Kurkowski et al. 2008). Les peuplements sous une dynamique de récurrence présentent peu de changement dans la composition au cours du temps, si ce n’est un changement dans la dominance du peuplement due à la croissance différentielle et à la compétition entre les espèces (Heinselman 1973, 1981). Ce type de dynamique est associé à des espèces capables de coloniser rapidement la station après feu. Par exemple, le pin gris (Pinus banksiana Lamb.) et l’épinette noire s’installent rapidement après feu grâce à un réservoir aérien servant à disperser les graines à la suite d’un feu (Cayford et McRae 1983, Rowe 1983). Certaines espèces comme le bouleau à papier (Betula papyrifera Marsh.) et le peuplier faux-tremble (Populus tremuloides Michx.) peuvent aussi recoloniser rapidement une station par multiplication végétative (Heinselman 1981, Rowe 1983). Ce type de dynamique est caractéristique des régions et des stations où l’intervalle de feu moyen est relativement court (Heinselman 1973, Oliver 1980). Le temps écoulé entre deux perturbations est généralement trop court pour que les peuplements atteignent le stade final d’une succession. Par exemple, dans la région de la baie de James, où l’intervalle moyen entre les feux est inférieur à 150 ans, les peuplements âgés qui ont atteint le stade de fin de succession sont rares (Carleton et Maycock 1978).
4
La dynamique de succession est définie comme l’établissement d’une espèce pionnière après une perturbation laquelle est graduellement remplacée par d’autres espèces plus compétitives et tolérantes (Frelich 2002). Le peuplement de fin de succession est supposé stable sur le plan de la structure et de la composition. En forêt boréale, ce type de dynamique est caractéristique des régions et des stations où les perturbations par le feu sont peu fréquentes. Ainsi, comme le temps écoulé entre deux perturbations est long, il est possible d’observer l’établissement graduel de nouvelles espèces dans la station. Dans les sapinières de l’est du Canada, ce type de dynamique est caractérisé par l’établissement d’espèces intolérantes à l'ombre après feu, comme le bouleau à papier, suivi par l’augmentation de l’abondance d’espèces plus tolérantes comme le sapin baumier (Abies balsamea (L.) Mill.) (Bergeron 2000, Lesieur et al. 2002, Couillard et al. 2012).
Chez la dynamique d’états alternatifs stables, la végétation subit, par exemple, des perturbations successives et rapprochées causant une réduction sensible de la régénération. Le peuplement ainsi formé est moins dense que celui qui se développait avant la perturbation. La structure du nouveau peuplement crée des conditions locales qui empêchent le retour, du moins à moyen terme, du peuplement d’origine et favorise ainsi la création d’une végétation correspondant à un état alternatif stable. Ce type de dynamique peut être associé à un changement du régime de perturbation, comme par exemple, une diminution de l’intervalle de feu moyen ou un effet cumulé de plusieurs perturbations. Un exemple de végétation à état alternatif stable est celui d’une pessière à mousses affectée par une régénération déficiente à la suite d’une épidémie d’insectes rapidement suivie par un feu (Payette et al. 2000, Jasinski et Payette 2005, Simard et Payette 2005). La faible densité du nouveau peuplement stimule la croissance du lichen, lequel ne favorise pas la survie des semis d’épinette noire, empêchant ainsi la densification du peuplement. Des feux rapprochés dans les peuplements d’épinette noire de l’Alaska ont causé également une diminution de la régénération, les épinettes étant trop jeunes pour produire un grand nombre de graines viables lors de la perturbation (Johnstone et Chapin 2006). L’absence de régénération a favorisé l’établissement de peuplements de bouleau à papier de l’Alaska (Betula neoalaskana Sarg.) (Johnstone et al. 2010). La régénération déficiente à la suite d'un feu est un phénomène répandu dans plusieurs régions du Québec, ce qui favorise l’expansion de la pessière à lichens (Girard et al. 2008).
Un aspect important de l’étude du lien entre la dynamique forestière et la fréquence des feux est la mesure adéquate de cette dernière. Une méthode appropriée du calcul de la fréquence des feux nécessite à la fois une mesure précise de la superficie des feux et de leur fréquence sur une longue période de temps (Payette 2010). Cette méthode permet de définir les variations du régime de feu dans le temps et d’évaluer l’importance des variables topographiques (pente, sol, altitude, présence de coupe-feu) sur le régime des feux.
5
Deux principales méthodes sont utilisées pour décrire la fréquence des feux : la datation et la cartographie des feux à l’aide des cernes de croissance des arbres et l’analyse des charbons de bois dans les lacs. La datation et la cartographie des feux consiste à déterminer l’année d’établissement des arbres et de cicatrices de feu dans le but de confectionner des cartes de l’âge et de l’étendue des feux (Heinselman 1973, Arno et al. 1977, Payette 2010). Cette méthode permet de confectionner des cartes au contour de feu précis spatialement, mais ne permet pas de reconstituer l’historique des feux sur une période plus longue que la durée de vie maximale des arbres de la région (Conedera et al. 2009, Payette 2010), période qui ne dépasse guère les 300 ans dans la pessière à mousses. Il est donc difficile de connaître les changements de la fréquence des feux dans le temps et elle ne peut être utilisée si la fréquence moyenne dépasse la durée de vie des arbres.
La seconde méthode de mesure de la fréquence des feux est l’analyse des charbons de bois dans les lacs (Clark 1990, Millspaugh et Whitlock 1995, Carcaillet et al. 2001). Cette méthode consiste à mesurer l’abondance des charbons de bois dans la colonne sédimentaire d’un lac. L’abondance des charbons d’une taille de 125-250 µm dans la colonne sédimentaire est considérée comme un indicateur de feux locaux (Whitlock et Larsen 2001). Cette méthode permet d’estimer localement ou régionalement l’activité des feux sur une longue fenêtre temporelle. Par contre, il est plutôt difficile d’identifier des feux locaux à l’aide de l’abondance des charbons (Higuera et al. 2010). De plus, la méthode ne permet pas d’évaluer la surface de chaque feu, étant donné que, par définition, la distance des feux locaux varie de 500 m jusqu'à 3 km autour du site d’échantillonnage (Clark 1988, Higuera et al. 2011).
La méthode utilisée dans le cadre de la présente étude est la datation de charbons de bois trouvés dans les sols forestiers (Talon et al. 2005, de Lafontaine et Payette 2012). Cette méthode consiste à dater au 14C les charbons que l’on trouve dans les fractions organique et minérale des sols forestiers. Elle permet de dater des feux anciens, dont certains ayant eu lieu immédiatement la dernière glaciation (Talon et al. 2005). Cette méthode a aussi comme avantage d’utiliser des preuves concrètes de l’existence de feux in situ étant survenus dans la station d’échantillonnage grâce à la présence de charbons macroscopiques qui ne peuvent être transportés sur de grandes distances. Enfin, la méthode permet d’identifier botaniquement les charbons de bois, ce qui permet d’aborder la question de la dynamique de régénération des espèces arborescentes après feu.
6
Objectifs
L’étude de la dynamique forestière comprend l’analyse de la composition et de la structure des peuplements forestiers à plusieurs moments au cours de la succession. Puisque la succession forestière de la pessière à mousses peut prendre plus de 100 années à se réaliser (Heinselman 1981, Viereck 1983, Payette 1992), on ne peut pas observer directement les changements de composition et de structure de la végétation au cours du temps. L’étude de la succession s’effectue plutôt en étudiant la chronoséquence après feu. La chronoséquence correspond à un ensemble de peuplements croissant dans les mêmes conditions topographiques et édaphiques alignés le long d’un gradient temporel exprimés par différents stades successionels (Pickett 1989, Johnson et Miyanishi 2008). La méthode de la datation des charbons bois des sols forestiers est complémentaire à l’étude des chronoséquences forestières. La datation au 14C des charbons de bois permet de reconstituer l’historique des feux d’un peuplement sur une longue période (Payette et al. 2012). Ainsi, il est possible de calculer un intervalle de feu moyen dans une station donnée et déterminer s’il est lié au type de dynamique du peuplement (récurrence, succession ou états alternatifs stables). L’identification botanique des charbons permet d’évaluer la composition botanique relative de la station au cours du temps. Il est donc possible de déterminer si les changements dans la composition des charbons sont liés au régime des feux.
Trois objectifs principaux sont visés dans cette étude. Le premier objectif est de tester l’hypothèse de l’existence d’une dynamique de succession dans la pessière à mousses. Cette hypothèse sera testée en déterminant l’état démographique, c’est-à-dire déterminer si la population arborescente est en expansion, à l’équilibre ou en extinction (Payette 2010), et en datant l’établissement de peuplements à différents stades successionnels le long d’une chronoséquence théorique. Cette dernière débute par l’établissement du pin gris après feu. Par la suite, l’épinette noire s’installe graduellement dans le peuplement, formant ainsi un peuplement mixte de pin gris et d’épinette noire. Le pin gris disparaît graduellement et est remplacé par l’épinette noire au cours du temps. Le peuplement monospécifique d’épinette noire constitue le stade final de la chronoséquence, où l’espèce est capable de se régénérer en l’absence de perturbation.
Le second objectif est de tester l’hypothèse que l’intervalle de feu moyen varie à l’intérieur de la région étudiée en fonction des facteurs topographiques et édaphiques. Pour ce faire, l’intervalle des feux moyen est calculé chez des peuplements forestiers qui diffèrent par le type de sol, l’altitude et la pente à l’aide de la datation au 14C des charbons de bois des sols forestiers.
7
Le dernier objectif est de tester l'hypothèse voulant que la fréquence des feux constitue un outil important servant à déterminer le type de dynamique (récurrence, succession ou état alternatif stable) dans un peuplement forestier. Ainsi, les variations de la fréquence des feux en fonction des facteurs topographiques et édaphiques devraient rendre compte de l’existence de plusieurs types de dynamique au sein d’une même région.
8
Méthodes
Région d’échantillonnage
La région d’étude se situe à 250 km au nord du Lac-Saint-Jean, à quelque 100 km au sud-est du lac Mistassini. Les stations d’étude se trouvent à l’intérieur d’une aire de 700 km2 entre les latitudes 50° 10’ et 50°
Figure 2 Localisation de la région d’étude et des stations. En bleu : les principaux cours d’eau de la
région. La courbe de niveau de 500 m est représentée par une ligne grise (MRNF 2000).
9
28’ N et entre les longitudes 72° 32’ et 72° 51’ O (Figure 2). Le relief de la région est formé de basses collines culminant à environ 600 m d’altitude. Le sol est en grande partie composé de sable loameux d’origine glaciaire sur les collines et de sable d’origine fluvio-glaciaire le long des principales rivières. La station météorologique Bonnard, située à 120 km au nordest de la région d’étude, enregistre une température moyenne annuelle de -1,8 °C et des précipitations moyennes annuelles de 950 mm, dont environ 30 % tombent sous forme de neige (Environnement Canada 2012).
L’épinette noire est l’espèce la plus abondante de la région où elle occupe tous les types de dépôt et toutes les positions topographiques. Le pin gris est présent généralement dans des peuplements mixtes avec l’épinette noire sur les sols composés de sable loameux, mais il forme des peuplements purs sur les dépôts sableux. Le sapin baumier et le bouleau blanc sont présents sur les versants de colline et ils sont accompagnés occasionnellement par l’épinette blanche (Picea glauca (Moench) Voss.). Le peuplier faux tremble et le mélèze laricin (Larix laricina (Du Roi) Koch.) sont aussi présents, mais ils forment rarement des peuplements monospécifiques. La strate arbustive est principalement constituée de Rhododendron
groenlandicum (Oeder) Kron et Judd., de Kalmia angustifolia L. et de Vaccinium angustifolium Ait.. Les
mousses comme Pleurozium schreberi (Brid.) Mitt., Ptilium crista-castrensis (Hedw.) de Not., Hyloconium
splendens (Hedw.) B.S. et Sphagnum sp. forment des tapis recouvrant le sol dans la plupart des peuplements.
On trouve aussi plusieurs espèces de lichens dont Cladonia rangiferina (L.) F.H.Wigg. et Cladonia stellaris (Opiz) Brodo dans les milieux secs et ouverts. L’exploitation forestière ayant débuté dans la région au début des années 2000, l’influence de l’homme sur la composition et la structure du paysage est négligeable.
Sélection des stations
La sélection des stations servant à tester l’hypothèse de la chronoséquence est basée sur la méthode de l’observation de la végétation de peuplements adjacents, établis sur des sols semblables, et jugés à différents stades de succession (Pickett 1989, Johnson et Miyanishi 2008). Cette méthode consiste à décrire des peuplements alignés sous forme de stades successionnels le long d’une chronoséquence théorique, dans des conditions édaphiques similaires, en supposant qu’ils suivent tous la même dynamique. Ainsi, la différence de composition et de structure dans les peuplements devrait être due uniquement au temps écoulé depuis la dernière perturbation. En second lieu, des peuplements situés le long d’une toposéquence (différentes positions topographiques et différents types de sol) ont été sélectionnés pour mesurer l’influence de la physiographie sur le régime des feux.
10
La structure, la composition botanique, le type de sol et la topographie de 27 stations ont été décrits. À l’aide de ces observations, trois catégories de position topographique et édaphique regroupant des peuplements de la pessière à mousses ont été établies. La première catégorie concerne les épandages fluvio-glaciaires sableux le long des rivières. La végétation de cette catégorie comprend une forte proportion de pinèdes à pin gris. La deuxième catégorie regroupe les peuplements sur dépôt glaciaire sablo-loameux de moins de 10° de pente. Dans cette catégorie, les peuplements sont formés de pin gris et d’épinette noire en proportion variable. Le sol des peuplements de la troisième catégorie est formé de sable loameux d’origine glaciaire dont la pente est supérieure à 15°. Dans cette catégorie, les peuplements sont surtout des pessières à épinette noire comprenant moins de 15 % de sapin baumier.
Des stations ont été sélectionnées pour représenter trois stades de succession différents le long de la chronoséquence théorique. Tous ces peuplements sont situés dans la catégorie topographique et édaphique dont le sol est un sable loameux et dont la pente est inférieure à 10°. Le stade « début de succession » est composé de peuplements dominés par le pin gris accompagné par l’épinette noire. Le stade « milieu de succession » est composé de peuplements dominés par l’épinette noire accompagnée par le pin gris. Le stade « fin de succession » est composé de peuplements constitués uniquement d’épinette noire.
Les stations recensées ont été classées dans chacune des catégories selon le type de sol, la situation topographique et la composition de la strate arborescente. Par la suite, une ou deux stations par catégorie et par stade de succession ont été sélectionnées aléatoirement (Tableau 1).
11 Tableau 1 Critères de sélection des stations, nombre de stations recensées et nombre de stations
sélectionnés dans chacune des catégories. PG : pin gris; ÉPN : épinette noire.
Catégories % PG % EPN Texture du sol Pente (°) recensées N sélectionnées N
Pinède à pin gris (PGS) > 75 < 25 Sable < 10 3 1
Pinède à pin gris et à
épinette noire (PGEN) > 50 < 50
Sable
loameux < 10 7 1
Pessière à épinette noire
et à pin gris (ENPG) < 25 > 75
Sable
loameux < 10 5 2
Pessière à épinette noire
(EN) 0 100
Sable
loameux < 10 5 2
Pessière à épinette noire
en pente (ENP) 0 100
Sable
loameux > 15 3 2
Échantillonnage de la végétation et description des stations
L’échantillonnage a été effectué dans une placette de 10 m par 50 m située à un minimum de 50 m de la bordure du peuplement et parallèle à la pente. Avant d’installer la placette, le peuplement était examiné pour s’assurer qu’elle était représentative du peuplement. Par la suite, un ruban gradué de 20 m a été disposé au hasard sur le sol. Un tirage au sort a été effectué afin de déterminer si les premiers ou les derniers 10 m de la ligne seraient utilisés comme extrémité de la placette. Le segment choisi a servi de base pour placer les segments de 50 m de manière à former une placette rectangulaire.
La première partie de l’échantillonnage a consisté à caractériser les variables physiques, la composition botanique, l’état démographique et la date d’établissement du peuplement. Une fois la végétation de chaque station inventoriée, on s’est assuré que chacune des stations, présumées à différents stades de la chronoséquence théorique, occupe le même type de sol et la même position topographique. L’état
12
démographique de chaque peuplement a aussi été décrit (expansion, extinction ou équilibre) afin d’en déterminer la dynamique successionnelle réelle.
L’état démographique des peuplements a été décrit en mesurant la structure de taille des arbres de chaque station. Tous les arbres et les troncs au sol à l’intérieur de la placette ont été identifiés à l’espèce et le diamètre à hauteur de poitrine a été mesuré (DHP : 1,3 m). L’état de l’arbre (vivant ou mort) a aussi été pris en note de même que l’origine des individus (plantule ou marcotte) chez les individus ≤ 2 cm DHP.
L’année d’établissement de tous les peuplements a été déterminée en datant 10 arbres dominants de la station. Chaque arbre a été abattu et une rondelle a été prélevée le plus près possible du sol. Pour déterminer précisément l’année du dernier feu, les alentours des stations ont été explorés à la recherche d’arbres portant des cicatrices de feu. Ces derniers ont été abattus et une rondelle a été prélevée. La présence d’épinette noire surcimée dans des peuplements de pin gris a été présentée comme un indice de succession d’un peuplement de pin gris vers un peuplement d’épinette noire (Carleton et Maycock 1978). Par contre, la formation de strate dans les peuplements constitués de ces deux espèces serait due à une croissance différentielle (Heinselman 1973, 1980, Bergeron et Dubuc 1988). Pour tester cette hypothèse, cinq arbres des deux espèces dominantes ont été échantillonnés chez les peuplements mixtes PGEN, ENPG1, ENPG2 (figure 2). Le taux de croissance en hauteur des pins gris et des épinettes noires échantillonnés précédemment a été déterminé chez les peuplements mixtes (PGEN, ENPG1 et ENPG2) à l’aide d’une analyse de tige. Des rondelles ont été prélevées à la base de l’arbre, à 1,3 m et à 25 %, 50 % et 75 % de la hauteur entre l’apex et 1,3 m. Toutes les rondelles servant à la datation des arbres et des cicatrices de feux, ainsi qu’à l’analyse de tige ont été finement poncées et les cernes ont été dénombrés sous une loupe binoculaire
Un relevé de végétation a été effectué selon la méthode du point d’intersection (Muller-Dombois et Ellenberg 1974) le long d’une ligne de 50 m correspondant à l’un des deux axes de 50 m délimitant la placette d’échantillonnage. La présence de toutes les espèces vasculaires et invasculaires touchant la pointe d’une tige métallique déployée verticalement a été prise en note à un intervalle de 50 cm le long de la ligne de 50 m. La fréquence relative de chaque espèce correspond au rapport entre la fréquence (en %) de l’espèce dans l’ensemble des points de mesure et la somme des fréquences (en %) de toutes les espèces.
Dans chaque station, la texture de l’horizon B a été évaluée par diffraction laser (Horiba Scientific). L’épaisseur des horizons organiques F, FH et H a été mesurée dans 25 microstations disposées tous les 5 m sur le pourtour de la placette d’échantillonnage et en son centre. Trois échantillons de l’horizon F ont été
13
récoltés aux mètres 0, 5 et 15 de la ligne d’échantillonnage de 50 m et ramenés au laboratoire pour mesurer le pH.
Les espèces arborescentes présentes avant le dernier feu ont été identifiées et dénombrées à l’aide des macrorestes ligneux trouvés à la surface du sol des peuplements mixtes (PGEN, ENPG 1 et 2). L’objectif de cet échantillonnage était de savoir si les espèces arborescentes du peuplement avant feu étaient les mêmes que celles du peuplement après feu. Tous les macrorestes issus de souches situés à l’intérieur de la parcelle ont été récoltés. Chaque macroreste ligneux a été examiné quant à la présence de traces de feu (surface noircie ou présence de charbons). L’identification botanique des pièces de bois, au genre ou à l’espèce lorsque possible, a été faite à l’aide des caractéristiques anatomiques du bois (Panshin et deZeeuw 1980).
Échantillonnage des charbons de bois
L'analyse des charbons présents dans les sols forestiers est basée sur l’enfouissement des charbons dans le sol après feu ou après une perturbation affectant la surface du sol. Les perturbations du sol par le déracinement des arbres suivant un chablis (Brown et Martel 1981, Bormann et al. 1995) incorporent la matière organique de surface et les charbons qu’elle contient dans le sol. L’enfouissement aléatoire des charbons au cours du temps ne favorise pas la stratification des charbons dans le sol en fonction de leur âge (Carcaillet 2001, Talon et al. 2005, Fesenmyer et Christensen 2010). Une fois enfouis, les charbons sont très résistants à la dégradation et peuvent perdurer plusieurs milliers d’années (de Lafontaine et al. 2011). Les charbons de plus 2 mm de diamètre sont généralement d’origine locale (Ohlson et Tryterud 2000).
Les charbons ont été échantillonnés selon la méthode décrite par Talon et al. (2005) et de Lafontaine et Payette (2011). Ils ont été récoltés dans 25 microplacettes à chaque 5 m d’intervalle sur le pourtour de la placette d’échantillonnage ainsi qu’en son centre. À chaque microplacette, la matière organique a été enlevée sur une surface d’environ 40 cm par 40 cm afin de dégager la surface du sol minéral. Trois ou quatre charbons situés à l’interface du sol minéral et de l’horizon organique (dénommés charbons de surface) ont été récoltés manuellement. Par la suite, la surface de l’horizon minéral a été nettoyée de toute trace de matière organique. Une carotte de sol minéral de 15 cm de haut et de 8 cm de diamètre (environ 750 cm3) a été prélevée à l’aide d’une sonde de sol.
En laboratoire, le sol minéral a été mis en suspension dans une solution de NaOH 1 % pendant au moins 12 heures de manière à disperser les agrégats de sol. Le sol a été tamisé à l’aide de tamis superposés
14
de 4 mm et de 2 mm sous un jet d’eau. Les charbons de surface et ceux extraits des tamis ont été nettoyés et pesés. Tous les charbons de plus de 5 mg (exceptionnellement entre 3 et 5 mg, lorsque le nombre de charbons de 5 mg était insuffisant) ont été identifiés au genre ou à l’espèce lorsque possible à l’aide des caractéristiques anatomiques du bois à l’aide d’une clef d’identification (Panshin et deZeeuw 1980) et de la collection de bois carbonisé du Centre d’études nordiques (Université Laval, Québec). Les espèces des genres Picea et Betula ainsi que les genres Picea, Abies et Larix sont difficiles à reconnaître lorsque les pièces sont petites. En cas de doute, les pièces ont été classées comme conifère, feuillu ou indéterminé. Vingt-cinq charbons provenant de la surface du sol et 20 charbons enfouis dans le sol minéral par station ont été datés à l’aide de la méthode de datation au radiocarbone de spectrométrie de masse par accélération (SMA). Le choix des charbons a été effectué de manière aléatoire, en s’assurant que toutes les espèces arborescentes recensées étaient représentées et provenaient de microplacettes différentes. Quelques charbons identifiés comme conifère, feuillu ou indéterminé ont été également datés. Les prétraitements pour la datation au 14C des charbons ont été effectués au Centre d’études nordiques (Université Laval) et les charbons ont été datés au Keck Carbon Cycle AMS Facility (University of California, Irvine, Californie, USA). Les dates 14C ± 2σ ont été étalonnées à l’aide du logiciel CALIB 6.1.0 (Stuiver et al. 2005) et de la courbe d’étalonnage IntCal09 (Reimer et al. 2009). Les datations sont toutes présentées en année BP étalonnée, où 0 BP étalonnée équivaut à l’année 1950 après J.-C.
Structure des peuplements
La structure de taille des espèces arborescentes de chaque peuplement a été décrite sous forme d’histogramme, les tiges étant regroupées en classe de 2 cm. La forme de la distribution permet de distinguer les populations en expansion, en extinction et en équilibre (Payette 2010). L’âge des peuplements correspond à l’âge du plus vieil arbre de chaque peuplement ou à l’âge des cicatrices de feux.
Dans les stations PGEN, ENPG1 et ENPG2 (figure 2), les taux de croissance en hauteur du pin gris et de l’épinette noire, calculés à l’aide de l’analyse de tige, ont été comparés à l’aide d’un test t (R Development Core Team 2011) pour tester l’hypothèse d’une croissance en hauteur plus lente de l’épinette noire par rapport à celle du pin gris. Pour comparer le taux de croissance des deux espèces au cours des années suivant leur établissement dans la station, le test t a été effectué à partir des taux de croissance en hauteur calculés entre la base de l’arbre et 1,3 m. Pour comparer le taux de croissance des espèces sur toute la durée de vie des individus, le test t a été effectué à partir des taux de croissance en hauteur calculés entre la base et l’apex de l’arbre.
15
Historique des feux
Les dates 14C étalonnées dans chaque station ont été regroupées sous la forme d’un graphique de probabilité cumulée à l’aide du logiciel Calib 6.1.0 (Stuiver et al. 2005). Ce graphique représente la somme de la distribution de probabilité de tous les charbons datés d’une station au cours du temps. Les probabilités sont standardisées pour que l’aire sous la courbe du graphique égale à 1. Ce graphique a l’avantage de représenter les dates 14C des charbons dans le temps ainsi que leur distribution de probabilité (Fesenmyer et Christensen 2010, de Lafontaine et Payette 2012).
À chaque station, les charbons provenant du même feu ont été regroupés en se basant sur l’intervalle de confiance (en âge calendaire) de leur date 14C. Lorsque les intervalles de confiance étalonnés de deux charbons se chevauchait, ces derniers ont été alors considérés comme provenant du même feu. Les intervalles de confiance variables associés à l’étalonnage des dates 14C doivent être pris en compte pour la reconstitution de l’historique des feux. Au cours de l’Holocène, le taux de 14C dans l’atmosphère a varié en fonction de l’activité solaire (Solanki et al. 2004, Reimer et al. 2009). Ces variations font en sorte que l’étalonnage des dates 14C forme parfois plusieurs intervalles de confiance répartis sur une longue période. Dans certains cas, l’étalonnage comprend plusieurs intervalles de confiance avec des probabilités semblables. Dans ces cas en particulier, il est difficile de déterminer quel intervalle doit être utilisé. Lorsque l’étalonnage d’une date 14C incluait deux distributions de probabilité relativement élevée, les deux distributions étaient utilisées pour créer deux scénarios d’historique des feux. Puisque les historiques des feux formés à partir des deux scénarios ne présentaient pas de différence significative, seul l’historique constitué à partir du scénario ayant la plus grande probabilité associée a été retenu.
La datation 14C correspond à l’âge du bois et non à celui du feu. Il est possible que des morceaux de bois mort au sol depuis un certain temps se transforment en charbon à l’occasion d’un feu, ce qui induit un biais dans la datation des feux, appelé erreur induite (Gavin 2001). Bien qu’elle ne puisse pas être négligée, plusieurs indices portent à croire que l’erreur induite est faible dans les forêts de la région d’étude. Dans la pessière à mousses, le taux de décomposition des restes ligneux au sol est rapide (Boulanger et Sirois 2006, Hagemann et al. 2009, Angers et al. 2012). Les arbres y ont une durée de vie dépassant rarement 250 ans et la fréquence des feux est relativement élevée (150-200 ans) (Bélisle et al. 2011), conditions qui ne favorisent pas l’accumulation de restes ligneux au sol. Par exemple, Payette et al. (2012) ont estimé que l’erreur induite est de moins de 25 ans dans les forêts de pin gris près de Sept-Îles. Pour estimer l’erreur induite, nous avons soustrait l’âge du peuplement daté à l’aide des rondelles des arbres à l’âge du feu recensé par les charbons le
16
plus récent dans chacun des sites. La différence d’âge entre les deux méthodes correspond à l’erreur induite de la datation.
Pour estimer l’activité des feux dans les stations au cours d’une longue période de temps, une analyse de la variation de l’abondance des charbons dans le temps, toutes stations confondues, a été effectuée. Les charbons plus jeunes que 5000 ans BP étal. ont été utilisés pour l’analyse en présumant une décroissance exponentielle de l'abondance des charbons avec le temps. Cette tendance proviendrait de la décomposition des charbons et du passage successif des feux brûlant à nouveau les charbons (Payette et al. 2012). Pour retirer cette tendance, une régression log-linéaire à l’aide de la fonction lm() du logiciel R (R Development Core Team 2011) a été effectuée sur la fréquence des dates de charbons de surface. Pour l’analyse, les dates 14C des charbons ont été regroupées par classe de 150 ans, étant donné que cette taille de classe maximisait le nombre de classes, tout en formant la distribution la plus uniforme possible. Par la suite, les résidus de la régression ont été utilisés pour exprimer la variation du nombre de charbons dans le temps toutes stations confondues.
L’échantillonnage aléatoire de charbons n’assure pas la détection de l’ensemble des feux. Afin de savoir si le nombre de feux détectés correspond au nombre de feux réels survenus dans chaque station, des courbes d’accumulation ont été tracées (de Lafontaine et Payette 2012). Ce type d’analyse a été utilisé pour évaluer l’effort d’échantillonnage nécessaire pour bien représenter une population donnée (Soberón et Llorente 1993). La courbe d’accumulation met en relation le nombre de feux détectés en fonction d’un nombre donné de charbons datés. Lorsque la courbe forme une asymptote, cela signifie que la datation éventuelle de nouveaux charbons ne permettrait pas de recenser de nouveaux feux, et que la plupart des feux de la station échantillonnée ont été effectivement détectés. La courbe d’accumulation a été calculée pour chaque station en utilisant la fonction accumresult() selon la méthode aléatoire de l’extension BiodiversityR du logiciel R (Kindt et Coe 2005, R Development Core Team 2011). Cette fonction trace 100 courbes d’accumulation en ré-échantillonnant aléatoirement les dates de charbon d’une station donnée. Ensuite, une courbe d’accumulation moyenne est tracée à partir de ces 100 courbes.
Un indicateur du nombre de feux réel a été calculé à partir des courbes d’accumulation moyenne de chaque station. Un indice a été calculé en effectuant une régression non linéaire du nombre de feux moyens détectés en fonction du nombre de charbons datés à l’aide de l'équation suivante :
17
𝐹(𝑛)= 𝐹𝑚𝑎𝑥(1 − 𝑒𝐾𝑛) [1]
où F(n) correspond au nombre de feux détectés, n au nombre de charbons datés, K à la constante contrôlant la forme de la courbe et Fmax à l’indice du nombre de feux. L’indice Fmax et la constante K ont été calculés avec la fonction nls() du logiciel R (R Development Core Team 2011). L’indice Fmax correspond au facteur contrôlant l’aplanissement de la courbe exponentielle négative considéré ici comme un estimateur du nombre réel de feux (Soberón et Llorente 1993, Payette et al. 2012, de Lafontaine et Payette 2012). L’indice Fmax a été ramené au nombre entier de manière à obtenir un nombre de feux entier.
L’intervalle de feu, défini comme le temps écoulé entre deux évènements de feu, a été calculé pour tous les feux détectés dans chaque station. L’intervalle de feu moyen (Im), c’est-à-dire la moyenne en années étalonnées de tous les intervalles de feu, a été calculé dans chaque station. Pour chaque station, l’histogramme du nombre d’intervalle en fonction de leur durée a été tracé pour représenter la distribution des intervalles. Les intervalles ont été regroupés en classe de 100 ans pour former une distribution la plus uniforme possible, tout en maximisant le nombre de classes. Afin d’apprécier la distribution des feux dans le temps, des distributions cumulatives ont été calculées avec la fonction Ecdf() du logiciel R (R Development Core Team 2011). Cette fonction calcule le pourcentage de feux recensé en fonction du temps.
Un intervalle moyen estimé (Ime) a été calculé [2] à partir de l’indice calculé par l’équation [1]:
𝐼
𝑚𝑒=
𝑛𝑃𝑒−1
[2]
où P correspond à la période de feux définie ici comme le temps écoulé entre le feu recensé le plus jeune et le feu recensé le plus vieux, ne au nombre de feux estimé (Fmax) à l’aide de la régression non linéaire de la courbe d’accumulation, où 1 a été soustrait étant donné que l’intervalle I débute et se termine par un feu. L’indice Ime est utilisé comme une valeur approximative de l’intervalle de feu moyen réel et pour évaluer la qualité du modèle en le comparant avec l’indice Im. Si la différence entre les deux indices est grande, cela signifie que plusieurs feux n’ont pas été identifiés et que la valeur de l’indice Im est incertaine.
18
Détection de feux régionaux
L’historique des feux des stations échantillonnées a été comparé afin d’estimer l’étendue des feux dans l’ensemble de la région étudiée. Pour ce faire, les dates de feu de toutes les stations qui présentaient moins de 25 ans de différence d’âge ont été regroupées. Un feu a été défini comme « régional » lorsqu’au moins 4 stations semblent avoir été affectées par un même feu. Pour chacun des feux régionaux recensés, un contour approximatif du feu a été tracé à l’aide des stations qui ont marqué le passage du feu.
19
Résultats
Caractéristiques des stations échantillonnées
Les stations comprenant des sols sableux et des sols sablo-loameux à pente faible sont toutes situées à moins de 500 m d’altitude. Les stations en pente se trouvent à plus haute altitude que les autres stations (respectivement 509 m et 550 m) (Tableau 2). Les sols de toutes les stations sont composés d’un sable loameux, sauf celui de la station PGS dont la texture est sableuse. L’épaisseur de la matière organique (horizons F et H) est inférieure à 10 cm dans la station PGS, tandis qu’elle varie entre 10 et 20 cm chez les autres stations. Le pH de l’horizon F de toutes les stations varie entre 3,6 et 4,2 (Tableau 3)
Tableau 2 Caractéristiques générales des stations échantillonnées.
Station Latitude N Longitude O Altitude (m) Pente (°)
PGS 50° 18' 11" 72° 32' 25" 435 3 PGEN 50° 17' 43" 72° 43' 15" 450 6 ENPG 1 50° 16' 59" 72° 37' 10" 465 5 ENPG 2 50° 11' 47" 72° 45' 56" 445 5 EN 1 50° 11' 50" 72° 36' 47" 455 5 EN 2 50° 17' 26" 72° 48' 51" 495 3 ENP 1 50° 22' 25" 72° 44' 28" 510 17 ENP 2 50° 26' 9" 72° 50' 19" 550 20
20
Tableau 3 Description générale des sols.
Station Texture du sol pH de l’horizon F (n=3/station) Épaisseur de la matière organique (n=25/station)
S S
PGS Sable 3,75 0,16 5 2,9
PGEN sable loameux 4,09 0,39 14,6 4,5
ENPG 1 sable loameux 3,68 0,21 13 4,6
ENPG 2 sable loameux 3,73 0,04 14,3 4,2
EN 1 sable loameux 4,19 0,08 23 6,4
EN 2 sable loameux 3,64 0,06 19,6 6
ENP 1 sable loameux 3,64 0,09 20,5 6
ENP 2 sable loameux 3,82 0,08 20,8 4,5
Les éricacées Kalmia angustifolia, Rhododendron groenlandicum, Vaccinium angustifolium et
Vaccinium myrtilloides Michx. dominent la strate arbustive dans toutes les stations. Les espèces Gaultheria hispidula (L.) Muhl. ex Bigelow, Cornus. canadensis L. et Clintonia borealis (Ait.) Raf. sont abondantes
uniquement dans les stations EN2, ENP1 et ENP2. La mousse Pleurozium schreberi abonde dans toutes les stations, sauf dans la station PGS où les lichens Cladonia rangiferina et Cladonia stellaris dominent la strate muscinale. Chez les stations PGEN et ENPG1, Pleurozium. schreberi est accompagné par des lichens, tandis que dans les stations ENPG2, EN1 et EN2, elle est accompagnée par Ptilium crista-castrensis et Sphagnum sp. Dans les stations ENP1 et ENP2, la strate muscinale est dominée par Ptilium crista-castrensis (Tableau
21 Tableau 4 Fréquence relative (%) des principales espèces d’éricacées, d’herbacées, de mousses et de
lichens des stations échantillonnées.
PGS PGEN ENPG1 ENPG 2 EN 1 EN 2 ENP 1 ENP 2
Ér ica cé es Rho gro 0 4 7 9 27 17 13 5 Kal ang 6 18 12 11 18 10 4 2 Vac ang 15 13 19 4 11 2 0 8 Vac myr 0 5 0 1 5 5 10 10 He rb ac ée s Gau his 0 0 0 10 1 13 18 14 Cor can 0 0 1 0 0 1 5 5 Cli bor 0 0 2 0 0 1 0 4 M ou ss es Ple sch 5 27 23 23 31 20 3 8 Pti cri 0 0 0 7 1 2 20 22 Sph sp. 0 0 1 1 5 9 7 1 Lic he ns Cla ran 7 10 7 1 6 0 0 0 Cla ste 30 15 9 0 2 0 0 0
Légende : Rho gro : Rhododendron groenlandicum, Kal ang : Kalmia angustifolia, Vac ang : Vaccinium
angustifolium, Vac myr : Vaccinium myrtilloides, Gau his : Gaultheria hispidula, Cor can : Cornus canadensis,
Cli bor : Clintonia borealis, Ple sch : Pleurozium schreberi, Pti cri : Ptilium crista-castrensis, Sph sp. :
22
Tableau 5 Nombre total de tiges, nombre de tiges >10 cm (DHP), surface terrière (S.T.) et surface terrière
relative (S.T. Rel.) des individus de la strate arborescente des stations échantillonnées.
PGS PGEN ENPG1 ENPG 2 EN 1 EN 2 ENP 1 ENP 2
n tota l Pic mar 149 1254 1225 1432 975 1146 917 494 Pin ban 67 50 11 40 0 0 0 0 Abi bal 0 0 0 0 0 25 0 464 Bet pap 0 0 0 0 4 1 4 1 n (d hp> 10 cm ) Pic mar 1 16 45 50 52 36 87 51 Pin ban 30 26 6 30 0 0 0 0 Abi bal 0 0 0 0 0 2 0 4 Bet pap 0 0 0 0 0 1 2 0 S. T. (m 2) Pic mar 0,09 0,99 1,61 1,56 1,61 1,41 2,64 1,61 Pin ban 0,69 0,59 0,13 0,57 0 0 0 0 Abi bal 0 0 0 0 0 0,05 0 0,27 Bet pap 0 0 0 0 0 0,02 0 0,01 S.T . R el. Pic mar 11 63 92 73 99 95 98 85 Pin ban 89 37 8 27 0 0 0 0 Abi bal 0 0 0 0 0 4 0 14 Bet pap 0 0 0 0 1 1 2 1
Légende : Pic mar : Picea mariana, Pin ban : Pinus banksiana, Abi bal : Abies balsamea, Bet pap : Betula
23
L’épinette noire domine quant au nombre d’individus dans toutes les stations (Tableau 5), étant donné le grand nombre de tiges issues de marcottage dans la classe 0-2 cm. La surface terrière de l’épinette noire est plus grande que celle du pin gris dans toutes les stations, à l’exception de la station PGS. Lorsqu’on prend en compte uniquement les tiges de plus de 10 cm de DHP, les tiges de pin gris sont plus nombreuses que les tiges d’épinette noire dans les stations PGS et PGEN, moins abondantes que les tiges d’épinette noire dans les stations ENPG1 et ENPG 2 et absentes dans toutes les autres stations (Figure 3).
0 200 400 600 800 1000 1200 1400 1600 1800 2000
PGS PGEN ENPG1 ENPG2 EN1 EN2 ENP1 ENP2
(n
tig
es
>1
0c
m)
/h
a
Stations
Picea mariana Pinus banksianaFigure 3 Nombre de tiges/ha de plus 10 cm au DHP de Pinus banksiana (en gris) et de Picea mariana (en
noir) dans chaque station.
Picea mariana Pinus banksiana
24
Figure 4 Structure de taille (DHP) des tiges vivantes et mortes du pin gris (en gris) et de l’épinette noire
(en noire) dans les stations PGS (a), ENPG2 (b) et EN2 (c). Les tiges sont regroupées en classes de 2 cm. Le nombre de tiges est additionné de 1 pour permettre la représentation logarithmique.
25
La structure de taille (DHP) des tiges de pin gris vivantes est caractérisée par une distribution en forme de cloche (Figure 4 a et 4 b). Peu de tiges de pin gris vivante sont présente dans les classes de diamètre inférieur à 4 cm. Les tiges de pin gris mortes sont distribuées uniformément dans les classes de moins de 12 cm. La structure de taille du pin gris est semblable dans toutes les stations où cette espèce est présente (Annexe 1). Les tiges d’épinette noire vivantes sont abondantes dans les classes de 0 à 4 cm (Figure 4 b). Le nombre d’individus est relativement stable dans les classes intermédiaires (4 à 10 cm). Le nombre d’individus décroît graduellement entre les classes de DHP de 10 à 20 cm. Ce type de distribution est semblable à celles des tiges d’épinette noire vivantes dans les stations ENPG1, ENPG2, EN2, ENP1, ENP2 (Annexe I). La station PGS présente quelques épinettes noires vivantes de plus de 10 cm seulement et une grande quantité de marcottes dans la classe 0-2 cm (Figure 4 a). La distribution des épinettes noires vivantes de la station EN2 est caractérisée par une décroissance du nombre d’individus dans les classes 2 à 12 cm, puis elle se présente sous forme de cloche entre les classes 12 à 22 cm (Figure 4 c). Les tiges d’épinette noire mortes de toutes les stations sont uniformément distribuées dans toutes les classes de moins de 25 cm.
Le taux de croissance en hauteur du pin gris calculé entre la base de l’arbre et 1,3 m au-dessus du sol est plus élevé que celui de l’épinette noire dans les stations PGEN et ENPG 2 (Tableau 6). Par contre, on n’observe aucune différence entre les deux espèces en ce qui concerne le taux de croissance en hauteur calculé entre la base et l’apex de l’arbre (Tableau 7). La croissance différentielle des deux espèces est observable sur les courbes de croissance des arbres des sites PGEN et ENPG2. Les courbes du pin gris sont au-dessus de celles de l’épinette noire en début de croissance, tandis qu’on n’observe pas de différence entre les deux groupes en fin de croissance (Figure 5). Dans la station ENPG1, on n’observe pas de différence significative du taux de croissance en hauteur des deux espèces calculé entre la base de l’arbre et 1,3 m au-dessus du sol ni entre le taux de croissance en hauteur calculé entre la base et l’apex des arbres.
26
Tableau 6 Taux de croissance en hauteur (cm/année) entre la base des arbres et 1,3 m au-dessus du sol
chez les pin gris et les épinettes noires des stations PGEN, ENPG1 et ENPG2. Un test de t a été effectué afin de vérifier si une différence significative existe entre le taux de croissance en hauteur des deux espèces.
Site n Pinus n Picea
Taux de croissance entre 0 et 1,3 m du sol Test t
Pinus Picea t p S S PGEN 5 5 6 0 13 6 -2,82 0,0473 ENPG1 5 4 11 1 9 2 1,82 0,1118 ENPG2 5 5 9 2 21 4 -5,99 0,0006
Tableau 7 Taux de croissance en hauteur (cm/année) des tiges de pin gris et d'épinette noire des stations
PGEN, ENPG1, ENPG2. Un test de t a été effectué afin de vérifier si une différence significative existe entre le taux de croissance des deux espèces.
n Pinus n Picea
Taux de croissance totale Test t
Pinus Picea t p S S PGEN 5 5 12 1 13 2 -0,74 0,4867 ENPG1 5 4 15 1 14 1 1,02 0,3486 ENPG2 5 5 16 1 16 2 1,96 0,5309
27 Figure 5 Hauteur des tiges de pin gris (ligne hachurée) et d’épinette noire (ligne continue) en fonction du
28
Datation des peuplements
Dans toutes les stations, l’année d’établissement minimum des arbres s’étend sur une période de 5 à 30 ans (Figure 6). Dans les stations PGS, PGEN et ENPG2, l’installation du pin gris s’est faite plus tôt que celle de l’épinette noire, tandis qu’aucune différence significative n’a été constatée dans le site ENPG1 (Tableau 8), une différence variant seulement entre 3 et 7 ans. Dans la station PGS, l’épinette noire se serait établie une trentaine d’années après le pin gris.
Tableau 8 Année d’établissement du pin gris et de l'épinette noire dans les stations PGS, PGEN, ENPG1,
ENPG2.
Station n Pinus n Picea
Année installation Test t
Picea Pinus t p 𝑥̅ S 𝑥̅ S PGS 10 3 1944 1 1917 1 48,33 <0,0001 PGEN 5 5 1930 3 1923 1 4,54 0,0019 ENPG1 5 4 1926 3 1923 2 1,73 0,1275 ENPG2 5 5 1927 2 1923 1 4,54 0,0019
Les plus vieux arbres des stations PGS, PGEN, ENPG1, ENPG 2 et EN1 se sont établis entre 1915 et 1926 (Figure 6, Tableau 9). Chez les stations PGS, ENPG1, ENPG2 et EN1, la date d’établissement du peuplement a pu être déterminée précisément grâce à la datation de cicatrices formées en 1915 et en 1917 (Tableau 9). Des cicatrices de feux datant de 1790 et de 1855 ont aussi été échantillonnées à proximité de la station PGS. Chez les stations PGS, ENPG1, ENPG2 et EN1, l’établissement des arbres portant des cicatrices se situe entre 1836 et 1864. Chez la station ENP1, la date d’établissement minimale du plus vieil
29
arbre est 1872. Dans les stations EN2 et ENP2, les dates d’établissement des tiges s’étalent entre 1833 et 1868.
Tableau 9 Datation des feux les plus récents survenus dans chaque station.
Station Dernier feu
PGS 19151, 18521,3, 17901 PGEN 19222 ENPG1 19171,18533 ENPG2 19171,18363 EN1 19171,18643 EN2 18332 ENP1 18722 ENP2 18402
On n’a pas noté de différence d’âge marquée entre les peuplements classés comme étant en début de succession (PGEN), en milieu de succession (ENPG1 et ENPG2) et en fin de succession (EN1) (Figure 6). Seul le peuplement de la station EN2 s’est établi avant ceux des autres stations servant à tester l’hypothèse de la chronoséquence.
1 Daté à l’aide des cicatrices de feux
2 Daté à l’aide de la date d’établissement des arbres
30
Figure 6 Fréquence des tiges de Picea mariana (en noir), de Pinus banksiana (en gris) et d’Abies balsamea
31
Un total de 73, 91 et 118 macrorestes ligneux ont été dénombrés dans les stations ENPG1, ENPG 2 et PGEN (Tableau 10). Étant donné que la décomposition des restes ligneux était avancée dans toutes les stations, il n’a pas été possible de localiser la souche des macrorestes ligneux chez la moitié des cas étudiés. Chez les macrorestes dont la souche n’a pu être localisée, il n’a pas été possible de confirmer leur appartenance à la parcelle d’échantillonnage. Dans le site ENPG1, 63 % des restes ligneux étaient des bois d’épinette, 23 % des bois de pin gris et 4 % des bois non identifiés. Dans la station ENPG2, 42 % des restes ligneux étaient des bois d’épinette et 68 % des bois de pin gris. Dans la station PGEN, 50 % des restes ligneux étaient des bois d’épinette, 43 % des bois de pin gris et 7 % des bois non identifiés.
Tableau 10 Nombre de macrorestes ligneux d’épinette et de pin dans les stations ENPG1, ENPG2 et PGEN.
Le chiffre entre parenthèses correspond au nombre de restes ligneux sans souche.
Stations
ENPG1 ENPG2 PGEN
Picea 31 (15) 30 (9) 42 (17)
Pinus 17 (7) 31 (21) 29 (22)
32
Composition botanique des charbons
Un total de 813 pièces de charbon de bois ont été récoltées à la base de l’horizon organique de surface, soit une moyenne de 102 pièces de charbon par station (Tableau 11). Par ailleurs, 43 % des charbons ont un poids supérieur à 5 mg et 65 % de ceux-ci ont été identifiés au genre ou à l’espèce. Dans la station PGS, seuls des charbons et des écailles de cônes carbonisées de pin gris ont été identifiés. Les charbons de la station PGEN et ENPG2 sont composés majoritairement d’épinette. Les pièces de charbon de la station ENPG1 comprennent uniquement du bois d’épinette. La station EN1 renferme un assemblage de charbons d’épinette et de mélèze laricin, alors que les stations EN2, ENP1, ENP2 se caractérisent par un assemblage dominé par l’épinette, le sapin baumier et le mélèze laricin.
Un total de 5418 pièces de charbon de bois ont été dénombrées dans le sol minéral, soit une moyenne de 677 pièces de charbon par station (Tableau 12). De ce total, 30 % des charbons ont un poids supérieur à 5 mg et 52 % de ceux-ci ont été identifiés au genre ou à l’espèce. Les pièces de charbon enfouies dans le sol minéral de la station PGS proviennent de bois de pin gris comme ceux du compartiment de surface. Les stations PGEN, ENPG1 et ENPG2 présentent un assemblage dominé par l’épinette accompagné par le pin gris et le bouleau. Les stations EN1, EN2, ENP1 et ENP2 renferment un assemblage dominé par l’épinette mais accompagné par plusieurs pièces de charbon de sapin baumier, en particulier dans la station ENP2.
33 Tableau 11 Composition botanique des charbons de surface des stations échantillonnées.
Stations
PGS PGEN ENPG1 ENPG2 EN1 EN2 ENP1 ENP2
Ch ar bon s d e b ois Picea sp. 0 20 24 22 29 23 17 21 Pin ban 18 8 0 8 0 0 0 0 Lar lar 0 0 0 2 2 0 3 0 Abi bal 0 0 0 0 0 2 1 6 feuillu ind. 1 8 8 12 7 5 0 2 conifère ind. 5 4 5 3 7 3 1 7 Ind. 4 0 3 3 2 7 3 6 Éca ille s d e cô ne s car bo nis ée s Picea sp. 0 0 5 2 8 0 0 0 Pin ban 6 0 0 1 0 0 0 0 Ind. 9 0 7 0 2 0 0 0 n N >5mg 43 40 52 53 57 40 25 42 n 57 121 125 120 131 84 75 100
Légende : Picea sp. : Picea sp., Pin ban : Pinus banksiana, Abi bal : Abies balsamea, Lar lar : Larix laricina feuillu
34
Tableau 12 Composition botanique des charbons du sol minéral des stations échantillonnées.
Stations
PGS PGEN ENPG1 ENPG2 EN1 EN2 ENP1 ENP2
Ch ar bo ns de b ois Picea sp. 0 65 31 34 9 300 31 17 Pin ban 66 1 1 4 0 0 0 0 Lar lar 0 1 0 0 0 3 0 1 Abi bal 0 0 4 0 4 16 11 31 Bet sp. 1 16 6 0 2 17 10 3 feuillu ind. 16 3 4 7 0 16 6 2 conifère ind. 29 33 34 12 11 110 17 39 ind. 67 35 55 13 18 78 33 70 Éca ille s d e cô ne s car bo nis ée s Picea sp. 0 6 9 0 2 37 26 78 Pin ban 17 0 0 1 0 0 0 0 ind. 28 6 1 1 9 18 6 10 n n>5 mg 224 166 145 72 55 595 140 251 n 1121 497 411 282 245 1286 521 1055
Légende : Picea sp. : Picea sp., Pin ban : Pinus banksiana, Abi bal : Abies balsamea, Lar lar : Larix laricina feuillu
35
Abondance et composition botanique des charbons de bois au
cours du temps
La date 14C et l’identification botanique des charbons de bois, ainsi que les intervalles de feu étalonnés ayant la plus haute probabilité sont présentés en Annexe II. Le nombre de charbons, toutes stations et tous compartiments de surface et de sol regroupés, diminue continuellement avec le temps (Figure 7). Pas moins de 97 % des charbons de bois sont plus jeunes que 5000 ans BP étal.; seulement 9 charbons de bois ont un âge variant entre 5000 et 8000 ans BP étal. Le nombre de charbons de bois du compartiment de surface diminue avec le temps (Figure 7). Plus de 95 % des charbons de bois de ce compartiment ont moins de 2000 ans; le plus vieux charbon de bois dans ce compartiment est un conifère indéterminé de la station ENP1 datant de 3520 ans BP. étal. Le nombre de charbons de bois dans le sol minéral ne diminue pas au cours du temps comme dans le cas du compartiment de surface (Figure 7). Le plus vieux charbon (conifère indéterminé) de ce compartiment provient de la station EN1et date de 7540 ans BP étal.
36 Fi gur e 7 No m br e t ot al de ch ar bo ns d e b ois d at és d an s t ou tes le s s ta tion s e n f on cti on d u t em ps (c las se de 1 50 an s) . ( To tal ), e n s ur fa ce (S ur fa ce ) e t e nf ou is da ns le so l m iné ra l ( Mi né ra l).