Activation des cellules dendritiques par des composantes de bioaérosols
Texte intégral
Figure
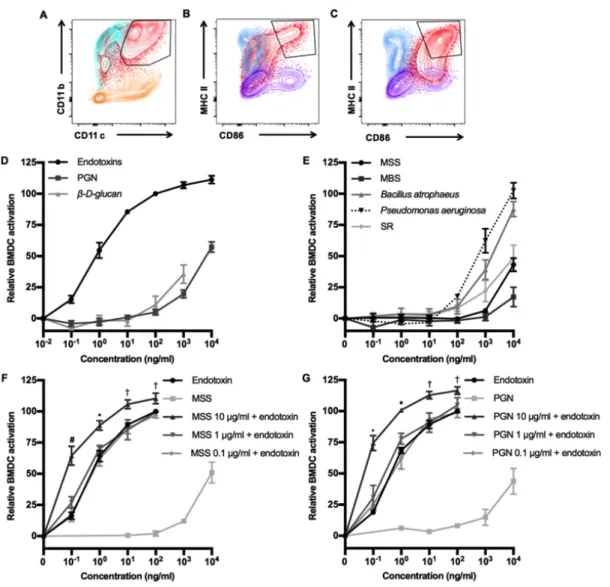
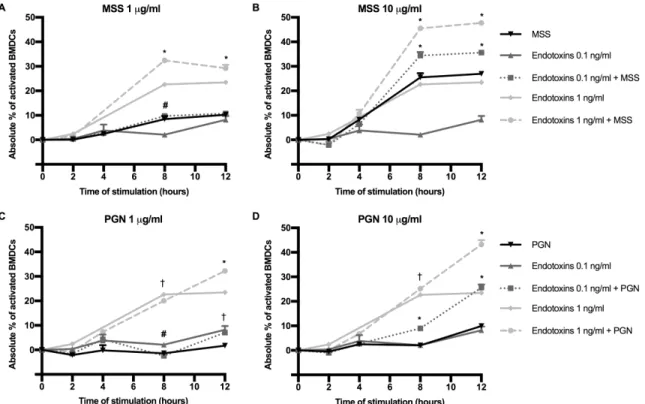
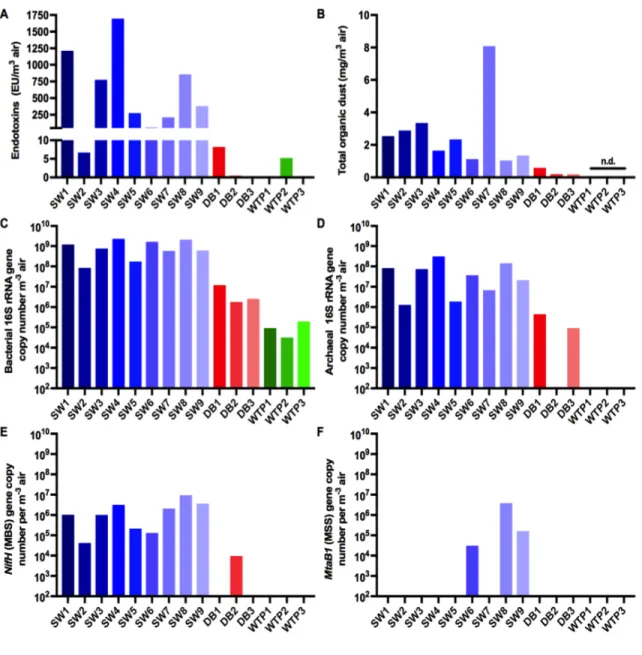


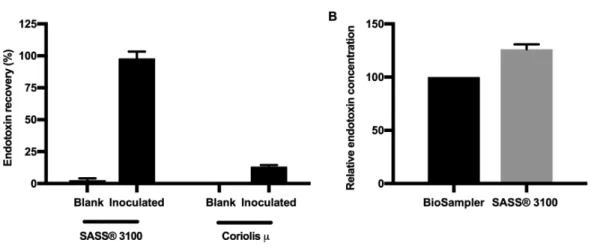

Documents relatifs
The CDI6(+) (fcgammaRlll(+)) subset of human monocytes preferentially becomes migratory dendritic cells in a model tissue setting.. Distinct functional role of Hepatitis C virus
Figure 1. Morphologie des pDC.. Ontogénie des cellules dendritiques ... Localisation et migration des pDC dans l’organisme. Activation des pDC ... Polarisation de l’immunité par
L’œdème de l’oreille induit par l’huile de croton chez la souris qui est aussi un modèle de l’inflammation aiguë, a été utilisé pour évaluer
Pour élaborer l’expression de la section efficace triplement différentielle SETD dans le référentiel du laboratoire, on applique le modèle du cœur gelé à un
Nous avons commencé par évaluer le potentiel chimique des deux types de coproduits pour la production de bioéthanol en terme de teneur en glucose avant d’examiner l’impact
Élaborer un protocole expérimental permettant de faire disparaître du cuivre métal puis de le faire réapparaître après une succession de réactions utilisant
L’archive ouverte pluridisciplinaire HAL, est destinée au dépôt et à la diffusion de documents scientifiques de niveau recherche, publiés ou non, émanant des
Ces modèles de sensibilisation permettent d’ évaluer le potentiel allergénique de protéines alimentaires, de disposer de tests permettant de contrôler les
