HAL Id: hal-02575604
https://hal.inrae.fr/hal-02575604
Submitted on 14 May 2020HAL is a multi-disciplinary open access archive for the deposit and dissemination of sci-entific research documents, whether they are pub-lished or not. The documents may come from teaching and research institutions in France or abroad, or from public or private research centers.
L’archive ouverte pluridisciplinaire HAL, est destinée au dépôt et à la diffusion de documents scientifiques de niveau recherche, publiés ou non, émanant des établissements d’enseignement et de recherche français ou étrangers, des laboratoires publics ou privés.
Étude ichtyologique des lacs d’altitude de la Corse,
fasc.1 : I- le lac de Bastani
B. Dumont, B. Rivier
To cite this version:
B. Dumont, B. Rivier. Étude ichtyologique des lacs d’altitude de la Corse, fasc.1 : I- le lac de Bastani. irstea. 1987, pp.50. �hal-02575604�
87/0019
CEMAGREF
CENTRE NATIONAL DU MACHINISME AGRICOLE DU GENIE RURAL DES EAUX ET DES FORETS GROUPEMENT D AIX-EN-PROVENCE
LE THOLONET
CEMAGREF PARC NATUREL REGIONAL
DE LA CORSE
ETUDE ICHTYOLOGIQUE DES LACS
D'ALTITUDE DE LA CORSE
I) LE LAC DE BAST ANI
Auteurs :
B. RIVIER Gt B. DDMONT
Ingénieurs Hydrobiologistes
Division Q.E.P.P FEVRIER 1987
Groupement d'Aix-en-Provence
S O M M A I R E
Remerciements
Introduction 1
Moyens et méthodes d'étude 2
1 Description des filets. Avantages et inconvénients dus à
leur utilisation 3
2 Manipulations effectuées sur les poissons. Biométrie.
Marquages 7
II Déroulement de la campagne de terrain 8
III Etude ichtyologique du lac de BASTIANI 10
1 Rappel des principales caractéristiques morpho-dynamiques.,
physico-chimiques et biologiques du lac 10
2 Origine des peuplements piscicoles. Observations
ichtyolo-giques antérieures à l'inventaire 13
IV Résultats 15
IV
Effort de pêche mis en oeuvre 15
2 Définition de l'unité d'effort de pêche 15
3 Bilan global des inventaires 15
4 Taux de survie et résultats des marquages 18
I V
5 Bilan global pondéré 21
6 Analyse des résultats par dimension de maille... 24
6-1 Efficacité comparée des différentes mailles 24
Principales caractéristiques de la population
de saumons de fontaine 30
1 Répartition des effectifs par classes de taille 30
2 Relations longueur/masse. Coefficients de condition 33 3 Sex-ratio et comparaison de la condition des mâles et
femelles 41
4 Cycle sexuel 41
4-1 Cycle ovarien 42
4-2 Cycle spermatogénétique 42
-3 Fécondité et état de maturité sexuelle 43
VI Orientations des travaux futurs 46
1 Eléments de biologie 46
2 Impact halieutique 47
Conclusion 48
REMERCIEMENTS
Plus que toute autre étude, les inventaires ichtyologiques en lacs d'altitude nécessitent la mise en oeuvre de moyens humains et matériels importants et le déroulement d'une campagne de terrain ne constitue souvent que l'aboutissement d'une longue démarche au cours de laquelle la bonne volonté de chacun est indispensable pour aboutir à la couverture financière et technique de l'opération.
L'étude du lac de Bastani n'a pu être envisagée que dans cette optique et il est particulièrement agréable d'adresser nos plus sincères remerciements à M. le Directeur du Parc Naturel Régional de Corse pour avoir autorisé le déroulement des inventaires et pour la part prise dans le financement de l'opération.
Nos remerciements s'adressent aussi à M. GAUTHIER, Conseiller Scientifique auprès du Parc Naturel qui en assurant le rôle ingrat de coordinateur entre les différents services a permis à l'opération de voir le jour.
Nous tenons également à exprimer notre profonde gratitude à l'égard de la Fédération des AAPP de la CORSE avec à sa tête M. le Président MARTINI qui en finançant l'opération d'héliportage a été à la base de toute l'opération.
Nous remercions aussi la Fédération des AAPP pour l'accueil chaleureux qu'elle nous a réservé sur l'île et pour le concours qu'elle nous a apporté par la participation aux opérations de terrain des gardes pêche MATTEI et PHILIPPI qui malgré des conditions de travail difficiles ont fait preuve d'un dévouement et d'un esprit d'initiative remarquables.
L'appui technique apporté par M. Bernard ROCHE du SRAE de CORSE sous la forme de communication de nombreux documents et de par sa participation aux opérations de terrain a également constitué un apport précieux à l'étude et nous l'en remercions vivement.
INTRODUCTION
Les études ichtyologiques des lacs d'altitude de Corse, entreprises en 1986 à la demande du Parc Naturel Régional, font suite aux travaux de Gauthier et al ( 1984 ) consacrés à la bathy-métrie, la physico-chimie, la flore aquatique et aux invertébrés benthiques.
Elles complètent donc, dans le cas des lacs renfermant des peuplements piscicoles, les diagnoses précédentes en permettant une approche synthétique des biocénoses de ces milieux.
Considérés sous l'aspect de la biocénotique puis de l'écologie générale des lacs, les poissons présentent un intérêt fondamental dans la mesure où leur position apicale au sein du réseau trophique et leur cycle de vie pluri-annuel leur confère un degré élevé d'intégration des conditions de milieu et en fait d'excellents révélateurs de l'évolution passée et à venir des biocénoses.
L'étude des ichtyocénoses au cours d'inventaires dont le but est la mise en évidence de la nature et de la structure des peu-plements, constitue par conséquent un élément indispensable en matière de gestion piscicole. En effet, les poissons constituent la seule fraction biocénotique directement exploitée par l'homme, sous forme dans le cas présent de pêche récréative, ce qui leur confère une valeur socio-économique indéniable. La nécessité de concilier les activités halieutiques et le maintien de l'équilibre qualitatif et quantitatif des peuplements ( que la pêche tend dans une certaine mesure à détruire, mais sans lequel elle ne peut exister ) conduit à définir des règles de gestion qui pour être rationnelles doivent inclure une connaissance aussi précise que possible des peuplements piscicoles.
I - MOYENS ET METHODES D'ETUDE
Lors d'études ichtyologiques en lacs de montagne, la
méthode d'échantillonage à mettre en oeuvre doit permettre de concilier deux impératifs principaux:
- être adaptée aux espèces susceptibles d'être rencontrées et à la dimension du milieu ( les lacs même d'importance réduite représen-tent un volume d'eau important ) de manière à être suffisamment effi-cace pour permettre l'obtention d'un échantillon donnant une image aussi proche que possible du peuplement réel.
- être suffisamment douce pour éviter au maximum les mortali-tés de poissons à haute valeur halieutique et ainsi autoriser l'étude ultérieure de certains paramètres individuels : croissance, longévité, maturité sexuelle ; ou démographiques : mortalités par pêche, recru-tement et évolution des stocks.
Alors que les engins actifs permettent la réalisation de la deuxième condition, leur usage s'avère impossible en lacs de montagne en raison de leur manque d'efficacité ou de l'impossibilité de les mettre en oeuvre : le volume d'eau à prospecter constitue un obstacle à l'utilisation de la pêche électrique dont l'effet attractif décroit rapidement avec la profondeur.
: la nature souvent très irrégulière des fonds empêche l'usage des filets tractés tels que sennes ou verveux.
: les lacs sont souvent trop petits et à fond trop irrégulier pour permettre l'usage des chaluts qui nécessitent par ailleurs pour être mis en oeuvre des embarcations et des moteurs im-possibles à transporter en altitude.
Des expérimentations antérieures effectuées avec des engins passifs susceptibles de capturer les poissons vivants : nasses ou trappes de capture ont mis en évidence l'inefficacité de ces
modes de capture pour les salmonidés qui constituent dans la plupart des lacs de montagne l'essentiel du peuplement.
Les modes de pêche précédents ne permettent donc pas de dresser un inventaire fiable des lacs de montagne en raison notamment de l'impossibilité de prospecter tous les habitats.
Parmi les autres moyens d'investigation, les contrôles des prises effectuées par les pêcheurs amateurs, bien que susceptibles d'apporter des informations sur la pression de pêche et sur la morta-lité piscicole qui en résulte, sont souvent trop fragmentaires pour être utilisés dans le cadre d'un inventaire précis des peuplements. L'impossibilité matérielle de contacter l'ensemble des pêcheurs si une structure fiable de recueil des données n'est pas mise en place ou si une présence permanente sur le terrain d'agents chargés du contrôle n'est pas assurée, conduit à une perte importante d'information. De plus, la pêche aux lignes est souvent très sélective car orientée vers certaines espèces et pour chacune d'elles vers certains stades de développement desquels sont exclus les juvéniles, de même qu'elle ne concerne qu'une partie des habitats surtout localisés dans les zones de bordure.
L'inadéquation des modes précédents d'échantillonnage a donc conduit à réaliser l'inventaire des lacs de montagne à l'aide de filets maillants couramment utilisés en limnologie ( LE CREN et al, LEOPOLD et al... in EIFAC 1975, BARBIER 1985 ) et qui sont employés traditionnellement par les pêcheurs professionnels.
I1 - DESCRIPTION DES FILETS. AVANTAGES ET INCONVENIENTS DUS A LEUR
UTILISATION.
Les filets maillants sont des engins de capture statiques dans lesquels la plupart des espèces se prennent par passage dos opercules à l'intérieur d'une maille unique pour
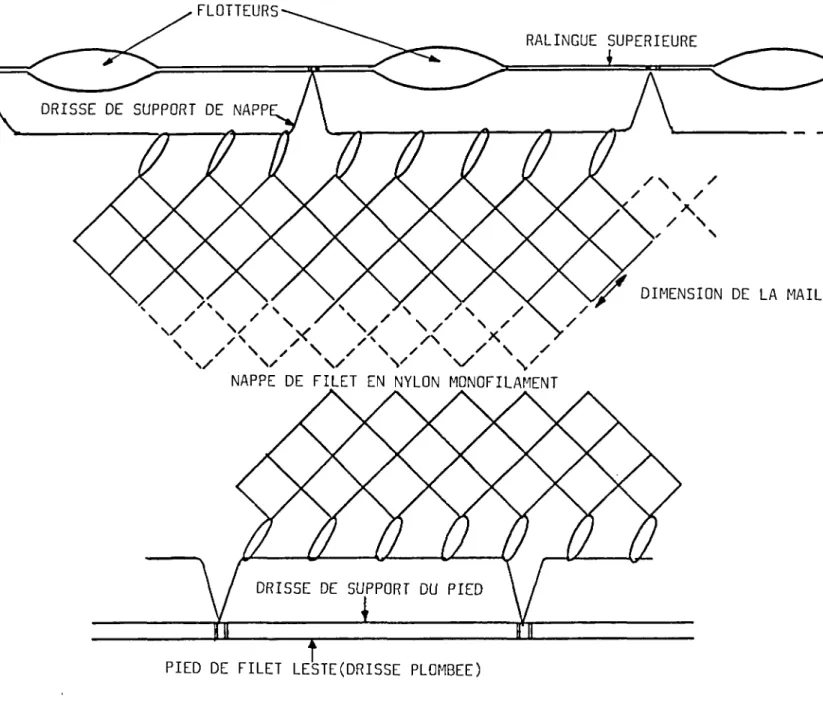
le plus souvent s'emmêler ensuite dans la nappe ( BRANDT 1975 ) . Ils sont constitués d'une nappe unique de filet en nylon monofilament, montée entre une ralingue supérieure munie de flotteurs et un pied de filet plombé ( Fig 1 ) . Sur les lacs de CGRSE, deux types de filets ont été utilisés :
1 ) Les araignées qui sont des filets reposant sur le fond grâce à un pied fortement lesté. Le filet est maintenu vertical par les petits flotteurs de la ralingue et échantillonne donc depuis le fond une tranche d'eau égale à sa hauteur.
2 ) Les filets de surface à l'inverse des précédents sont peu lestés et se maintiennent en surface grâce à leurs flotteurs. Ils opèrent donc dans les couches superficielles depuis la surface jusqu'à une profondeur égale à la hauteur de la nappe.
Les avantages des filets maillants sont liés à :
- leur faible encombrement et leur faible poids qui les rendent facilement transportables en altitude.
Leur facilité d'utilisation :
- la pose et la levée des filets ne nécessitent qu'un effort de mise en oeuvre réduit et peuvent être facilement effectuées à la rame depuis une embarcation pneumatique.
- la possibilité qu'ils offrent d'être utilisés dans tous les habitats y compris les zones les plus profondes et les plus accidentées.
L'inconvénient majeur des filets maillants est de provoquer la mor-talité quasi totale des poissons capturés ( qui maillés et emmêlés s'asphyxient ou s'épuisent en tentant de se libérer des mailles ) lorsqu'ils sont utilisés classiquement c'est-à-dire pour des temps de pose s'étalant sur plusieurs heures.
Les expériences précédemment effectuées par le CEMAGREF sur les grands plans d'eau ont cependant montré que les salmonidés présen-taient la particularité de se prendre non par les opercules, mais
FLOTTEURS
RALINGUE SUPERIEURE
DRISSE DE SUPPORT DE NAPP
DIMENSION DE LA MAILLE
\
\/
/
\ / \ /
\ / \y \/ V \/ \ ^
NAPPE DE FILET EN NYLON MONOFILAMENT
DRISSE DE SUPPORT DU PIED
PIED DE FILET LESTE(DRISSE PLOMBEE)
la plupart du temps par les dents sans se mailler au sens strict. Ces observations ont donc abouti à la conclusion que l'usage des filets maillants était possible en lacs de montagne à condition de ne pêcher que pendant des durées très courtes afin d'éviter l'emmêlage et ainsi obtenir des taux de survie compatibles avec la condition de protection des peuplements.
L'autre désavantage des filets maillants se situe dans leur sélec-tivité à l'égard d'une classe de taille donnée et dans l'existence d'artefacts dus au fait que des poissons de taille relativement importante par rapport à la maille du filet sont susceptibles de se prendre par simple emmêlage sans se mailler.
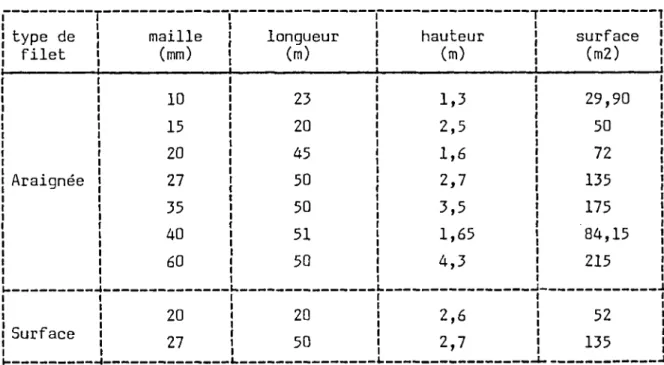
Afin d'éviter ces biais une gamme de filets de mailles s'étendant de 10 à 45 mm a été utilisée. Les caractéristiques des engins sont rassemblées dans le tableau 1.
1 1 type de ¡ filet i Araignée i i i i 1 i Surface maille (mm) 10 15 20 27 35 40 60 | 20
i
27
i longueur (m) 23 20 45 50 50 51 50 C M L A 1 l 1 n hauteur (m) 1,3 2,5 1,6 2,7 3,5 1,65 4,3 2,6 2,7 surface ¡ (m2) ¡ 1 129,90 ¡
50 ¡
72 ¡
135 j
175 ¡
84,15 1
215 ¡
¡ i52 ¡
135 ¡
i iTABLEAU I : Types et dimensions des filets utilisés sur les lacs de CORSE au cours de la campagne de 1986.
I2 ) MANIPULATIONS EFFECTUEES SUR LES POISSONS. BIOMETRIE. MARQUAGES.
Lors de la relève, les poissons capturés sont retirés des mailles sur le lieu de pêche s'ils sont simplement pris par les dents et conservés dans des récipients distincts pour chaque filet ou bien démaillés et désemmêlés sur le bord après avoir été transportés avec les engins dans des bacs remplis d'eau puis replacés avec les sujets pris dans le même filet.
Après tranquillisation par immersion dans un bain constitué d'une solution de phénoxy-2-éthanol à une concentration de 0,5 ml par litre d'eau, les mesures des paramètres biométriques portant sur les longueurs et les poids sont effectuées.
Les longueurs prises en compte sont :
- la longueur à la fourche (LF) comprise entre l'extrémité du museau et l'échancrure de la nageoire caudale.
- la longueur totale (LT) comprise entre l'extrémité du museau et la partie postérieure des lobes de la nageoire caudale placés en position naturelle.
Les valeurs des longueurs sont appréciées au millimètres près.
Les poids ( P ) sont lus avec une précision de l'ordre du gramme à l'aide de pesons, de différentes gammes de poids suivant la masse des poissons. Les sujets n'ayant pas survécu aux opérations de pêche font ensuite l'objet d'observations relatives au sexe et à l'état de
maturité. Les tubes digestifs et les têtes sont conservés dans une
solution formolée afin d'étudier les régimes alimentaires et d'effectuer les déterminations d'âge par otolimétrie.
Les poissons survivants sont identifiés par apposition de marques mâchoires (marques boucles de type PRESADOM ) portant un numéro individuel et après récupération retournés au lac.
II ) DEROULEMENT DE LA CAMPAGNE DE TERRAIN
La campagne d'inventaire effectuée du 21 au 26 juillet 1986 devait être consacrée à l'étude des lacs de BASTANI et de
ROTONDO ( Fig 2 ) situés en Corse du Nord et appartenant respectivement aux bassins versants du FIUM'ORBU et de la RESTQNICA puis du TAVIGNANO.
Alors que les pêches ont pu se dérouler normalement pour le lac BASTANI, les conditions météorologiques existant sur la CORSE dans la deuxième partie delà semaine : vents d'Ouest soufflant en rafales avec des vitesses de l'ordre de 80 km/h n'ont pas permis de travailler de manière satisfaisante à ROTONDO. L'accès au lac rendu extrêmement périlleux par les vagues et les rafales de vent n'a permis la pose que de quelques filets dans la zone littorale et a occasionné des dégâts importants au matériel. Seules trois truites fario ( TRF ) et un saumon de fontaine ( SDF ) dont les longueurs, poids et numéros de marque sont indiqués dans le tableau 2 ont été capturées.
Ces résultats beaucoup trop fragmentaires pour aboutir à des conclusions montrent seulement que le lac de ROTANDO renferme des sujets de taille importante dont la présence semble liée à une longévité élevée.
Dans l'hypothèse où les inventaires seraient reconduits en 1987, il serait donc souhaitable de réinscrire le lac de ROTONDO au programme des opérations.
i 1 r , ESPECE
TRF
¡ TRF
¡ TRF
¡ SDF
. LF (mm) 348 415 438 360. LT (mm)
360 430 450 375 P ( g ) 440 740 900 540 . n° de marque ¡ EL 6715 ¡ EL 6717 ¡ EL 6749 ¡ EL 6716 ¡TABLEAU 2 : Caractéristiques biometriques et n° des marques des poissons capturés sur le lac de ROTONDO.
BASTIA
LAC DE BASTAN^ / FIUM
BONIFACIO
IQ
III ) ETUDE ICHTYOLOGIQUE DU LAC DE BASTANI
1 1 1 1 RAPPEL DES PRINCIPALES CARACTERISTIQUES H0RPH0-DYNAMIQUE5 PHYSICO-CHIMIQUES ET BIOLOGIQUES DU LAC
La description détaillée du lac de BASTANI a été effectuée par GAUTHIER et ROCHE ; ( in GAUTHIER et al. 1984 ) , aussi ne seront rappelés ici que les éléments pouvant être utiles à l'interprétation des données ichtyologiques.
Formé durant le tardi-glaciaire, environ 14000 ans avant notre ère, soit à une période géologique récente, le lac de BASTANI est un lac de cirque fermé par une moraine et est situé sur un substrat constitué de granodiorites et de granites monzonitiques. L'érosion de ces formations relativement altérables occasionne des apports sa-bleux importants qui transportés par les eaux de fonte de névé oc-casionnent la formation d'un delta sous lacustre dans la portion Ouest du lac ( GAUTHIER 1984 ) . Les rives sont formées de gros blocs morainiques au Nord et à l'Est délimitées par une falaise à l'Ouest et formées par le fond du cirque au Sud-Ouest. Les principales caractéristiques du lac sont résumées dans le tableau 3.
L'altitude supérieure à 2000 m explique la période de gel qui dure en moyenne 7 à 8 mois et la position du lac dans l'étage de végétation subalpin à aulne odorant ( Alnus sualveoleus ) .
D'une profondeur moyenne de 10,1 m pour une profondeur maximale de 24 m ( ROCHE 1984 ) le lac de BASTANI est relativement profond. La carte bathymétrique ( fig 3 ) met en évidence une dissy-métrie transversale des fonds : la rive Est est la plus abrupte,
les pentes étant plus douces dans la portion Nord où la profondeur est la plus faible et au Sud au pied des éboulis.
40mm 22/07(3
20 mm 22/07(3)
35mm 22/07(3
10mm 22/07(3)
40 mm 22/07(1)
20mm 22/07(1
40mm 22/07(2)
20mm 22/07(2)
27mm 22/07
10mm 22/07(2)
10mm 22/07(7)
35mm 22/07(1) /
27mm 22/07(2)
0
50
100m
FIG 3;BATHYMETRIE DU LAC DE BASTANI,(d'après ROCHE 1984) ET EMPLACEMENT DE POSE
DES FILETS
X-
-*:ARAIGNEE ••
•+ : SURFACE
Les chiffres entre parantheses font reference aux différentes poses du 22/07/86
12
Altitude ( m )
Superficie du bassin versant ( HA )
Superficie du lac ( HA )
Rapport superficie lac/Superficie BU ( °a )
Profondeur moyenne ( m )
Profondeur maximale ( m )
Périmètre ( m )
Volume ( m3 )
Taux de renouvellement théorique ( mois )
2089 32,4 4,38 13 10,1 24 925 450 000 15 à 18
Tableau 3 : Principales caractéristiques physiques du lac de BASTIANI (d'après ROCHE 1984).
13
Les profils verticaux de température estivale n'indiquent pas de stratification nette vraisemblablement en raison de la pré-sence fréquente de vents violents favorisant le brassage des eaux
( ROCHE 1984 ).Les teneurs en oxygène dissous sont au cours de la même période, proches de la saturation sur toute la tranche d'eau et parfois très supérieures à cette valeur en raison d'une activité phytoplanctonique intense.
Les eaux peu minéralisées ( conductivité voisine de 25 VS ) sont caractérisées par un pH de 8 à 9,3 et par des teneurs en nitrates faibles liées à l'importance des peuplements phytoplanctoniques et à la mobilisation de ces ions par les organismes.
La valeur de la productivité primaire qui atteint
300 mg C/m2-j et de la biomasse phytoplanctonique ( 45 mg de chloro-phylle a /m3 ) qui a pour corollaire une faible transparence con-duisent à classer le lac de BASTANI dans la catégorie des lacs méso-trophes f statut trophique relativement rare pour ce type de milieu en
altitude.
Les données relatives aux invertébrés benthiques montrent que la faune des sédiments est constituée de chironomidés appartenant à la famille des orthocladiinae et chironominae ( tribu des tanytarsinii ) alors que les trichoptères des familles limnephilidae et polycentropodidae ( Plectroenemia corsicana ) sont rencontrés dans la zone rivulaire.
I H2 ORIGINE DES PEUPLEMENTS PISCICOLES. OBSERVATIONS ICHTYOLOGIQUES ANTERIEURES A L'INVENTAIRE.
Les données du présent chapitre sont issues des comptes rendus d'alevinage fournies par la Fédération des AAPP de la Corse, des précisions dont a bien voulu nous faire part M. le Président MARTINI au cours de communications personnelles et des observations effectuées par LEONI ( 1975 ) et ROCHE ( 1984 ) .
14
La seule référence aux peuplements piscicoles du lac de BASTANI antérieure à la deuxième guerre mondiale mentionne
l'exis-tence d'une population de truites autochtones : Salmo trutta macrostigma totalement détruite au cours de l'occupation italienne par les trou-pes qui a des fins alimentaires auraient utilisé les explosifs et dif-férents produits chimiques pour capturer ces poissons ( MARTINI,
communication personnelle ) .
En 1971 l'importante opération d'alevinage effectuée par héliportage inclut le lac de BASTANI où sont introduites au mois de septembre des juvéniles (o+) de truites communes ( Salmo trutta L.) et de saumons de fontaine ( Salvelinus fontinalis MITCHELL ) en prove-nance de la pisciculture de Vermenoux ( Nièvre ) et transportés sous oxygène et tranquillisant ( M S 222 ) en sacs contenant 20 litres d'eau, 5 kg de poissons et 3 1 de glace.
Le nombre total de poissons répartis sur l'ensemble des lacs étant connu : 6 000 saumons et 24 000 truites
fario, de même que la répartition par sac, on peut conclure en ad-mettant une distribution homogène des poissons à l'intérieur des sacs qu'ont été introduits sur le lac de BASTANI 1 200 saumons de fontaine et environ 5 000 truites d'une longueur voisine de 10 cm. Alors que ROCHE ( 1984 ) en se basant sur les chiffres précédents mentionne l'inversion à partir de 1980 du rapport des nombres de truites et de saumons dans les captures effectuées par les pêcheurs, il semble que les causes de l'absence actuelle de la truite fario résultent avant toute chose de l'état des poissons au moment de
l'introduction. Les observations du Président MARTINI montrent que le lac de BASTANI a été aleviné au cours de la dernière rotation d'hé-licoptère et que les truites qui apparamment avaient moins bien supporté le transport que les saumons de fontaine,ont eu à subir un taux de mortalité important.
15
IV RESULTATS
I V1 EFFORT DE PECHE MIS EN OEUVRE.
Quatre poses distinctes de filets, trois effectuées le 22 juillet et une le lendemain ont permis de prospecter l'ensemble du lac.
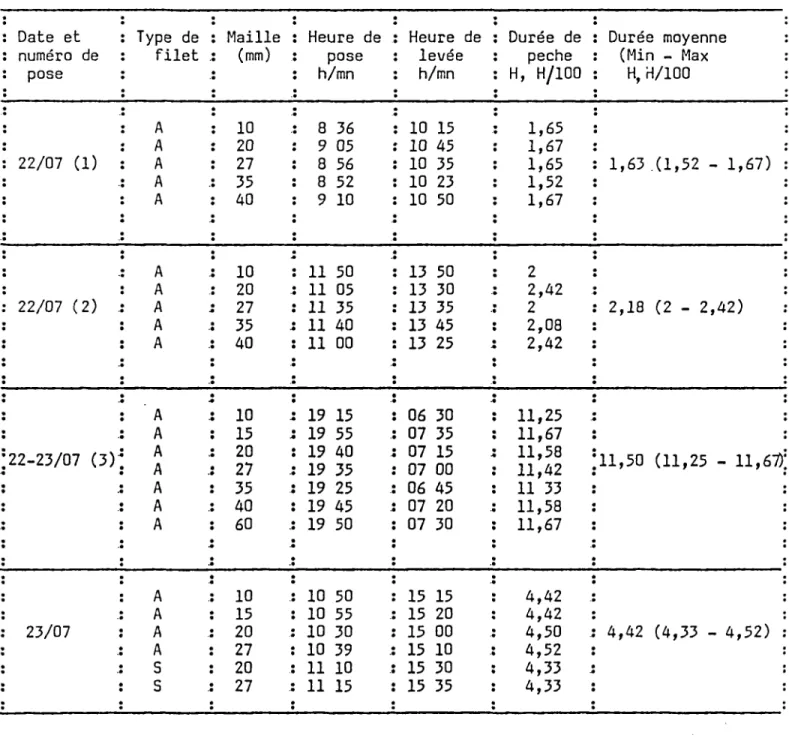
L'emplacement des filets pour chacune des pêches est indiqué sur la figure 3. Au cours de la première pose, désignée par ( 1 ) les filets ont été posés dans la partie Sud et Sud Ouest, la deuxième pose ( 2 ) concernant la rive Est. Devant la faiblesse des effectifs les filets posés sur la rive Ouest ( 3 ) ont été lais-sés en action durant la nuit et relevés le 23 juillet au matin. L'échantillonage a été complété dans la partie Nord et Nord Est du lac le 23 juillet. Les types de filets utilisés lors de chaque pose et les temps de pêche exprimés en heures et centièmes d'heures sont mentionnés dans le tableau 4
I V2 DEFINITION DE l'UNITE D'EFFORT DE PECHE.
Afin de pouvoir apprécier l'efficacité des filets de dif-férents types et de différentes dimensions de maille, ainsi que pour comparer les résultats obtenus dans le cadre du présent travail avec d'autres lacs, les résultats seront exprimés en fonction d'une unité conventionnelle : l'unité d'effort de pêche ( UEP ) . Les résultats correspondent aux effectifs numériques ou pondéraux qui auraient été capturés par un filet théorique d'une surface de Im2 péchant pen-dant une durée d'une heure. Les valeurs sont donc exprimées en nombres ou en grammes par mètre carré et par heure. Les nombres d'UEP par pose et par filet sont regroupés dans le tableau 5.
I U3 BILAN GLOBAL DES INVENTAIRES
Les pêches aux filets ont permis la capture de 47 saumons de fontaine ( Salvelinus fontinalis. MITCHELL ) représentant une
bio-16 : Date et : numéro de : pose : 22/07 (1) : 22/07 (2) .
•
• • ••
*
;22-23/07 (3)'*
•
• • • • • • • : 23/07 : Type de : filet: A
: A
: A
: A
: A
: A
: A
: A
: A
: A
: A
: A
: A
: A
A
: A
A
A
A
A
A
S
S
: Maille : (mm) : 10 : 20 : 27 : 35 : 40 : 10 : 20 : 27 : 35 : 40 : 10 : 15 : 20 : 27 : 35 : 40 : 60 i i »i 1 0
: 15 . 20 27 20• 27
»
: Heure de : pose : h/mn : 8 36 : 9 05 : 8 56 : 8 52 : 9 10 : 11 50 : 11 05 : 11 35 : 11 40 : 11 00i 19 15
: 19 55 : 19 40 : 19 35 : 19 25 : 19 45 : 19 50 • » i i ! 10 50 : 10 55 . 10 30 • 10 39 : 11 10 : 11 15 : Heure de : levée : h/mn : 10 15 : 10 45 : 10 35 : 10 23 : 10 50 : 13 50 : 13 30 : 13 35 : 13 45 : 13 25 : 06 30 : 07 35 : 07 15 : 07 00 : 06 45 : 07 20 : 07 30 i ! 15 15 : 15 20 15 00 15 10 • 15 30 15 35 : Durée de : peche : H, H/100 : 1,65 : 1,67 : 1,65 : 1,52 : 1,67 : 2 : 2,42 : 2 : 2,08 : 2,42 ! 11,25 : 11,67 : 11,58 : 11,42 : 11 33 : 11,58 : 11,67 > 4,42 . 4,42 4,50 4,52 4,33 , A,33 : Durée moyenne : : (Min - Max : : H, H/100 : : 1,63.(1,52 - 1,67) : : 2,18 (2 - 2,42) : ¡11,50 (11,25 - 11,67); 4,42 (4,33 - 4,52) :•
• • • • •Tableau 4 : Types de filets et durées de pêche pour chaque pose au cours de la campagne de 1986
17 ! Filets date et n° de pose 22/07(1) 22/07(2) 22.23/07(3) 23/07 ___ _ __ _ ____ nombre d'UEP par filet ________________j nombre d'UEP par maille AIO 49 i 60 i 336 132 _______ 577
577
A15 i 584 221 805 ________ 805 A20 i 120 1 174 834 324 _______ 1452 I S20 ' 225 _______ 225 ___——__1677
[ 1 A27 223 i i 1 270 _ _ _ _ _ j 1542 610 i 2645 f S27 f i j 585 i 585 j 3230r~ —
A35 266 ._r
i 364 i — 1983 i 2613 L______J 2613 ir
A40 141 L j 204 _____ 974 1319 L__——_—J 1319r
A60 L ___ _ 2509 L _ J 2509 L—__—J 2509 r 1 Nombre ' dîUEP par| pose j 799 | L j1072 j
_ j 87622097 ¡
L _ _12730 ¡
i |__ _, _ _ _ _ _ _ _| | | |Tableau 5 : Nombre d'unités d'effort de pêche par poses, par filet par maille.
18
masse de 7,41 kg. Ce premier résultat révélant l'existence d'un peuplement monospécifique pose donc le problème de l'opportunité d'introduction d'une ou d'autres espèces de salmonidés qui malgré le degré de trophie relativement élevé du lac n'apparaît pas souhaitable. En lacs d'altitude, la simplification extrême des chaînes alimentaires, la disponibilité de la nourriture seulement à certaines périodes de l'année ( émergence des nymphes de chirono-midés, poussées démographiques du zooplancton ) , la limitation des habitats physiques ; induisent nécessairement une compétition in-terspécifique qui risque de compromettre l'ensemble des peuplements piscicoles ( RIVIER 1985 ) .
I V4 TAUX DE SURVIE ET RESULTATS DES MARQUAGES
Les temps de pêche étant différents, suivant les poses de filets, les taux de survie seront analysés séparément pour chaque série. Les résultats sont rassemblés dans le tableau 6.
Les taux de survie sont relativement faibles : 34,04?á pour l'ensemble des pêches et nettement inférieurs à ceux obtenus sur les Ombles chevaliers et les truites des lacs de la vallée de Névache où les pourcentages de survie avoisinnaient 70 % pour des durées de pêche de 1 à 2 heures.
Dans le cas présent le meilleur taux de survie ( 50 % ) est obtenu pour la durée de pêche moyenne la plus faible : pose n° 1 ( temps moyen de pêche = 1,63 ) . Toutefois le nombre de poissons capturés ( N = 4 ) est trop faible pour être interprété de manière satisfaisante. Il n'existe ensuite pas de relation nette entre la durée de pose des filets et le taux de survie, le plus fort pourcen-tage ( 37,5 % ) étant obtenu pour la durée de pose la plus élevée : N° 3. Ces faibles valeurs montrent donc que le saumon de fontaine est une espèce relativement fragile et qu'il y aura lieu de tenir compte de ces observations lors des prochains inventaires de même
19 ¡ Date et n° de ' pose i ! 22/07 (1) i | 22/07 (2) i ¡ 22.23/07 (3)
j 23/07
| TOTAL Nombre de poissons capturés4
4
32 i i| 7
|
r
Í 4 7
Taux de survie i i( a ) ¡
50 |25 ¡
37,50 ¡
14 |
34 ¡
Tableau 6 : Pourcentage de survie des saumons de fontaine capturés sur le lac de Bastani.
20
qu'il sera utile de vérifier 1 importance du paramètre température qui influe nettement sur les taux de survie. Les faibles valeurs du taux de survie ne mettent cependant pas en péril la population des saumons de fontaine du lac de BASTANI. L'échantillon re-cueilli ne représentant avec l'effort de pêche mis en oeuvre qu'une infime partie des peuplements totaux.
Les problèmes liés aux taux de survie doivent être considérés en replaçant l'inventaire ichtyologique dans le contexte général de l'étude écologique du lac : le but de l'inventaire est la mise en évidence des principales caractéristiques des ichtyocénoses incluant l'étude de la croissance ( qui nécessite la lecture de l'âge des poissons ),de l'état de maturité sexuelle, des régimes alimentaires qui impliquent par conséquent le sacrifice d'une partie des sujets ; mais comprenant aussi le marquage et le retour au lac du plus grand nombre possible de poissons en vue d'études ultérieures : estimation des stocks par capture recapture, estimation de la mortalité par pêche, étude directe de la croissance.
En conséquence, suivant que l'inventaire sera unique ou au contraire appelé à être renouvelé, l'importance des prélèvements et des marquages sera susceptible de varier. Considérant que les pêches du lac de BASTANI se situaient dans la perspective d'une opé-ration unique, il a été décidé de prolonger la durée de pose des filets de manière à obtenir un échantillon représentatif et de pré-lever le plus possible de matériel pour les études en laboratoire tout en retournant au milieu la totalité des poissons survivants. Le lac de BASTANI fournit en définitive un bon exemple illustrant la préoccupation essentielle des ichtyologistes, lors des études de ce type, qui se situe dans la recherche constante du meilleur compro-mis entre l'efficacité de pêche que nécessite l'obtention d'un bon échantillon et la protection du patrimoine piscicole représenté par des espèces à haute valeur halieutique et écologique.
21
L'examen du taux de survie par dimensions de maille ( tab. 6 bis) montre que pratiquement tous les poissons capturés par les filets à maille de 10 mm survivent, que les pourcentages diminuent nettement pour les mailles de dimension moyenne ( 20, 27 mm ) et remontent ensuite pour la maille de 35 mm. Dans les filets à petites mailles, les poissons se prennent essentiellement par les dents et ne peuvent ensuite s'emmêmer dans le réseau dense des mailles dont l'ouverture n'autorise pas le passage des nageoires. Les poissons rencontrant les filets à maille de 20 à 27 mm se
prennent par les opercules puis s'emmêment alors que les captures effectuées avec la maille de 35 mm sont également dues au maillage strict, la grande dimension des mailles limitant l'emmêlage des poissons. Sans préjuger des rendements de pêche ( chapitre I V ) , les mailles de 10 et 35 mm apparaissent donc comme les plus intéres-santes à utiliser dans le cas où un pourcentage de survie élevé
est recherché.
Les caractéristiques biométriques et les numéros de marque des poissons identifiés au cours de l'inventaire sont ras-semblés dans le tableau 7.
IV,
. 1 BILAN GLOBAL PONDERELes valeurs des rendements numériques et pondéraux
respectivement de 0,004 et 0,582 sont relativement faibles. Toutefois, ces valeurs n'indiquent pas forcémentune rareté des poissons et sont sans doute liées à la dégradation des conditions météorologiques
dans la nuit du 22 au 23 juillet qui peut expliquer une diminution d'activité des poissons. Ces valeurs devront être confirmées ou in-firmées au cours de travaux complémentaires.
22 i Dimension de i maille (mm) L—_________ — _ — _ — -! 1 0 ! 1 5
¡
20
!
27 35 Nombre de poissons capturés ù2
17
14
8
i Nombre de poissons survivants i i i! 5
i0
4
4
Pourcentage i de survie i 83,3 ! 0 ! 23,5 | i i21,4 ¡
i i50
j
Tableau 6 bis : Nombre et pourcentage des poissons survivants en fonction de la dimension de maille.
Remarque : Deux saumons de fontaine survivants n'ont pas été marqués, ce qui explique les différences observées par rapport aux tableau n° 7.
23
¡ LF (mm)
! 250 208 243i
235
! 165i
215
i
237
¡
200
j
185
i
246
i¡
245
i | 235 iÌ
157
I
355
1 1 LT (mm) 260 218 251 247171
221 245 209 192 255 263 245 162 362P (g)
230 140 190 190 120 140 190 115 80 220 225 180 55 530 n° de marque EL 6463 EL 6456 EL 6776 EL 6777 EL 6778 EL 6779 EL 6784 EL 6785 EL 6789 EL 6790 EL 6791 EL 6793 EL 6794 EL 6709Tableau 7 : Caractéristiques biométriques et numéros de marque des saumons de fontaine relâchés après identification.
24
I V6 ANALYSE DES RESULTATS PAR DIMENSION DE MAILLE TU EFFICACITE COMPAREE DES DIFFERENTES MAILLES iV6.1
Les caractéristiques morphodynamiques du lac ont conduit à utiliser préférentiellement les araignées lors des inventaires, les filets de surface n'ayant été mis en oeuvre qu'au cours de la dernière pose et n'ayant capturé chacun qu'un poisson.
L'analyse ne portera donc que sur. les filets de type araignée. Les résultats bruts et pondérés des captures sont re-groupés dans le tableau 8. Les mailles de 40 et 60 mm dont la pose est nécessaire lors d'un premier inventaire pour vérifier la présence de poissons de grande taille sont caractérisées par l'absence de captures, alors que la maille de 20 mm apparaît comme étant la plus efficace aux plans numérique et pondéral, ce qui est en accord avec les observations effectuées précédemment sur les ombles chevaliers des lacs alpins ( RIVIER 1985 ) .
La maille de 10 mm présente ensuite une efficacité rela-tivement élevée liée principalement à la prise par les dents de poissons de taille élevée ( cf chapitres IV 4 et V 2 ) . Les autres filets n'ont ensuite qu'un rendement numérique très faible compris entre 0,003 et 0,004 poissons par UEP. La valeur du rendement pon-déral obtenue avec la maille de 27 mm : 0,994 g / UEP est à mettre en relation avec la prise par les dents de poissons de taille im-portante qui s'emmêlent ensuite très facilement dans les mailles de cette dimension.
TU SELECTIVITE iV6.2
Les nombres de poissons appartenant à chaque classe de longueur totale ( intervalle = 10 mm ) capturés par les différentes mailles sont représentés sur la fig 4 , les longueurs et poids moyens figurant dans le tableau 9.
25 NOMBRE 5 - i 4 -3 _ 2 -1 _
n
n
rQ
n
n
n
J~L
Un ihn
150 200 250 300 lQmm 15mm 20mmn n
27mm1 ,
35mm 1 1 1 350LONGUEUR
UOTALE(mm)
FIG 4: REPARTITION PAR CLASSES DE TAILLE DES SAUMONS DE FONTAINE CAPTURES
PAR LES ARAIGNEES DE DIFFERENTES MAILLES
26 i M P I 1 1 p (mm ^
! io
! 15 tI 2 0
iI 2 7
I 3 5
! 401 60
i Resultats bruts Nombre5
3
16 138
-—
Biomasse(g) 840 190 2371 2630 1130 Résultats NB/UEP 0,009 0,004 0,011 0,005 0,003-—
pondérés • iB/UEP (g) ¡
i i1,456 1
0,236 j
1,633 ! 0,994 ! i0,432 j
i i iTableau 8 : Nombres et biomasses bruts et par unité d'effort de pêche pour chaque dimension de maille.
nailles(mm)¡ moyennes LF (mm) LT (mm) P (g) 10
168 (- 66,85)
175 (- 66,24)
168 ( í 44,24) ¡63,33 ( í 75,90)
220 (Î40,65)
228,20 (t 42,63)
15 20¡208,38 (±29,96)
¡217,25 ( í 30,15)
¡148,19 (Î 75,94)
27 237,38(Í 26,82) 246,77(± 27,52 ) 202,31(î 67,17 ) 35 203,38(í 38,70) 213 (- 41,13 ) 141,25(^70,06 )Tableau 9 : Longueurs et poids moyens des saumons de fontaine capturés par chaque dimension de maille.
LF : Longueur à la fourche
LT : Longueur totale
P : Poids
( - X ) : Intervalle de confiance des moyennes au seuil 5?ó
28
Les mailles de 20 et 27 mm sont celles qui capturent la gamme la plus étendue des peuplements : longueurs totales com-prises de 160 à 362 mm pour la première et 155 à 342 mm pour la seconde . L'importance de la prise par les dents pour la maille
de 10 mm évoquée précédemment apparaît à nouveau clairement, la longueur totale moyenne des poissons capturés se situant à 228,2 mm et étant supérieure aux valeurs obtenues pour les mailles de 15,20 et 35 mm.
En résumé, les mailles de 20 et 27 mm sont celles
qui permettent le mieux d'apprécier l'étendue de la gamme des tailles et'qui sont par conséquent le mieux à même de renseigner sur la struc-ture réelle des peuplements. Ces filets en permettant un mode de cap-ture " mixte " : prise par les dents suivie d'emmêlage pour les
poissons les plus gros ou bien maillage strict pour les individus de taille plus réduite semblent être les mieux adaptés à l'inventaire des lacs peuplés de saumons de fontaine. La sélectivité nette de la dimension de maille observée par ailleurs pour les truites et ombles chevaliers des lacs d'altitude n'apparaît pas dans le cas du lac de BASTANI, les différences entre moyennes pour les longueurs et les poids n'étant pratiquement jamais significatives ( tableau 10 ) .
Ces résultats qui devront être confirmés par l'examen d'autres populations de saumons de fontaine peuvent être liés soit à la morphologie de l'espèce soit à l'existence de conditions locales particulières parmi lesquelles la faiblesse de la transparence des eaux peut constituer un facteur influençant fortement la nature des captures.
Dans le cadre d'un premier inventaire, il convient cependant d'utiliser une gamme de filets de dimensions de maille standard afin de ne pas risquer d'introduire un biais important dans
29 Paramètre comparaison 10.15 10.20 10.27 10.35 15.20 15.27 15.35 20.27 20.35 27.35
LF
NS
NS
NS
NS
NS
*
NS
NS
NS
NS
LT
NS
NS NSNS
NS
*
NS
NS
NS
NS
W
#•*N S i
NS
NS !
NS !
*#
NS !
NS
NS !
N S i
Tableau 10 : Comparaison des longueurs et poids moyens des saumons de fontaine capturés dans les araignées de différentes dimensions de maille.
Test t de STUDENT sur variables transformées ( X = Log LF, LT, W )
NS
-*•*
différence non significative au seuil 5% différence significative au seuil 5?ó différence significative au seuil 1%.
30
V) PRINCIPALES CARACTERISTIQUES DE LA POPULATION DES SAUMONS DE FONTAINE.
Vl REPARTITION DES EFFECTIFS PAR CLASSES DE TAILLE
Les histogrammes du nombre de lonqueurs totales appartenant à des classes d'intervalle de 10 ( fig 5 ) et 20 mm ( fig 6 ) révèlent l'existence de quatre groupes principaux de tailles (140 à 160 mm, 170 à 220 mm, 240 à 280 mm et au delà de 290 mm ) correspondant vraisemblablement à quatre classes d'âges distinctes (*). La population est donc relativement bien équilibrée, la dominance des poissons de longueur comprise entre 240 et 280 mm constituant un potentiel en géniteurs élevé. Les histogrammes ré-vèlent toutefois la faiblesse relative des classes de taille com-prises entre 140 et 180 mm dont il n'est pas possible après un seul inventaire de cerner avec précision les causes. Celles-ci peuvent être soit conjoncturelles car liées à la faiblesse du recrutement au cours des années précédentes à la suite de conditions météo-rologiques défavorables, soit explicables par la structure même de la population et par le phénomène de compétition intraspécifique qui se manifestant au détriment des juvéniles aurait tendance à entraîner un vieillissement de la population dont la cause peut être liée à une sous exploitation du lac par la pêche.
Les observations complémentaires effectuées lors des inventaires : captures d'alevins dans les zones de bordure au
(*) Les lectures d'âge par otolimètrie sont actuellement en cours et permettront de confirmer ou d'infirmer cette hypothèse ainsi que
d'établir par rétromesure et comparaison avec les valeurs obtenues dans l'échantillon, les modèles de croissance. Ces résultats ainsi que ceux relatifs aux régimes alimentaires feront l'objet d'une publication séparée.
31
8
7 _
6
5
4
-3 _
2
-1
NOMBRE 150nn mp
LONGUEUR TOTALE(mm) 200 250 300 350 40032
10
-NOMBRE 150 200I
250 300I
350 LONGUEUR TOTALE (mm)33
áu moyen de filets à plancton, observations sub-aquatiques ont mis en évidence la présence d'alevins de longueur totale moyenne d'en-viron 50 mm qui à l'examen du vomer peuvent être rattachés au genre salveninus et dont les otolithes permettent de préciser qu'il s'agit de poissons d'âge 0 . La reproduction des saumons de fontaine sur le lac de BASTANI est donc certaine, ce qui constitue la preuve que l'espèce peut accomplir la totalité de son cycle vital sur le lac et que la population est à même de se maintenir naturellement.
V2 ) RELATIONS LONGUEUR/FIASSE. COEFFICIENTS DE CONDITION. Les nombres exprimant la relation entre les masses et les longueurs à la fourche et longueurs totales des saumons de fontaine sont représentées sur Jes fig 7 et 8. Les données s'ajustent au modèle puissance caractéristique de la quasi totalité des poissons fusiformes ( TESCH 1971 ) :
M = aLb
avec M = Masse du poisson en grammes a = Masse du poisson unité
L = Longueur t o t a l e ou longueur à l a fourche en millimètre b = Coefficient d'allométrie
La valeur de b généralement voisine de 3 chez les salmonidés renseigne sur l e type de croissance qui est du type a l l o -métrique négative dans le cas présent ( b < 3 ) ce qui indique qu'au cours de l a croissance, l a longueur augmente plus rapidement que l a masse et que par conséquent l a c o n d i t i o n des p o i s s o n s e s t inversement c o r r e l l é e avec l a t a i l l e . La v a l e u r du c o e f f i -c i e n t d ' a l l o m é t r i e obtenue sur l e l a -c de BASTANI peut ê t r e considérée comme moyenne par r a p p o r t à c e l l e s rapportées par POWER ( 1980 ) qui c i t e pour l e s saumons de f o n t a i n e du c o n t i n e n t Nord américain des v a l e u r s du c o e f f i c i e n t d ' a l -l o m é t r i e comprises e n t r e 2,781 et 3,213.
RELATION CALCULEE AVEC LA LONGUEUR A LA FOURCHE MODELE PUISSANCE: U=A*L~B
I I I COEFFICIENT DE CORRELATION SUR VARIABLES TRANSFORMEES...0.93 I I I COEFFICIENTS A ET B A I 3 I 3.033613E-05 1 2.355 I
34
1 SOURCE DE VARIATION MOYENNE REGRESSION ECARTS TOTAL 1 1 1 1 1 DL. 1 1 45 47 1 1 1 1 1 S.CE. 1105.164 13.9731 .7545662 1124.397 1 1 1 1 1 CM. 1.676314E-02 « 9 9 339 „ 399 . 4 3 9 . *99 . 339 . 399 . S3« . 19« . is» . t o o . 3 9 . « I L O T I O U POtOSC « a U O M C U C U RS 2
¡f LU P0U>CH(/POISIJ
¿
*
i
t iì
í
L
1
m \ t i ? : ;FIG 7: RELATION MASSE/LONGUEUR A LA FOURCHE POUR LES SAUMONS DE FONTAINE
DU LAC DE BASTANI
RELATION CALCULEE AVEC LA LONGUEUR TOTALE MÚDELE PUISSANCE: U=A*L'B
35
i i I COEFFICIENT DE CORRELATION SUR VARIABLES TRANSFORMEES...0.979 I I I COEFFICIENTS A ET Q A I B I 2.22S403E-05 I 2.891 I 1 SOURCE DE VARIATION 1 MOYENNE 1 REGRESSION 1 ECARTS TOTAL 1 1 1 1 1 DL. 1 1 43 47 1 1 1 1 1 S.CE. 1103.164 18.93382 .7933472 1124.397 1 1 1 1 1 CM. 1.775216E-02 1 «lUDTION < a o _ ' r o x o«< « 93» . sa» . 41t . S3» . saa . as» . saa . taa . l a a . LOHCUfUR > • TOTAlC/POrOt / / -/ /
f
S 3 S
í t # /F.IG 8: RELATION MASSE/LONGUEUR TOTALE POUR LES SAUMONS DE FONTAINE
DU LAC DE BASTANI
36
En complément à l'étude des relations masse-longueur, l'examen de la condition des poissons qui représente une autre forme d'expression de la masse en fonction de la longueur, permet de carac-tériser l'état " d'embonpoint " des poissons qui est le reflet de leur activité nutritionnelle, de leur plus ou moins grande aptitude à utiliser les ressources alimentaires et en définitive de leur degré d'adaptation au milieu.
L'indépendance entre la condition et la croissance a d'ailleurs été mentionnée par ST PIERRE et MOREAU ( 1986 ) et POWER
( 1980 ) qui font remarquer que la disponibilité en nourriture peut conférer une bonne condition aux poissons sans leur assurer une crois-sance plus rapide.
En exprimant les longueurs ( L ) en centimètres et les masses ( M ) en grammes, la valeur du coefficient de condition ( K ) s'écrit :
K = 100 M/L3
avec L = Longueur à la fourche ou longueur totale.
Les valeurs moyennes de K obtenues avec les longueurs à la fourche et les longueurs totales sont respectivement de 1,392 et 1,228, valeurs qui dénotent une condition excellente pour les saumons de fontaine du lac de BASTANI. Ces valeurs représentent toutefois les maxima susceptibles d'être rencontrées au cours du cycle annuel : l'élévation de la température des eaux induit un taux de métabolisme élevé et une alimentation très active à partir de la faune d'origine zoobenthique et planctonique dont la production est alors maximale et disponible
(émergence des nymphes de chironomides, prolifération du zooplancton ) , alors que les apports exogènes ( faune terrestre ) constituent également une source importante de nourriture.
L'histogramme des coefficients de condition (*) répartis en classes arbitrairement fixées à 0,1 et la courbe des nombres cumu-lés de K ( fig 9 ) mettent en évidence l'homogénéité des coefficients
(*) Seuls les coefficients de condition calculés avec la longueur totale ont été considérés dans ce qui suit. Une valeur aberrante résultant d'une erreur de pesée ou de transcription a été éliminée, ce qui ramène le nombre d'observations à 46.
3 0
-
20-10 _
0.9
37
38
dont 87 % des valeurs sont comprises entre 1,1 et 1,4. La distribution obtenue montre donc que la valeur moyenne élevée résulte du bon état de condition de l'ensemble des poissons, ce qui sous entend vraisem-blablement un degré de compétition alimentaire et territoriale réduit entre les différentes classes d'âge.
Le calcul des coefficients de condition par classes de taille ne permet pas de déceler de variations nettes de ce para-mètre en fonction de la longueur ( fig 10.) ce qui correspond aux ob-servations de CARLANDER ( 1969 in POWER 1980 ) qui signale l'absence de relation entre les paramètres précédents.
La comparaison des coefficients de condition répartis en classes de taille (intervalle = 10 mm avec regroupement des cinq dernières classes) effectuée par analyse de variance non paramétrique : Test de
KRUSKALL-WALLIS sur les rangs,n'indique aucune différence dans la condition en fonction de la taille des poissons (Chi2 = 11,96, DL = 9, non signi-ficatif au seuil 5 ?¿ ) .
Le calcul des coefficients relatifs ( Kr ) qui repré-sentent les différences entre les poids réels et les poids théoriques calculés pour une longueur donnée à partir de la relation masse/longueur :
Kr = Mm - Mt ( Mt - aLb )
avec Mm = Masse observée Mt = Masse théorique
n'indique aucune tendance dans les variations de la condition en fonc-tion de la taille des poissons : les écarts positifs et négatifs ainsi que les valeurs absolues se répartissent de manière hétérogène-par rap-port aux longueurs classées ( tableau 10 ) . La comparaison des lon-gueurs et des coefficients de condition relatifs effectuée par calcul du coefficient de corrélation de rangs de SPERMANN met en évidence l'absence de corrélation entre les deux variables ( R = 0,205 : Non significatif au seuil 5 % )
39 Longueur totale ( m m ) 1*5 150 152 155 160 160 162 182 183 18* 185 185 189 190 192 195 196 197 207 209 209 218 220 Poids
(g)
30 50 55 50 50 55 55 70 72 70 7577
77
75
80 100 90 90 105 105115
1*0 100 Kr - 9,*39 + 6,500 + 9,802 + 2,175 - 2,*23 + 2,577 + 0,660 - 6,801 - 5,296 - 8,523 - *,763 - 2,763 - 7,852 - 11,156 - 8,80* + 7,125 - *,259 - 5,656 - 5,377 - 8,*88 + 1,512 + 11,800 - 31,629 Longueur totale ( m m ) 220 221 2*0 2*0 2*5 2*5 2*5 2*7 250 251 251 251 255 260 260 261 263 26* 278 29* 333 3*2 362 Poids(g)
120 1*0 170 175 170 180 190 190 200 200 220 190 220 230 200 210 225 2*0 255 280 470 500 530Kr
- 11,629 + 6,63* + 0,722 + 5,722 - 9,675 + 0,325 + 10,325 + 6,052 + 9,518 + 7,307 + 27,307 - 2,693 + 18,295 + 16,6*8 - 13,352 - 5,733 + *,*53 + 17,020 - 3,906 - 2 * , 369 + 33,690 + 28,721 -25,*37Tableau 10 :
Valeurs des coefficients de condition relatifs (Kr)
1.6
1.5
1.4
-1.3 _
1.2
-1.1 .
KT
LONGUEUR
' ' ' ' ' ' TOTALE(mm)
150 200 250 300 350
FIG 10: VALEURS MOYENNES DES COEFFICIENTS DE CONDITION PAR CLASSES DE TAILLE(intervalle 10mm)
41
V 3 ) SEX RATIO ET COMPARAISON DE LA CONDITION DES HALES ET DES FEMELLES.
Le sexage des poissons effectué in situ a révélé la présence dans l'échantillon de 16 mâles et 11 femelles, soit un sex-ratio de 1,45 en faveur des mâles. L'application de la règle des proportions binomiales aux valeurs expérimentales : calcul de l'écart normal ( Zc ) corrigé pour la continuité ne permet pas de montrer
l'existence d'un sex ratio différent de 1 ( Zc = 0,77 n = 27 non signi-ficatif).
La comparaison des coefficients de condition des mâles et des femelles obtenue par calcul du test t de STUDENT sur les loga-rithmes de K ( X = Loge ( K + 1) )ne montre aucune différence signi-ficative dans la condition liée au sexe des poissons ( t = 1,07 ) .
En définitive, les saumons de fontaine du lac de BASTANI sont caractérisés par une condition estivale élevée et par l'homogénéité de ce paramètre pour l'ensemble de la population. L'obtention de ces valeurs est le signe d'un degré de trophie relativement élevé pour un lac situé à cette altitude et de la bonne adaptation du saumon de fontaine au milieu.
V
4 ) CYCLE SEXUEL
Après une seule opération d'inventaire il n'est pas possible de décrire avec précision le déroulement du cycle sexuel des saumons de fontaine du lac de BASTANI et notamment de situer avec exactitude la période de reproduction. Les données suivantes, relatives au déroulement de la gamétogénèse et de la ponte sont issues de la bibliographie POWER ( 1980 ) . et simplement destinée à fournir des éléments qui devront être confirmés par des observations plus préci-ses sur le terrain.
42
V 4.1. CYCLE OVARIEN
La regulation du cycle ovarien est essentiellement contrôlée par photopériode, la température ne jouant vraisemblable-ment qu'un rôle secondaire ( POWER 1980 ) .
Le cycle ovarien comprend deux phases essentielles : génération des ovocytes et maturation.
1) Au cours de la première phase, les ovaires des femelles immatures contiennent de nombreux ovules de 0,1 à 1 mm de diamètre, auxquels s'ajoutent durant l'été de nouveaux oeufs qui atteindront la matu-rité au cours des années suivantes. Cette phase au cours de laquelle la croissance des ovules est lente se déroule durant les mois d'été.
2) Les ovules arrivant à maturité pour une année donnée se dévelop-pent rapidement à partir de juillet jusqu'à la ponte automnale au mo-ment de laquelle les oeufs à forte charge vitelline ont un diamètre d'environ 4 mm et présentent une coloration jaune orangée intense. Les oeufs arrivés à maturité sont directement libérés dans la cavité générale, puis déposés sur les frayères après passage dans un court conduit génital.
U 4.2. CYCLE SPERMATOGENETIQUE
Comme le cycle ovarien, la spermatogénèse est sous la dépendance de la photopériode, la phase finale étant liée à la décrois-sance de la durée de la phase diurne.
La première phase de la spermatogénèse consiste en la prolifération des spermatogonies qui se produit de janvier à mar3.
La spermatogénèse est ensuite peu active de mars à Juin/juillet, période
pendant laquelle la maturation des cellules germinales consiste en la division
43
mois d'août, le cycle sexuel s'accélère, les spermatozoïdes
commencent à gagner la lumière des lobules, les mâles arrivant à maturité de septembre à novembre et étant à même de se repro-duire durant 4 à 6 semaines.
Alors qu'en cours d'eau les lieux de ponte les plus favorables aux saumons de fontaine sont les fonds de gravier; POWER ( 1980 ) mentionne les substrats de sable propre au niveau des émergences souterraines comme étant propices à la reproduction de cette espèce. L'existence dans la portion ouest du lac d'un delta sous lacustre correspondant exactement à ce type de substrat est donc à même d'offrir des zones potentielles de frayeres impor-tantes.
V 4.3. FECONDITE ET ETAT DE MATURITE SEXUELLE
Les données de la littérature concernant la fécondité du saumon de fontaine exprimées en nombre d'oeufs par kg de femelle se situent à environ 2000 d'après SPILLMANN ( 1961 ) et à 2200 à 2500 d'après VLADYKOV 1956 ( in POWER 1980). Des tentatives de mesure de fécondité ont été effectuées sur le lac de BASTANI par pesée des opnades et comptage des ovules dans un sous échantillon, pour deux femelles de longueurs et poids indiqués ci-après :
Femelle Femelle
I
II
LT
294 342W
280 500 Nombre d'ovules 650 1110Les nombres totaux d'ovules par kilogramme de femelle se situent donc à respectivement 2321 et 2220 valeurs tout à fait comparables à celles rencontrées dans la bibliographie.
L'état de maturité sexuelle a été apprécié en divisant les femelles en trois stades basés sur le diamètre des
44
ovules, compte tenu des observations effectuées lors de la des-cription du cycle ovarien.
Stade I : Ovules de diamètre inférieur à 1 mm correspondant au stade immature.
Stade II : Ovules de diamètre compris entre 1 et 3 mm pouvant correspondre à des femelles n'ayant pas encore pondu pour la première fois.
Stade III : Ovules de diamètre supérieur à 3 mm arrivant à maturité à l'automne suivant.
La répartition des femelles à l'intérieur de ces trois catégories et représentée sur le tableau 11.
Il apparaît que les femelles dont la longueur totale est inférieure à 195 mm sont immatures, alors que celles en état de se reproduire dans l'année ont une longueur supérieure à 260 mm.
La gradation du stade de développement des ovules avec la longueur totale semble indiquer une périodicité annuelle de ponte pour les femelles ayant atteint la maturité. Les femelles dont la longueur est comprise entre 200 et 260 mm présentent des ganades en état de prématuration dont le développement des ovules est suscep-tible de se dérouler de deux manières différentes :
- l'ovogénèse chez les femelles de petite taille s'effectue plus tardivement que chez les poissons plus âgés ou se déroule plus len-tement et les oeufs atteignent la maturité la même année, mais avec un décalage dans le temps.
- les femelles dont les oeufs ont atteint le stade II présentent une phase hivernale de ralentissement ou d'arrêt de l'ovogénèse
suivie d'une reprise de développement l'année suivante donnant des oeufs arrivant à maturité.
45 ¡ longueur ¡ totale (mm) J 160
j 185
j 195
¡ 220
{ 245j 251
| 260 | 261i
278
i
294
j 3 4 2 I
i i i j Poigs (g) ——— ———-._——— _________ ! 5 5 ! 7 5 100 100 170 200 200 210 ¡i
255 | 280 ¡ 500 ! j stade de i j développement i i !i ¡
i í
ii ! ii ! II!
in |
II!
in |
in i
in !
Tableau 11 : Longueur, poids et stades de développement des ovules pour les femelles de saumon de fontaine du lac de BASTIANI
46
Les études en cours concernant la lecture des oto-lithes permettront de préciser l'âge de première reproduction et ainsi de décrire de manière plus approfondie le déroulement du cycle sexuel.
La fécondité totale des saumons de fontaine du lac de BASTANI , bien que ne pouvant être évaluée en l'absence d'une estimation du stock total , semble élevée si l'on se réfère à la distribution en classes de tailles des poissons et à la for-te représentation des génifor-teurs pofor-tentiels ( poissons ayant atfor-teint la maturité ou le stade de prématuration ) .
VI ) ORIENTATION DES TRAVAUX FUTURS
L'inventaire ichtyologique bien que constituant une phase
essentielle dans la connaissance des peuplements ne représente qu'une étape permettant de mettre en évidence les grands traits des peuple-ments et d'aborder un certain nombre de questions qui devront ensuite être résolues par des observations complémentaires.
Dans le cas présent, la population de saumon fontaine ne pré-sente pas de signes de déséquilibre et il y a donc seulement lieu de veiller à ce que cette situation puisse être maintenue.
Les travaux complémentaires sont donc à orienter suivant deux axes principaux : - meilleure connaissance de la biologie des saumons
de fontaine,
- évaluation de l'impact halieutique.
V I 1 ELEMENTS DE BIOLOGIE
A la suite de l'inventaire il apparaît indispensable de mieux cerner la biologie des premiers stades de développement
47
en effectuant des observations en période de fraie puis dès le dégel du lac.
Au cours de la première période, le recensement des secteurs de frayeres et l'évaluation du nombre des géniteurs qui les fréquentent permettra de préciser l'importance du potentiel de reproduction et de situer l'époque exacte de la reproduction ainsi que
son étalement dans le temps si les observations sont renouvellées durant plusieurs années.
Immédiatement après le dégel du lac l'observation des alevins constitue également un apport précieux en matière de biologie. Les éléments a prendre en compte concernent en particulier les zones de concentration des juvéniles et l'observation de leurs
caractéristi-ques aux plans biométrique et nutritionnel. En effet si en conditions natu-relles les pourcentages de fécondation sont élevés ( entre 80 et 90?¿), les données de la littérature mentionnent dans tous les cas des taux de mortalité importants durant les premiers mois de vie. La survie est influencée par l'époque d'eclosión et le début de la période d'ali-mentation elle-même conditionnée par la disponibilité des proies que
constituent certaines espèces zooplanctoniques. Le manque de synchro-nisme entre la période de fin de résorption de la vésicule vitelline suivie de l'émergence des alevins et la présence de ressources en nourriture est susceptible de conduire à des mortalités importantes qui ne sont qu'en partie compensées par le fractionnement dans le temps des pontes et par conséquent des dates d'éclosion.
L'observation de la taille des alevins permet enfin de préciser les dates d'éclosion alors que leur nombre constitue un mode d'évaluation du recrutement.
V I 2 IMPACT HALIEUTIQUE
S'il s'avère pratiquement illusoire de prétendre quantifier dans son ensemble l'impact de la pêche sur les
48
constituent une cause importante d'évolution des peuplements qui doit impérativement prise en compte dans le cadre d'une gestion rationnelle. Aussi, la sélection d'un certain nombre de pêcheurs pratiquant régulièrement sur le lac et acceptant de déclarer leurs captures pourrait constituer une approche dans ce sens. Les retours de marques d'un autre côté constituent un autre élément d'apprécia-tion de l'impact halieutique en même temps qu'ils permettent
d'appréhender directement les études de croissance si les poissons sont correctement mesurés et pesés et si les pêcheurs savent auprès de quel organisme ou association restituer leurs marques.
CONCLUSION
L'inventaire ichtyologique du lac de BASTANI a montré que les saumons de fontaine introduits en 1971 par la Fédération des AAPP de la CORSE s'étaient parfaitement acclimatés et consti-tuaient de nos jours un patrimoine piscicole et écologique de grande valeur.
Il ne semble pas souhaitable de diversifier les peu-plements en salmonidés en tentant par exemple à nouveau l'intro-duction de truites fario. En effet, malgré un degré de trophie assez élevé pour un lac d'altitude, la présence de plusieurs espè-ces de salmonidés a pour corollaire l'existence d'une compétition interspécifique tant au plan nutritionnel qu'à celui de l'habitat et si en terme de biomasse il importe peu que le stock soit repré-senté par une ou plusieurs espèces, il est préférable que le po-tentiel de reproduction ne soit pas fractionné entre plusieurs es-pèces.
Les actions à entreprendre en matière d'ichtyologie concernent seulement les compléments de biologie qui n'ont pu être envisagés dans le cadre d'une opération ponctuelle d'inventaire et le suivi halieutique.
La gestion piscicole du lac de BASTANI devra être essentiellement axée vers le maintien de l'équilibre actuel de la population : la reproduction naturelle du saumon de fontaine assurant la pérennité des peuplements. Il y aura seulement lieu de veiller au maintien d'un nombre suffisant de géniteurs pour que la structure des classes d'âge soit respectée. Ceci implique donc que l'on puisse évaluer l'importance de l'impact halieutique afin de décider si besoin est de mesures permettant ce maintien <» Parmi celles-ci sont envisageables les soutiens d'effectifs par de nouvelles introductions, la limitation du nombre des jours de pêche et des captures ou encore la mise en réserve du lac pendant des durées et à une périodicité permettant aux stocks de se re-constituer et à la population de se maintenir.
BIBLIOGRAPHIE
-50
BARBIER B.,1985. Les techniques de capture.Engins passifs:les filets maillants ,in D.GERDEAUX et R.BILLARD,Gestion piscicole des lacs et retenues artificielles,I.N.R.A. Paris,81-90
BRANDT A.V.,1975. Enmeshing nets:the theory of their efficiency,Symposium sur les méthodes de prospection,de surveillance et d'évaluation des
ressources ichtyologiques dans les lacs et les grands cours d'eau E.I.F.A.C. Tech.Pap. No 23(Suppl.l),Vol 1,96-116
CARLANDER K.D.,1969. Handbook of freshwater fishery biology.Vol.1.Iowa State Univ.Press,752pp.(in POWER 1980).
GAUTHIER A.,ROCHE B.,FRISONI G.F.,1984. Contribution à la connaissance des lacs d'altitude de la CORSE.Rapport C.R.D.P. Parc Naturel Regional de la CORSE,220pp.
LE CREN E.D.,BAGENAL T.B.,KIPLING C.,1975. Experiences with fish sampling methods
in Windermere.Symposium sur les méthodes de prospection,de surveillance et d'évaluation des ressources ichtyologiques dans les lacs et les grands cours d'eau E.I.F.A.C. Tech.Pap. No 23(Suppl.l),Vol 1,58-65 LEOPOLD M. et al.,1975. Effectiveness of gill net catches as a tool for the
estimation of fish populations in Polish lakes.Symposium sur les
méthodes de prospection,de surveillance et d'évaluation des ressources ichtyologiques dans les lacs et les grands cours d'eau E.I.F.A.C. Tech.Pap. No 23(Suppl.l),Vol 1,90-95
LEONI.,1975. Rapport P.N.R. CORSE.Non publié(in GAUTHIER et al.1984)
POWER G.,1980. The brook charr,Salvelinus fontinalis.in E.K.BAL0N,Charrs.Salmonid Fishes of the genus Salvelinus.W.JUNK Pub.,141-203
RIVIER B.,'1985. in Ecologie de quatre lacs naturels du Briançonnais.Tome 2: Peuplement piscicole.Rapport C.E.M.A.G.R.E.F.,D.P.N.,1-40
SAINT PIERRE M. et MOREAU G.,1986. Reproduction de l'omble de fontaine,Salvelinus fontinalis(Mitchell) dans des lacs de different ph.Hydrobiologia^ 141(3),237-248
SPILLMANN C.J.,1961. Faune de FRANCE,65.Poissons d'eau douce.Ed.Paul Lechevalier, 303pp.
TESCH F.W.,1971. Age and Growth.in Methods for the Assessment of Fish Production in Freshwater.I.B.P. Handbook No 3,Ed.W.E. Ricker,pp98-130,2nd Ed. Blackwell Scientific
VLADYKOV V.D.,1956. Fecundity of wild speckled trout(Salvelinus fontinalis) in Quebec lakes.J.Fish Res.Bd.Can.,13,799-841.(in Power 1980)