Dynamique d'infection des sous-populations lymphocytaires T CD4 chez le macaque Rhésus infecté par le Virus de l'Immunodéficience Simienne
Texte intégral
Figure
![Figure 1 : Origine des virus du Sida Figure tirée de Sharp et al., 2011 [2]](https://thumb-eu.123doks.com/thumbv2/123doknet/5586403.134142/18.918.104.764.96.639/figure-origine-des-virus-sida-figure-tirée-sharp.webp)
![Figure 2 : Prévalence mondiale du Sida en 2013 Figure tirée des données de Nations Unies sur Actualix.com [23]](https://thumb-eu.123doks.com/thumbv2/123doknet/5586403.134142/19.918.148.748.631.980/figure-prévalence-mondiale-figure-tirée-données-nations-actualix.webp)
![Figure 3 : Structure du virus VIH-1 Figure tirée de Gemrot et al., 2009 [49]](https://thumb-eu.123doks.com/thumbv2/123doknet/5586403.134142/22.918.115.781.127.479/figure-structure-virus-vih-figure-tirée-gemrot-al.webp)
![Tableau modifié à partir de Gobeil et al., 2012 [53]](https://thumb-eu.123doks.com/thumbv2/123doknet/5586403.134142/23.918.146.824.106.649/tableau-modifié-partir-gobeil-al.webp)
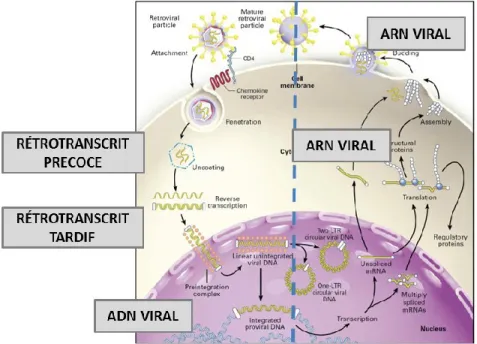
![Figure 6 : Étapes du cycle de réplication du VIH Figure modifiée à partir de Pasternak et al., 2013 [78]](https://thumb-eu.123doks.com/thumbv2/123doknet/5586403.134142/27.918.236.699.100.403/figure-étapes-cycle-réplication-figure-modifiée-partir-pasternak.webp)
![Figure 7 : Évolution typique de l’infection à VIH Figure tirée de [95]](https://thumb-eu.123doks.com/thumbv2/123doknet/5586403.134142/28.918.186.722.662.1011/figure-évolution-typique-l-infection-vih-figure-tirée.webp)
![Figure 9 : Interaction entre Tfh et cellule B au niveau du centre germinatif Figure tirée de Nutt et al., 2011 [134]](https://thumb-eu.123doks.com/thumbv2/123doknet/5586403.134142/33.918.151.801.137.491/figure-interaction-cellule-niveau-centre-germinatif-figure-tirée.webp)
Outline
Documents relatifs
Van Rullen (VanRullen, Carlson, & Cavanagh, 2007) montre que le traitement d’informations spatialement distribuées se fait de manière sérielle, les capacités
This method is preferably used in very large defects; thus, large free flaps such as the latissimus dorsi from the contralateral side, the free transverse rectus abdominis flap or
[r]
Dans un deuxième travail dont l’objectif principal était d’étudier le tissu synovial de 24 patients atteints de PR et traités par rituximab pour identifier
En effet, comme nous l’avons souligné dans notre partie qui concerne les hémoglobines hyperaffines, 6 patients sur 12 ont été diagnostiqués ces deux dernières années (2016,2017
C’est pour séduire le Mexique presque autant que l’opinion hispanique américaine, que le président Bush a lancé juste avant le Sommet de Monterrey le projet d’une
à la non-violation des articles 10 et 11 de la Constitution, estimant qu’il ne peut pas « être reproché au législateur de ne pas avoir prévu une catégorie distincte de
model (originally proposed on [23]), we illustrate how the proposed framework can be used in order to develop context models on different application domains in
