ETUDE DU MECANISME D'HABITUATION A LA THAXTOMINE A CHEZ LES CELLULES DE PEUPLIER
Par
Marie Girard-Martel
memoire presents au Departement de biologie en vue de l'obtention du grade de maitre es sciences (M.Sc.)
FACULTE DES SCIENCES UNIVERSITE DE SHERBROOKE
1*1
Library and Archives Canada Published Heritage Branch 395 Wellington Street Ottawa ON K1A0N4 Canada Bibliotheque et Archives Canada Direction du Patrimoine de I'edition 395, rue Wellington Ottawa ON K1A0N4 CanadaYour file Votre reference ISBN: 978-0-494-49506-3 Our file Notre reference ISBN: 978-0-494-49506-3
NOTICE:
The author has granted a non-exclusive license allowing Library and Archives Canada to reproduce, publish, archive, preserve, conserve, communicate to the public by
telecommunication or on the Internet, loan, distribute and sell theses
worldwide, for commercial or non-commercial purposes, in microform, paper, electronic and/or any other formats.
AVIS:
L'auteur a accorde une licence non exclusive permettant a la Bibliotheque et Archives Canada de reproduire, publier, archiver,
sauvegarder, conserver, transmettre au public par telecommunication ou par Plntemet, prefer, distribuer et vendre des theses partout dans le monde, a des fins commerciales ou autres, sur support microforme, papier, electronique et/ou autres formats.
The author retains copyright ownership and moral rights in this thesis. Neither the thesis nor substantial extracts from it may be printed or otherwise reproduced without the author's permission.
L'auteur conserve la propriete du droit d'auteur et des droits moraux qui protege cette these. Ni la these ni des extraits substantiels de celle-ci ne doivent etre imprimes ou autrement reproduits sans son autorisation.
In compliance with the Canadian Privacy Act some supporting forms may have been removed from this thesis.
Conformement a la loi canadienne sur la protection de la vie privee, quelques formulaires secondaires ont ete enleves de cette these.
While these forms may be included in the document page count,
their removal does not represent any loss of content from the thesis.
Canada
Bien que ces formulaires
aient inclus dans la pagination, il n'y aura aucun contenu manquant.
Le 27 novembre 2008
lejury a accepte le memoire de Mme Marie Girard-Martel dans sa version finale.
Membres dujury
Mme Nathalie Beaudoin Directrice
Departement de biologie
Mme Carole Beaulieu Membre
Departement de biologie
M. Kamal Bouarab President-rapporteur Departement de biologie
SOMMAIRE
La thaxtomine A (TA) est la principale toxine produite par Streptomyces scabiei, une bacterie filamenteuse du sol, qui est l'un des agents responsables de la gale commune de la pomme de terre. Le pouvoir pathogene de S. scabiei est dependant de la production de TA, laquelle est essentielle au developpement des symptomes de la maladie. A ce jour, la TA est connue comme un inhibiteur de la synthese de cellulose. En depit du role fondamental de la cellulose dans la paroi, il a ete possible d'adapter des suspensions cellulaires de peuplier hybride (Populus trichocarpa x Populus deltoides) a croitre et se diviser en presence de concentrations induisant normalement une hypertrophie et une mortalite cellulaire importante. Lors de ce processus d'habituation, les cellules de peuplier ont ete cultivees en augmentant progressivement la concentration de TA sur une periode de 12 mois. Depuis 4 ans, les cellules habituees a la TA sont desormais maintenues en l'absence de la toxine. Malgre tout, les cellules habituees a la TA sont encore deux fois plus tolerantes a la TA que les cellules non-habituees et sont moins sensibles a deux autres inhibiteurs de la synthese de cellulose : le dichlobenile et l'isoxabene. En reponse a la TA, les cellules habituees a la TA n'evitent pas la fragmentation de leur ADN nucleaire, une caracteristique de la mort cellulaire programmed qui est observable chez les cellules non-habituees. Cependant, le gonflement cellulaire est quasi absent chez les cellules de peuplier habituees a la TA, suggerant que leur paroi est alteree. En effet, celle-ci contient moins de cellulose cristalline et davantage de composes pectiques. Afln de mieux comprendre comment ces modifications de la paroi conferent une resistance a la TA, le profil d'expression genique des cellules habituees a la TA a ete etudie. Des changements prononces de leur transcriptome ont ete
detectes : 403 genes sont induits et plus de 865 genes sont reprimes (FC > 2,5). Les genes
dont l'expression est modifiee sont impliques dans differents processus incluant la synthese de la paroi et le metabolisme secondaire. De facon surprenante, aucun gene relie directement a la synthese de la cellulose n'a une expression modifiee, mais plusieurs genes impliques dans la synthese et le remaniement du reseau des hemicelluloses sont reprimes. Au niveau
du metabolisme secondaire, des genes codant pour des enzymes qui conduisent a la synthese des monolignols de la lignine, de meme qu'a la synthese des flavono'ides sont reprimes. Par rapport a la nature meme du mecanisme d'habituation a la TA, le nombre important de genes dont l'expression est modifiee revele Pimplication possible de changements epigenetiques, une hypothese appuyee par la possibilite de restaurer en partie la sensibilite des cellules habituees a la TA lors d'une exposition prolongee a la lumiere. Les cellules habituees a la TA s'averent done un outil utile pour comprendre les mecanismes de tolerance possible face a Tinhibition de la synthese de cellulose chez les vegetaux.
REMERCIEMENTS
En premier lieu, je tiens a remercier ma directrice de recherche, la docteure Nathalie Beaudoin, qui m'a donne initialement la chance de realiser un stage COOP dans son laboratoire, alors que je revais de travailler en biologie vegetale. C'est d'ailleurs grace a cette experience que j ' a i decide de poursuivre mes etudes a la maitrise. Un merci pour ta grande disponibilite, ton sens de l'ecoute et les conseils fort apprecies que j ' a i recus tout au long de ce projet. Un merci particulier pour 1'attention que tu portes a la reussite de tes etudiants, autant qu'a 1'amelioration de leurs points faibles. J'ai egalement beaucoup apprecie ma participation a deux congres scientifiques, soit a ACFAS et au Canadian Plant Genomics Workshop.
Merci a mes conseillers, la docteure Carole Beaulieu et le docteur Kamal Bouarab pour leurs conseils et l'interet porte pendant ce projet. Merci a Gilles Grondin pour son aide a la realisation des photos en microscopie.
Je remercie les membres presents (Helene, Marie-Andree, Madjid, Evguenia, Lydia) et passes (Viviane, Josee, Isabelle, Genevieve, Olivier, Mathieu, Jean-Pierre) de mon labo pour votre aide, vos conseils et votre compagnie! Merci egalement aux etudiantes et etudiants des labos voisins pour l'entraide et les nombreuses discussions (de sciences ou autre!!).
Merci a ma famille de Sherbrooke (Pat et Cat), mes amis et amies, ainsi que tous les coureurs et coureuses avec qui j'ai partage quelques kilometres, au sein du Vert et Or. Un dernier merci, et non le moindre, a mes parents, Isabelle et Alain. Malgre la distance (470 km de mon Lac natal), vous etes toujours disponibles pour m'ecouter et m'encourager dans les projets que j'entreprends.
Merci a vous tous, qui chacun a votre facon, m'avez permis de mener a bien cette maitrise et qui avez su faire naitre en moi des projets pour le futur des plus interessants.
TABLE DES MATIERES
SOMMAIRE ii REMERCIEMENTS iv
TABLE DES MATIERES v LISTE DES ABREVIATIONS viii LISTE DES TABLEAUX x LISTE DES FIGURES xi INTRODUCTION 1
A LA GALE COMMUNE DE LA POMME DE TERRE 1 A.l Symptomes et impacts de la gale commune 1 A.2 Agents pathogenes causant la gale commune 1 A.3 Implication des thaxtomines dans la pathogenese 3 A.4 Thaxtomine A et l'inhibition de la paroi cellulaire 6
B. PAROI DE LA CELLULE VEGETALE 10 B.l Importance de la paroi cellulaire 10 B.2 Structure de la paroi cellulaire 10 B.3 Composants de la paroi cellulaire 13
B.3.1 La cellulose 14 B.3.2 Lacallose 16 B.3.3 Les hemicelluloses 16
B.3.4 Les pectines 18 B.3.5 Leslignines 19 B.3.6 Les proteines parietales 22
C. HABITUATION DES CELLULES VEGETALES 23
C.l Habituation des cellules a l'isoxabene 24 C.2 Habituation des cellules au dichlobenile 24 D OBJECTIFS ET HYPOTHESES DE TRAVAIL 26
CHAPITRE1 28 MATERIEL ET METHODES 28
1.1 EXTRACTION ET PURIFICATION DE LA TA 28 1.2 CULTURE DES CELLULES DEPEUPLIER 28 1.3 HABITUATION DES CELLULES DE PEUPLIER A LA TA 29
1.4 TRAITEMENTS DES CELLULES 29 1.5 DETECTION DE LA MORTALITE CELLULAIRE 30
1.6 TESTTUNEL 30 1.7 MESURE DES DIMENSIONS CELLULAIRES 30
1.8 COLORATION DES PECTINES 31 1.9 EXTRACTION ET QUANTIFICATION DES PECTINES DE LA PAROI 31
1.9.1 Extraction des parois cellulaires 31 1.9.2 Extraction des pectines de laparoi cellulaire 32
1.9.3 Quantification des pectines de laparoi cellulaire 32
1.10 ANALYSE DE MICRO-PUCES A ADN 33
1.10.1 Extraction d'ARN totaux 33 1.10.2 Hybridation des micro-puces a ADN 33
1.10.3 Confirmation des profils d'expression genique par RT-PCR 34 1.10.4 Analyse informatique des genes dont l'expression est modifiee 36
CHAPITRE2 37 RESULTATS 37
2.1 CARACTERISATION DES CELLULES DE PEUPLIER HABITUEES
A LATA 37 2.1.1 Habituation a la TA chez les cellules de peuplier 37
2.1.2 Tolerance des cellules de peuplier habituees a la TA 39
2.1.3 Diminution de l'hypertrophie cellulaire 41 2.1.4 Resistance a 1TSOX et au DCB des cellules habituees a la TA 45
2.1.5 Fragmentation de 1'ADN 47 2.1.6 Modification de laparoi cellulaire 51
2.2 ANALYSE DE L'EXPRESSION DIFFERENTIELLE 53 2.2.1 Expression differentielle des cellules habituees a la TA 53
2.2.2 Resultats de 1'expression differentielle 54 2.2.3 Validation des resultats de micro-puces a ADN par RT-PCR 54
2.2.4 Identification des genes dont Pexpression est regulee 57 2.2.5 Analyse fonctionnelle des genes dont l'expression est regulee 57
2.2.6 Analyse de l'expression des genes impliques dans la paroi cellulaire 59 2.2.7 Analyse de l'expression des genes impliques dans le metabolisme
secondaire 62 2.2.8 Analyse globale des genes regules 64
2.3 IDENTIFICATION D'UN TRAITEMENT PERMETTANT UN RETOUR AU
PHENOTYPE SAUVAGE CHEZ LES CELLULES HABITUEES A LA TA 66
2.3.1 Augmentation de la sensibilite a la TA 67
CHAPITRE3 69 DISCUSSION 69
3.1 CARACTERISATION DES CELLULES DE PEUPLIER HABITUEES
A LATA 70 3.2 EXPRESSION DIFFERENTIELLE DES CELLULES DE PEUPLIER
HABITUEES A LA TA 72 3.3 IDENTIFICATION D'UN TRAITEMENT PERMETTANT UN RETOUR AU
PHENOTYPE SAUVAGE CHEZ LES CELLULES HABITUEES A LA TA 76
CONCLUSION 77 ANNEXE 1 79 ANNEXE 2 88 BIBLIOGRAPHIE 90
LISTE DES ABREVIATIONS
ADN acide desoxyribonucleique
ADNc acide desoxyribonucleique complementaire AGP arabinogalactane proteines
ARN acide ribonucleique
ARNc acide ribonucleique complementaire At Arabidopsis thaliana
CDT A cyclohexane-trans-1,2-diaminetetra-acetate CESA cellulose synthase
CHS chalcone synthase
CSL cellulose synthase potentielle, "cellulose synthase like" DAPI diamidino-4',6-phenylindol-2-dichlorhydrate DCB dichlobenile
EST etiquette de sequence exprimee, "expressed sequence tag" F3H flavonone 3-hydroxylase
FC "fold change"
FT-IR spectroscopic infrarouge a transformation de Fourier g gramme
GPvP proteines riches en glycines
HCT hydroxycinnamoyl-CoA shikimate/quinate hydroxcinnamoyl transferase
HGA homogalacturonane
HPLC chromatographic liquide de haute performance
HRGP glycoproteines riches en hydroxyproline ISOX isoxabene
kDa kilo Dalton
M molaire MCP mort cellulaire programmee
mg ml mm mM MS nm P pb PBS PCR PRP R/ RG-I RG-II RMA rpm RT-PCR TA TUNEL U UDP ug ul fun uM UV XET XTH milligramme millilitre millimetre millimolaire
milieu Murashige et Skoog nanometre
valeur p, valeur predictive paire de bases
tampon phosphate salin, "phosphate buffer saline"
reaction de polymerisation en chaine, "polymerase chain reaction' proteines riches en prolines
coefficient de migration rhamnogalacturonane-I rhamnogalacturonane-II
Robust Multichip Average methodology revolution par minute
"reverse transcription PCR" thaxtomine A
"terminal deoxynucleotidyl transferase-mediated dUTP nick end labelling" unite uridine diphosphate microgramme microlitre micrometre micromolaire ultraviolet xyloglucane endotransglycosylases xyloglucane endotransglycosylases/hydrolases
LISTE DES TABLEAUX
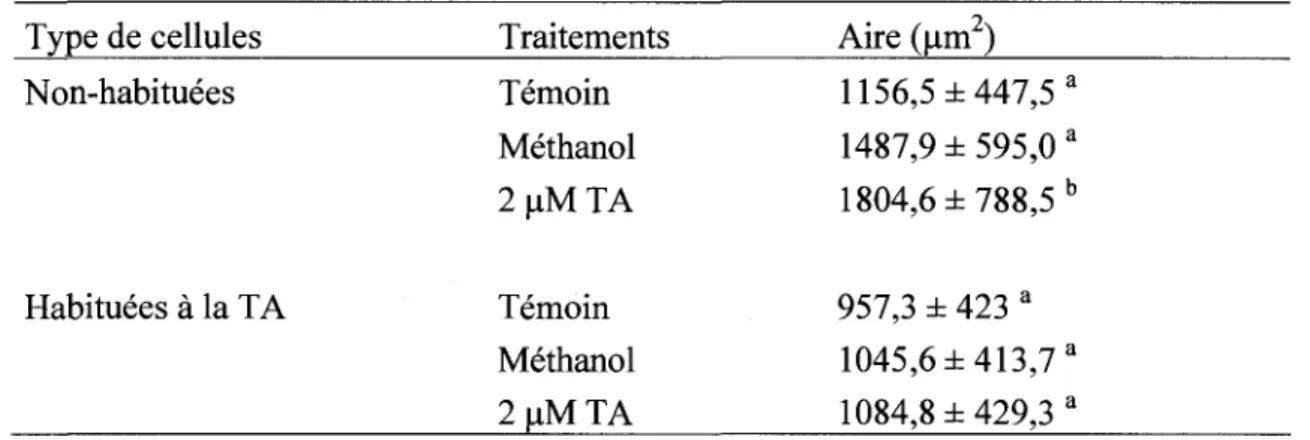
Tableau 1. Liste des amorces utilisees pour le RT-PCR 35 Tableau 2. Mesure des cellules de peuplier non-habituees et habituees a la TA apres un
traitement a la TA 44 Tableau 3. Genes impliques dans la synthese et la modification de la paroi cellulaire dont
l'expression varie chez les cellules de peuplier habituees a la TA 61 Tableau 4. Comparaison des genes impliques dans la synthese et la modification de la
paroi dont l'expression varie chez les cellules de peuplier habituees a la TA par rapport a ceux identifies chez les cellules d'A. thaliana habituees a
l'isoxabene 62 Tableau 5. Genes impliques dans le metabolisme secondaire dont l'expression varie chez
les cellules de peuplier habituees a la TA 64 Tableau 6. Genes relies a la synthese et la modification de la paroi cellulaire dont
l'expression est regulee (FC > 2,5) chez les cellules de peuplier habituees a la
TA 80 Tableau 7. Genes relies au metabolisme secondaire dont l'expression est regulee
LISTE DES FIGURES
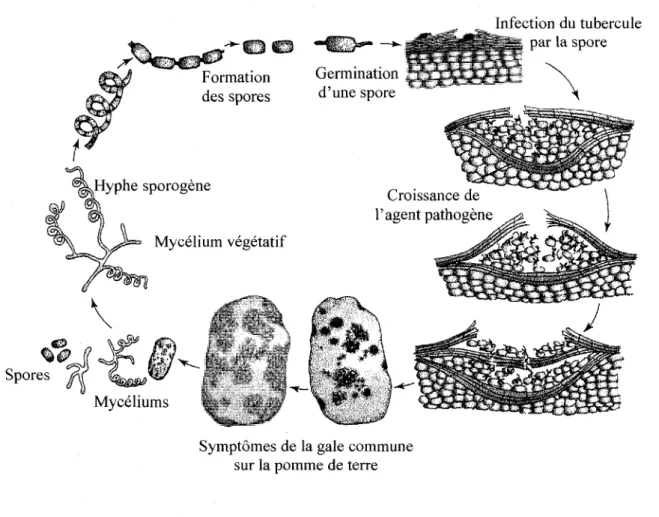
Figure 1. Cycle de la maladie de la gale commune de la pomme de terre causee par
Streptomyces scabiei 2
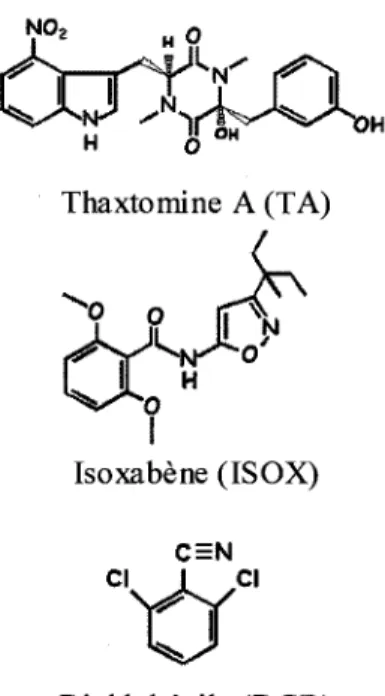
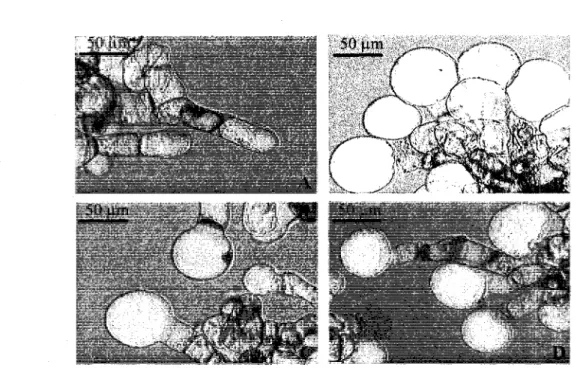
Figure 2. Structure de trois inhibiteurs de synthese de cellulose 4 Figure 3. Effets morphologiques de la TA, de 1'ISOX et du DCB sur les cellules
&A. thaliana 7
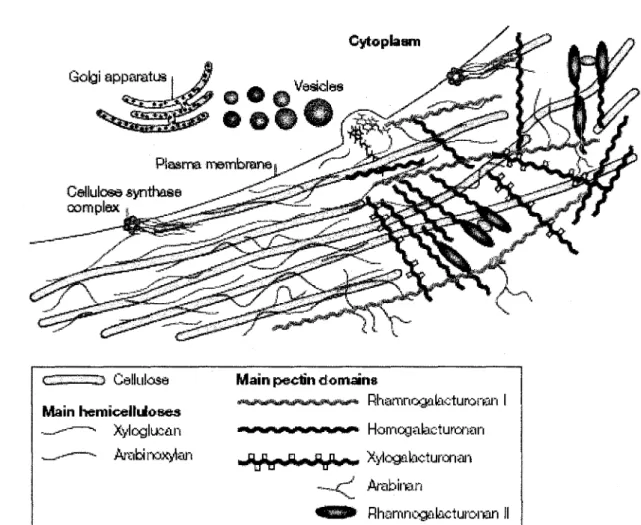
Figure 4. Structure et synthese de la paroi cellulaire vegetale primaire 12 Figure 5. Schema de la voie metabolique des lignines et des flavonoi'des chez
A. thaliana 21
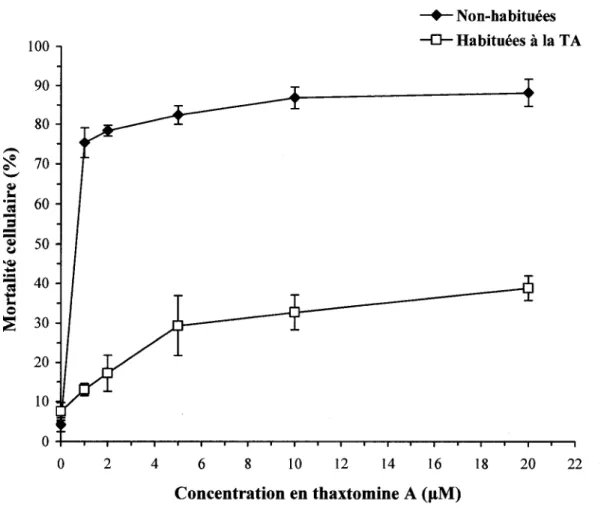
Figure 6. Mortalite des cellules de peuplier non-habituees et habituees a la TA,
apres 48 h de traitement avec differentes concentrations de TA 40 Figure 7. Morphologie des cellules de peuplier non-habituees et habituees a la TA,
apres 48 h de traitement avec la TA 43 Figure 8. Resistance des cellules de peuplier non-habituees et habituees a la TA,
apres 48 h de traitement a la TA, au DCB ou a l'ISOX 46 Figure 9. La TA induit la fragmentation de l'ADN nucleaire chez les suspensions
cellulaires de peuplier 49 Figure 10. Mortalite cellulaire evaluee par le bleu de trypan ou de noyaux positifs au
TUNEL, en presence de TA chez les cellules de peuplier temoins et habituees
a l a T A 50 Figure 11. Coloration des pectines chez les cellules de peuplier non-habituees et
habituees a la TA 52 Figure 12. Expression genique differentielle des cellules de peuplier habituees a la TA a
1'aide des micro-puces a ADN 55 Figure 13. Validation de la fidelite des micro-puces a ADN par RT-PCR 56
Figure 14. Classification des genes dont l'expression est regulee en categorie
Figure 15. Effet de la TA sur la mortalite des cellules de peuplier non-habituees ou habituees a la TA, en fonction des conditions de culture cellulaire, soit a
Pobscurite ou a la lumiere 68 Figure 16. Voie de synthese des flavonoi'des chez le peuplier 89
INTRODUCTION
A LA GALE COMMUNE DE LA POMME DE TERRE
A.1 Symptomes et impacts de la gale commune
La gale commune est une maladie importante de la pomme de terre (Solarium tuberosum L.) et de diverses cultures legumieres (betterave, carotte, radis, panais, navet) (Goyer et Beaulieu, 1997). Dans le cas des infections les moins severes, la maladie se manifeste par des lesions brunatres a la surface des tubercules. Avec le temps ces lesions superficielles peuvent s'elargir et devenir plus profondes, de 3 a 4 mm dans le tubercule, formant ainsi des lesions liegeuses et rugueuses (figure 1) (Agrios, 1997). La maladie reduit la qualite esthetique des racines-legumes, mais n'affecte pas leur qualite nutritionnelle (Babcock et al, 1993). Cependant, lorsque les lesions s'etendent a presque toute la surface du tubercule, cela reduit sa qualite marchande au point ou il devient impossible a vendre (Acuna et al, 2001; Loria era/., 1997).
A.2 Agents pathogenes causant la gale commune
En 1891, Thaxter a identifie l'agent causal de la gale commune qu'il a nomme Oospora
scabies, une bacterie filamenteuse du sol, desormais connu sous l'appellation de Streptomyces scabiei (Loria et al, 1997). Bien que la majorite des especes du genre des Streptomyces soit des saprophytes, S. scabiei est Pespece pathogene dominante dans le
monde entier (Loria et al., 2006). D'autres especes de Streptomyces peuvent egalement causer des symptomes similaires, par exemple S. acidiscabies est l'agent causal de la gale commune dans les sols acides (Lambert et Loria, 1989), S. turgidiscabies et S. aureofaciens
causent la gale commune comme S. scabiei (Goyer et al, 1996). Le cycle de la maladie se deroule en plusieurs etapes telles que detaillees dans la figure 1. Ainsi, l'agent pathogene peut se retrouver sous forme de mycelia ou encore de spores directement dans le sol ou sur des tubercules de semence contamines. L'infection par la spore survient habituellement au moment de la tuberisation de la pomme de terre.
Symptomes de la gale commune sur la pomme de terre
Figure 1. Cycle de la maladie de la gale commune de la pomme de terre causee par
Streptomyces scabiei.
L'agent pathogene penetre dans les tubercules en formation par les ouvertures naturelles (lenticelles) ou les blessures (Agrios, 1997). Suite a son entree, la bacterie se met a croitre entre les couches cellulaires entrainant une necrose au site d'infection du tubercule. La plante reagit ensuite a la presence du parasite par une proliferation de cellules liegeuses contenant un important depot de lignine. Cette couche cellulaire a pour but d'isoler le parasite et de le rejeter a l'exterieur de la plante, d'ou l'apparition des gales profondes ou surelevees. La maladie se developpe principalement sur les tubercules, les tiges souterraines et les racines. Puisque aucun pesticide homologue n'est efficace pour lutter contre cet agent pathogene, les strategies de lutte actuelles reposent principalement sur le choix de cultivars de pomme de terre presentant une resistance a la maladie et des methodes culturales appropriees, soit une bonne irrigation, un sol legerement acide et la rotation des cultures (Agrios, 1997).
A.3 Implication des thaxtomines dans la pathogenese
Dans le but d'elucider le mecanisme de virulence de S. scabiei, King et al. (1989) ont analyse la composition chimique des lesions de pommes de terre infectees par la bacterie. Leur etude a permis de relever la presence d'un groupe de phytotoxines, appelees thaxtomines, dont le role s'est avere essentiel dans le pouvoir pathogene de S. scabiei (Goyer et al, 1998; Healy et
al, 2000; Lawrence et al, 1990). En poursuivant leur etude sur les thaxtomines, ils ont
identifie au total 11 molecules de thaxtomines produites par S. scabiei, mais le compose predominant est la thaxtomine A (TA), une molecule de 4-nitroindol-3yl contenant un cycle
H 6 OM Thaxtomine A (TA)
Isoxabene (ISOX)
CI I CI
Dichlobenile (DCB)
Figure 2. Structure de trois inhibiteurs de synthese de cellulose. Modifiee de Corio-Costet et al (1991) et de Sheible et al (2003).
L'implication des thaxtomines dans la phytotoxicite de S. scabiei a ete correlee par plusieurs etudes utilisant differentes approches. Tout d'abord, l'application d'extraits de thaxtomines directement sur des tubercules de pommes de terre steriles mime les symptomes de la maladie (King et al, 1989; King et al, 1992; Lawrence et al, 1990). Au niveau de la plante elle-meme, les effets nefastes des thaxtomines ont ete detectes. Ainsi, la TA appliquee en faible concentration (0,05 a 0,1 uM) cause une augmentation dramatique du volume des cellules de differentes plantules dicotyledones et monocotyledones (Fry, 2004), tandis qu'a des concentrations elevees (50 a 100 uM), elle inhibe l'elongation cellulaire et cause la necrose des plantules (Leiner et al, 1996). Les etudes de King et al. (2001) et de Hiltunen et
al (2006) ont analyse les proprietes herbicides des differentes thaxtomines pour constater
qu'elles induisent des symptomes similaires, incluant l'arret et le gonflement des extremites racinaires, mais la TA est physiologiquement plus active que les autres molecules de
thaxtomines. Du cote de l'agent pathogene lui-meme, l'isolement de mutants de S. scabiei affectes dans la synthese de TA a permis d'etablir une correlation positive entre la quantite de phytotoxines produite et le pouvoir pathogene de Pactinomycete. Ainsi, deux etudes ont revele que des mutants de S. scabiei ne produisant plus de TA ne peuvent induire les symptomes caracteristiques de la gale commune (Goyer et ah, 1998; Healy et ah, 2000). Bien que certaines souches de Streptomyces ne produisant pas de TA puissent induire la gale commune (Wanner, 2004), la production de TA est habituellement correlee avec la severite de la maladie. Ces diverses etudes revelent principalement Pimportance de la TA dans la maladie. Cependant on peut se questionner sur les effets synergiques ou antagonistes qui pourraient exister entre les differentes thaxtomines. Un groupe de recherche s'est recemment interesse a ces interactions meconnues (Hiltunen et ah, 2006). lis ont observe que les 4 principales thaxtomines (la thaxtomine A, Pisomere ortho de la thaxtomine A, la thaxtomine B et la thaxtomin D) induisent des symptomes semblables sur des micropropagules de pommes de terre en culture in vitro, soit la reduction de la croissance des plantules et des racines, le gonflement racinaire et la necrose. Les thaxtomines A, B et D montrent des effets additifs, mais non synergiste lorsqu'elles sont appliquees en combinaison. De plus, les thaxtomines A et B ont un effet antagoniste avec l'isomere ortho de la TA.
Au niveau des facteurs controlant la synthese de TA, nous savons que la bacterie ne synthetise pas la phytotoxine en milieu minimal. Cependant, la TA est produite lorsqu'on ajoute des extraits de vegetaux dans le milieu de culture tel que le gruau ou le son d'avoine (Babcock et ah, 1993; Goyer et ah, 1998) et en milieu supplemente de pommes de terre (Beausejour et ah, 1999). Le compose activateur est la suberine puisque le milieu minimal
supplements en cellulose, pectate et amidon ne permet pas la biosynthese de TA, sauf lorsque
la suberine y est egalement ajoutee (Beausejour et ah, 1999). La suberine est un melange complexe d'acides gras hydroxyles et d'alcools produits par les cellules vegetales de facon a epaissir leur paroi, creant ainsi une barriere qui limite le transport de Peau et des nutriments et protege les plantes de Pinvasion par des agents pathogenes (Franke et Schreiber, 2007). Une etude recente (Johnson et ah, 2007) avance plutot que la cellobiose seule
(1-b-D-glucopyranosyl-4-D-glucose, le disaccharide de la cellulose) remplace les polymeres complexes d'origine vegetale dans 1'induction de la biosynthese des thaxtomines en culture in
vitro de S. acidiscabies et S. turgidiscabies. En culture in vitro, la production de TA par S. scabiei est sujette a une repression induite par le glucose, le tryptophane et la tyrosine
(Babcock et al, 1993; Lauzier et al, 2002). Nous connaissons mieux aujourd'hui les facteurs controlant la production de TA par les Streptomyces pathogenes bien que plusieurs aspects restent encore a etre elucides.
A.4 Thaxtomine A et l'inhibition de la paroi cellulaire
Les thaxtomines sont des dipeptides cycliques resultant de la condensation de deux acides amines methyles, la phenylalanine et le nitro-tryptophane, lies entre eux par un peptide synthetase bimodulaire d'origine non-ribosomale (NRPS): TxtA et TxtB (Healy et al, 2000). Au niveau structural, il a ete demontre l'importance pour la phytotoxicite de la portion phenyle de la phenylalanine et du groupement nitro associe au tryptophane (King et
al, 1989). Malgre l'importance des effets nefastes causes par la TA, son mode d'action
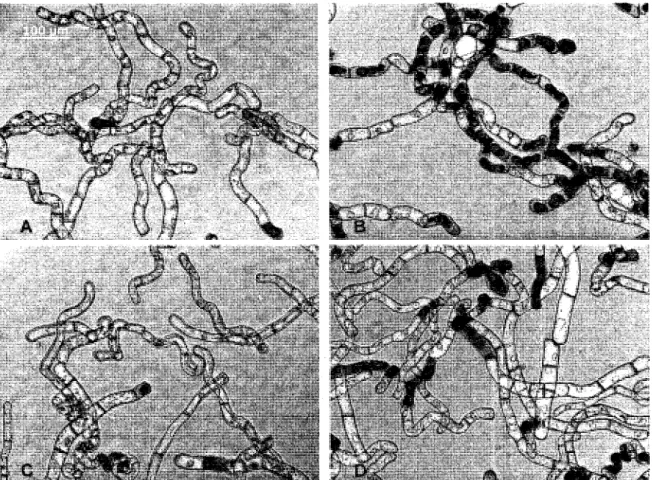
precis n'est pas encore identifie. Deux etudes ont avance la possibilite que la TA affecte la synthese ou la deposition normale de la paroi cellulaire vegetale (Fry, 2004; King et al, 2001), puisque les dommages causes par la TA sont fortement similaires a ceux occasionnes par des inhibiteurs de la synthese de la cellulose tels que le dichlobenile (DCB) et l'isoxabene (ISOX) (figure 2). Chez des suspensions cellulaires d'Arabidopsis thaliana, nous pouvons visualiser les effets similaires de ces trois inhibiteurs, soit l'hypertrophie marquee des
cellules ayant ete mises en contact pendant 24 heures avec la TA, 1'ISOX ou le DCB (figure
Figure 3. Effets morphologiques de la TA, de PISOX et du DCB sur les cellules d'A
thaliana.
Morphologie de cellules d'A. thaliana ayant ete en contact durant 24 heures avec (A) du methanol (comme temoin), (B) 2,0 uM de TA, (C) 2,0 uM d'ISOX et (D) 7,5 uM de DCB. Tiree de Brochu (2005).
Fry et Loria (2002) suggerent que la TA interagit avec une ou plusieurs cibles conservees chez les plantes, puisque la TA cause une augmentation drastique du volume cellulaire chez des hypocotyles de jeunes plants d'oignon et de radis, ainsi que le gonflement de cellules de tabac en suspension. L'inhibition de la synthese de la cellulose a ete mise en evidence chez
A. thaliana. L'equipe de Scheible et al. (2003) a montre une diminution de 1'incorporation
de glucose radiomarque dans la fraction de cellulose cristalline extraite de la paroi de plantules traitees pendant 24 heures avec de la TA. Par l'utilisation d'une autre methode permettant 1'analyse des parois cellulaires, la microspectroscopie infrarouge a transformation de Fourier (FT-IR), Robert et al. (2004) a appuye cette reduction en cellulose. L'etude de
Scheible et al. (2003) a egalement identifie un mutant d'A. thaliana resistant a la TA, txrl
(thaxtomin A resistant i). Chez ce mutant, ils ont detecte une diminution du taux
d'incorporation de TA par rapport aux cellules de plantules sauvages, demontrant que la proteine serait impliquee dans le transport de la TA dans les cellules. Le gene Txrl a ete identifie comme un gene codant pour une petite proteine de fonction inconnue n'ayant pas de motif apparent ni de signal de localisation specifique pour les organelles. Cette proteine a des homologues dans tous les genomes d'eucaryotes sequences dont plusieurs ne produisent pas de paroi cellulaire, ce qui suggere qu'elle ne fait pas partie du complexe de synthese de la cellulose. La mutation qu'ils ont identifiee n'est probablement pas la cible directe de la phytotoxine.
Des donnees recentes indiquent que la TA induit un flux d'ions rapide a travers la membrane plasmique, observations realisees chez les tubes polliniques et les racines d'arabidopsis et de tomate (Tegg et al, 2005). Ainsi, seulement une minute apres Pajout de TA, ils ont note un afflux de calcium transitoire au niveau de la membrane plasmique, de meme qu'une sortie de protons (H+) au niveau de tous les tissus analyses. On sait egalement que la TA active un
programme de mort chez des suspensions cellulaires d'A. thaliana (Duval et al, 2005) necessitant une expression genique active et la synthese de nouvelles proteines. Le type de mort cellulaire programmee qui est active en reponse a la TA chez les suspensions cellulaires
d'A. thaliana semble active des suites des dommages importants que subit la paroi sous
l'effet de la TA, de facon similaire a l'isoxabene. La mort cellulaire programmee (MCP) est definie comme une sequence d'evenements qui conduit a une destruction controlee et organisee de la cellule (Lockshin et Zakeri, 2004). II s'agit en fait d'un type de mort initie
par la cellule elle-meme, sous un controle genetique hautement regule (McCabe et Leaver,
2000). La MCP tient une place indispensable pendant le developpement des plantes, mais elle peut egalement etre activee lors de stress abiotiques et biotiques, afin de restreindre l'invasion des agents pathogenes (Reape et al, 2008). Chez les plantes, une forme de MCP confinee au site d'infection d'un agent pathogene se nomme la reponse hypersensible (Lam
du cytoplasme et de la chromatine dans le noyau (McCabe et Leaver, 2000). Les marqueurs biochimiques sont, quant a eux, 1'activation de certaines endonucleases (Williams et ah, 1974) et la fragmentation de l'ADN en fragments de 180 paires de bases (pb) correspondant a l'ADN compris entre deux nucleosomes (McCabe et Leaver, 2000). Cependant, la mort cellulaire en reponse a des stress peut-etre programmed ou necrotique. Par opposition a la MCP, la necrose est plutot decrite comme une forme de mort cellulaire incontrolee, souvent irreversible, qui survient habituellement lorsqu'il y a perte de l'integrite de la membrane plasmique et lors de stress ou les voies de la MCP n'ont pu etre activees. Durant la necrose cellulaire, on observe un gonflement du protoplasme plutot qu'un retrecissement decrit lors de la MCP. Ce gonflement est cause par la perte de la capacite d'osmoregulation de la cellule, ce qui a pour effet une entree d'eau et d'ions dans sa vacuole (Lennon et ah, 1991).
Bref, au cours des dernieres annees notre comprehension des mecanismes de virulence des actinomycetes causant la gale commune de la pomme de terre s'est de beaucoup enrichie. Ces connaissances sont d'autant plus importantes puisqu'une etude recente a identifie, en essai in vitro, des substances inhibitrices de la motilite des spermatozoi'des produites par des souches de S. turgidiscabies et S. scabiei (Kotiaho et ah, 2008). Maintenant que le genome de S. scabiei souche 87.22 est entierement sequence, d'autres decouvertes interessantes devraient ameliorer nos connaissances de cette maladie d'importance en agriculture. Au niveau de la plante, la cible cellulaire exacte de la TA est toujours inconnue, bien que nous sachions qu'elle altere directement ou indirectement la formation de la paroi primaire.
B. PAROI DE LA CELLULE VEGETALE
B.l Importance de la paroi cellulaire
La paroi des cellules vegetales est une matrice extracellulaire rigide et dynamique, contenant des biopolymeres parmi les plus abondants et complexes sur la terre (Lerouxel et al., 2006). Compte tenu des caracteristiques statiques des vegetaux, la paroi est une composante essentielle de la cellule vegetale. Contrairement aux animaux, ou le systeme squelettique s'est specialise pour assurer l'integrite structurale et l'appui physique de tout le corps, chez les plantes superieures la rigidite, la flexibilite et la forme globale dependent des proprietes cumulatives de toutes les parois qui entourent chacune des cellules (Farrokhi et al, 2006). Constituant une barriere physique a 1'entree et aux attaques des agents pathogenes, la paroi permet egalement a la cellule de resister a la pression de turgescence exercee par sa vacuole. La paroi a des fonctions importantes dans la croissance de la plante, la differentiation cellulaire, la communication intercellulaire, le mouvement de l'eau et la reponse de defense (Cosgrove, 2005; Pilling et Hofte, 2003). Des lors, nous ne pouvons qu'imaginer la complexity de la paroi. Loin d'etre une structure inerte, la composition et l'organisation de la paroi sont toujours en remaniement pendant le cycle cellulaire et en reponse aux differents stress biotiques et abiotiques (Farrokhi et al, 2006).
B.2 Structure de la paroi cellulaire
La paroi se compose d'un reseau de cellulose inclus dans une matrice de polysaccharides complexes, composee principalement d'hemicellulose et de pectines. Les microfibrilles de cellulose sont reliees entre elles par les hemicelluloses formant ainsi un reseau inclus dans un gel de pectines (figure 4) (Cosgrove, 2005). La cellulose et Phemicellulose conferent
principalement la structure et la rigidite a la paroi cellulaire, tandis que les pectines apportent plutot une plasticite (Reiter, 2002). La paroi contient egalement des proteines de structure, des enzymes et des polymeres phenoliques (Taiz et Zeiger, 2002). La paroi est impliquee dans les reponses de defense, par la formation rapide d'un reseau tres ramifie de proteines et la deposition de callose et de lignine creant une barriere physique pour prevenir 1'entree des agents pathogenes microbiens (Farrokhi et al, 2006). On distingue generalement deux types de paroi cellulaire : la paroi primaire, qui entoure la jeune cellule en croissance, et la paroi secondaire produite dans la cellule dont la croissance s'est achevee.
La paroi primaire. Dans les cellules en croissance active, la paroi primaire constitue une couche mince et flexible (0,1 a 1 urn d'epaisseur) composee en majorite par des polysaccharides complexes et quelques glycoproteins (Cosgrove, 2005). Le poids sec de la paroi primaire est compose approximativement de 25 % de cellulose, 25 % d'hemicellulose, 35 % de pectines et finalement, entre 1 a 8 % de proteines structurales (Taiz et Zeiger, 2002). Ces proportions changent d'un type cellulaire a Pautre, une plante ayant approximativement 40 types de cellules (Farrokhi et al, 2006), la composition des parois est done tres diversified. La croissance des plantes est un processus complexe. Les cellules vegetales etant etroitement liees ensemble, la migration des cellules n'est pas possible. La plupart du temps, la morphogenese des plantes est une question de divisions cellulaires localisees et d'une augmentation du volume cellulaire. Un tel accroissement peut etre impressionnant: les vaisseaux de xyleme comptent parmi les plus grandes cellules qui peuvent augmenter en volume jusqu'a 30 000 fois par rapport a leur stade meristematique (Cosgrove, 2005). Un autre exemple frappant est observe chez le coton ou les trichomes a la surface des jeunes graines s'allongent de 1 000 fois avant leur maturite. Ainsi, l'expansion cellulaire joue un role central dans le developpement de la biomasse d'une plante. En theorie, la biophysique expliquant l'expansion cellulaire est plutot simple : la cellule ne peut subir une expansion que lorsqu'un desequilibre provisoire est cree entre la pression hydraulique de sa vacuole et l'extensibilite de la paroi cellulaire ce qui fait en sorte qu'il y a une augmentation dramatique du volume cellulaire.
GZ——«3- Cellulose Main pectin domains ,»Mm*mamtfimtm0zmms0i* Rhamnogalacturoren 1 main hemioelliioses ._---—•» Xylogluean « M * * * » » W H » H * < « ^ Homogabcturorian —"-"™*- Arabinoxylan ^JffHBHJ4^J4nft^w Xytegalacturonan -*~-<~/ Arabinan 4§B9* Rhamnogalocturonan II
Figure 4. Structure et synthese de la paroi cellulaire vegetale primaire. Tiree de Cosgrove (2005).
L'elongation de la cellule n'est done permise que lorsque la rigidite de la paroi diminue permettant une plus grande prise d'eau dans la vacuole et que simultanement il y a ajout de nouveaux polymeres dans la paroi existante. En realite, l'elongation et la croissance cellulaire sont des processus beaucoup plus complexes et hautement regules necessitant plusieurs enzymes, puisque le relachement de la paroi requiert la rupture et le remodelage des interactions moleculaires entre une variete de composants de la paroi cellulaire. De nombreux agents sont impliques dans le relachement incluant les expansines, les
xyloglucanes endotransglycolases/hydrolases, les endo-(l,4)-p-d-glucanases, les "yieldines" et les radicaux hydroxyles (Humphrey et al, 2007). On a propose que les expansines etait une classe de proteines agissant comme agent de relachement de la paroi par leur capacite a cliver les liaisons hydrogenes entre la cellulose et les xyloglucanes. Les expansines et les xyloglucanes endotransglycolases/hydrolases (XTHs) sont des enzymes possiblement impliquees dans le relachement et le renforcement de la paroi. Les XTHs sont abondantes dans l'apoplasme (Rose et al, 2002) et la plupart des XTHs coupent des xyloglucanes qu'elles joignent a la microfibrille de cellulose adjacente (action xyloglucane endotransglucosylase). Seulement quelques XTHs utilisent l'eau comme substrat accepteur, ce qui resulte en Phydrolyse des xyloglucanes (action xyloglucane hydrolase), tandis que d'autres XTHs possedent les deux activites enzymatiques.
La paroi secondaire. Dans certains types cellulaires, une fois que la cellule cesse de croitre et qu'elle entame sa maturation, une paroi secondaire est deposee a l'interieur de la paroi primaire. La structure et la composition de la paroi secondaire definissent generalement l'etat de differenciation de la cellule (Reiter, 2002). Cette paroi est plus rigide et epaisse que la paroi primaire. La proportion des constituants de la paroi est egalement differente : 45 % de cellulose, un peu moins d'hemicellulose, peu de composes pectiques, la plupart des parois secondaires contiennent egalement des lignines qui peuvent representer de 25 a 35 % de la masse seche des tissus ligneux (Hopkins, 2003; Plomion et al, 2001).
B.3 Composants de la paroi cellulaire
La composition de la paroi est similaire chez les plantes superieures. Elle est generalement constitute de cellulose, de callose, d'hemicelluloses, de pectines, de lignines et de proteines parietales. La synthese de la paroi est egalement semblable chez 1'ensemble des plantes. La matrice de polysaccharides, d'hemicelluloses et de pectines est synthetisee dans l'appareil de
Golgi et secretee dans la paroi cellulaire, tandis que la cellulose et la callose sont synthetisees directement a la membrane plasmique (Hazen et al, 2002). Regardons en detail les composantes de la paroi cellulaire vegetale, ainsi que leur synthese et leur assemblage au sein de la paroi.
B.3.1 La cellulose
La cellulose est le polysaccharide principal de la paroi primaire et secondaire. La molecule de cellulose est un long polymere lineaire compose uniquement de cellobiose, un dimere de glucose lie en P-1,4 par des liaisons hydrogenes. Ce motif repetitif est constitue de deux residus de glucose, l'un droit et l'autre renverse. Selon le type cellulaire, une seule chaine de cellulose contient entre 500 et 15 000 molecules de glucose (Brett, 2000). Des leur synthese dans la paroi, les molecules de cellulose s'associent parallelement pour former les microfibrilles de cellulose. Cette structure cristalline aide la stabilisation des chaines de cellulose par le jeu de liaisons hydrogenes et de forces de Van der Waal's (Nishiyama et al, 2002). Ce type d'arrangement confere aux microfibrilles un caractere cristallin en meme temps qu'une grande solidite. En effet, la resistance a la rupture d'une microfibrille est impressionnante, soit equivalente a celle d'un fil d'acier de meme diametre (Hopkins, 2003). Le diametre des fibrilles de cellulose varie de 5 a 10 nm chez la plupart des plantes, jusqu'a 25 a 30 nm chez Valonia aegagropila et d'autres algues vertes (Ha et al, 1998; Herth, 1983). La cellulose est construite a l'exterieure de la cellule, dans l'apoplasme, par des complexes multi-proteiques transmembranaires. La microscopie electronique en cryofracture indique
clairement la presence de structures symetriques en rosette hexamerique
d'approximative-ment 25 a 30 nm de diametre a la membrane plasmique (Kimura et al, 1999). Ces rosettes, desormais appelees complexes terminaux, se sont averees etre associees a la partie terminale des microfibrilles de cellulose et responsable de leur synthese. Un complexe terminal est en fait constitue de six complexes, chacun compose de six unites de cellulose synthases. En se basant sur les mesures precises de la dimension des microfibrilles et leur taille estimee, Herth
(1983) a propose que chacun des six complexes d'une rosette peut synthetiser six chaines de p-l,4-glucane qui se cristallisent ensemble pour former une microfibrille de cellulose estimee a 36 chaines. Jusqu'ici, les seuls composants identifies en tant qu'element des complexes terminaux sont les celluloses synthases (CESA), une famille de glycosyltransferase. A l'origine, les CESA ont ete identifiees chez les plantes en comparant les sequences d'une librairie d'ADNc de fibres de coton avec les celluloses synthases d'origine bacterienne,
Acetobacter xylinum (Pear et al, 1996). Le genome d'A thaliana a revele 10 genes codant
pour des CESA, soit des proteines ayant une homologie de sequence de l'ordre de 64 % (Holland et al, 2000; Richmond et Somerville, 2000), alors que le peuplier compte 18 genes codant pour des CESA (Djerbi et al, 2005). Beaucoup de nos connaissances au sujet de la cellulose sont derivees de 1'identification de mutants deficients pour la synthese de la cellulose chez A. thaliana (Robert et al, 2004). Les mutants isoles ont des phenotypes tres differents, par exemple une diminution de la quantite de cellulose au niveau de la paroi primaire ou secondaire, des symptomes lies a la baisse de rigidite de la paroi, des defauts de croissance, du gonflement des racines, etc. Suites a de nombreuses etudes, il est clairement apparu que la synthese de la paroi primaire et secondaire, tout en partageant des composants communs, emploient un ensemble different de cellulose synthases (Taylor, 2008). Chez A.
thaliana, la synthese de la paroi primaire necessite la presence de trois isoformes de cellulose
synthases differentes dans le complexe terminal, AtCESAl, AtCESA3 et AtCESA6 (Somerville, 2006), tandis que l'assemblage de la paroi secondaire exige la presence d'AtCESA4, AtCESA7 et AtCESA8. L'orientation finale des microfibrilles dans la paroi n'est pas encore bien comprise. Nous savons que les microfibrilles de cellulose sont generalement deposees perpendiculairement a l'axe d'elongation de la cellule en croissance et de facon parallele aux microtubules du cytosquelette. Une etude recente a demontre chez
A. thaliana une association fonctionnelle entre la AtCESA6 et les microtubules corticaux
(Paredez et al, 2006), apportant une evidence de plus a l'hypothese que les microfibrilles de cellulose s'orientent a l'aide de microtubules, qui servent directement de rampes limitant le mouvement des complexes terminaux.
B.3.2 La callose
La callose est un polymere compose uniquement de glucose, ce qui en fait un polymere homogene tres similaire a la cellulose par sa structure. Mais, les glucoses de la callose sont plutot reunis en 0-1,3 ce qui confere a l'ensemble de la molecule une forme en helice a pas large (Guignard, 2000), contraire a 1'arrangement cristallin de la microfibrille de cellulose. Cette structure conduit a la formation de depots amorphes dans la paroi. Durant la croissance et le developpement des plantes, la callose est retrouvee transitoirement comme un composant normal de la plaque de division cellulaire. La callose est egalement un composant majeur de la paroi du pollen et du tube pollinique, en plus d'etre un composant structural du phloeme en dormance et des plasmodesmes. La callose joue un role central dans la prevention des invasions par les bacteries, differents champignons et les oomycetes pathogenes (Hardham et al, 2007). La synthese de la callose et sa deposition dans Papoplasme en reponse aux stress biotiques et abiotiques se produisent rapidement par l'activation des callose synthases, des complexes proteiques membranaires ayant un haut poids moleculaire (Kjell et al, 2004; Li et al, 2003).
B.3.3 Les hemicelluloses
Les hemicelluloses sont des polymeres beaucoup moins homogenes que les deux precedents. Par definition, les hemicelluloses sont des polysaccharides neutres ou legerement acides qui
possedent la caracteristique de pouvoir etre extraits de la paroi a l'aide de solutions alcalines
(Fry, 2004). La structure des hemicelluloses differe legerement entre les especes vegetales et selon le type de paroi, mais les principaux types d'hemicellulose sont les xyloglucanes, les arabinoxylanes et les mannanes. Les xyloglucanes sont les hemicelluloses les plus connus et les plus abondants de la paroi (Cosgrove, 2005). De facon generale, ils sont composes de chaines de glucoses lies en P-1,4 tout comme le squelette de la cellulose, mais presentent
egalement des ramifications formees de courtes sequences d'unites de xylose ajoutees sur l'oxygene 6 du glucose. Les xyloses peuvent former des embranchements avec des residus de galactose et de fucose. La presence de ces ramifications fait en sorte que leur structure empeche la formation de fibrille. Les xyloglucanes sont capables de lier deux microfibrilles de cellulose a la fois, par l'intermediaire de liaisons hydrogenes, assurant tout de meme a la paroi la cohesion entre la cellulose et les constituants ramifies de la paroi. Les arabinoxylanes sont, quant a eux, composes d'un squelette de D-xylanes lies en P-1,4 auquel s'ajoutent des embranchements d'arabinose. D'autres residus, tels que l'acide glucuronique et les esters d'acides feruliques peuvent egalement y etre attaches. Les mannanes sont un type d'hemicellulose peu connu contenant un squelette de mannanes retrouve dans la paroi primaire (Cosgrove, 2005).
Les hemicelluloses sont synthetises dans l'appareil de Golgi et sont par la suite transportes a la paroi a l'aide de vesicules d'exocytose. Les enzymes impliquees dans leur synthese, les cellulose synthases-like (CSLs), sont meconnues. Les CSLs ont ete initialement identifiees en se basant sur leur similitude avec le site actif des celluloses synthases connues et classees arbitrairement en six classes (AtCSLA, AtCSLB, AtCSC, AtCSLD, AtCSLE et AtCSLG). Les premieres analyses de localisation les positionnent au niveau de l'appareil de Golgi, tout comme le lieu de synthese des composes non cellulosiques de la paroi cellulaire (Richmond et Somerville, 2000). Temoignant de la diversite en structure des hemicelluloses, plusieurs CSLs sont requises, entre autres chez A. thaliana plus de 30 genes codant pour des CSLs sont connus (Hazen et al, 2002). Cependant, peu d'etudes ont demontre que les CSLs sont responsables directement de la synthese des hemicellulose. Une premiere etude (Dhugga et
al, 2004) a identifie une CslA et mis en evidence son activite p-mannane synthase pour la
formation du squelette de mannanes lorsqu'elle est exprimee chez des embryons somatiques de soya. Chez A. thaliana, une etude qui portait sur les genes de cellulose synthases-like a montre que les proteines de la famille des CSLA presentent une activite p-mannane synthase (Liepman et al., 2005). Recemment, des travaux realises sur le riz ont mis en evidence que le
gene CSLF serait responsable de la synthese des (l-3,l-4)-(3-D-glucanes (Burton et al, 2006). Cocuron et al. (2007) ont montre, chez A. thaliana, que le gene CSLC serait responsable de la synthese des P-1,4 D-glucanes composant le squelette de xyloglucanes. Ces differents resultats apportent de nouveaux elements en faveur de l'hypothese selon laquelle les proteines de la famille des cellulose synthases-like pourraient etre impliquees dans la synthese des polysaccharides non cellulosiques.
B.3.4 Les pectines
Les pectines sont un ensemble de polysaccharides heterogenes riches en acides galacturoniques qui sont lies ensemble par des liens covalents (Scheller et al, 2007). Les pectines sont definies comme les polysaccharides de la paroi qui sont solubilises par des tampons aqueux et des solutions diluees d'acide ou des chelateurs de calcium (Cosgrove, 2005). Les pectines sont pratiquement absentes de la paroi secondaire. Les trois principaux types de pectines de la paroi primaire sont les homogalacturonanes (HGA), les rhamnogalacturonanes I (RG-I) et les rhamnogalacturonanes II (RG-II) (Willats et al, 2001). Tout d'abord, les HGA sont des chaines lineaires d'acides D-galacturoniques liees en a-1,4 qui sont souvent methylesterifiees sur le groupe carboxyle en C6 et acetylees en C2 ou C3. Les RG-I sont des chaines lineaires contenant principalement des residus d'acides D-galacturoniques et de rhamnoses. II s'agit du type de pectines le plus abondant dans la paroi (Taiz et Zeiger, 2002). Differents substituants polysaccharidiques neutres sont capables de se greffer a ce squelette, formant ainsi de larges chaines laterales. Les RG-II ont une structure
beaucoup plus complexe puisque les chaines lineaires principales peuvent etre composees
d'au moins 12 differents monosaccharides. Les residus dans les RG-II incluent plusieurs monosaccharides qui sont rares dans les autres polysaccharides de la paroi, dont le D-apiose, le 2-O-methyl L-fucose, le 2-O-methyl D-xylose ou le L-galactose. La complexity des pectines suggere qu'un nombre important d'enzymes doit etre implique dans leur biosynthese. Plusieurs etudes ont montre que la biosynthese de pectine a lieu initialement
dans les vesicules de Pappareil de Golgi. Les enzymes requises dans ce processus sont principalement des glycosyltransferases et des enzymes ajoutant divers groupements, incluant des methyltransferases, des acetyltransferases et des feruloyltransferases (Scheller et
al, 2007). Bien que les fonctions importantes des glycosyltransferases soient maintenant
bien definies chez les mammiferes et les bacteries, un nombre relativement limite de glycosyltransferase a ete identifie chez les plantes. Seulement quelques genes responsables de la biosynthese des pectines ont ete caracterises jusqu'ici. Au moins 46 glycosyltransferases sont necessaires pour synthetiser les structures des polysaccharides pectiques qui sont trouvees dans HGA, RG-I et RG-II (Scheller et al, 2007). Malgre les nombreuses evidences que la synthese des pectines se deroule dans l'appareil de Golgi, nous ne pouvons pas exclure la possibility que les premieres etapes de biosynthese des pectines aient lieu dans le reticulum endoplasmique ou que certaines etapes de leur maturation aient lieu directement dans la vesicule de transport menant la pectine vers la paroi en formation. La complexity des pectines s'accentue meme une fois assemblee au niveau de la paroi puisque la structure fine de la matrice pectique est egalement modifiee pendant le developpement de la plante et en reponse aux conditions locales par differentes enzymes (Fry, 1995; Hadfield et Bennett, 1998). Ainsi, les pectines peuvent etre methyl-esterifiees a des degres variables, suggerant 1'implication de methylesterases agissant directement in muro (Fry, 2004). Les pectines de-esterifiees au niveau du groupement carboxyle sont capables d'agir comme chelateur des ions calcium ayant pour effet de renforcer la rigidite de la paroi.
B.3.5 Les lignines
Les lignines sont des polymeres phenoliques qui peuvent etre lies de facon covalente a certains polysaccharides de la paroi par des ponts benzyl ether (Wallace et al, 1995; Watanabe et al, 1989). L'arrangement des fibrilles de cellulose enchassees dans la lignine est responsable de Fextreme solidite du bois. Cette structure est tres solide, plus encore que les microfibrilles de cellulose (Hopkins, 2003). La lignine est composee principalement
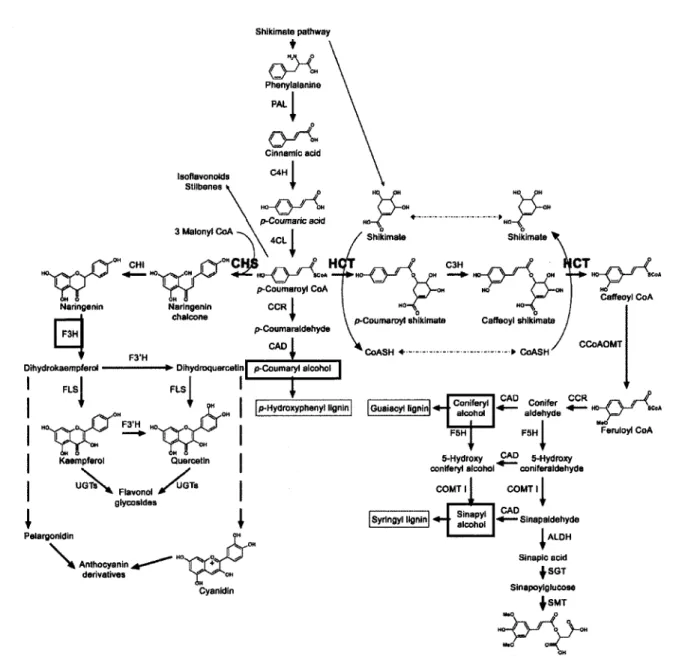
d'unites guaiacyl, syringyl et hydroxyphenyl qui sont les produits issus de la polymerisation de trois monolignols (ou d'alcools hydroxycinnamyliques) respectivement, l'alcool coniferylique, l'alcool sinapylique et l'alcool p-coumarylique qui sont encadres en rouge sur le schema de la voie des flavonoides et des lignines (figure 5). Ces monolignols sont synthetises a partir de la phenylalanine, puis secretes a la paroi ou ils sont oxydes sur place par des enzymes peroxydases et des laccases (des diphenols oxydases). L'etude de Boerjan
et al. (2003) a permis d'annoter plus de 34 genes candidats impliques dans la biosynthese des
monolignols et 27 genes apparentes. Cependant, la phenylalanine est egalement le precurseur de la voie de biosynthese des flavonoides. Ainsi, cette voie conduit non seulement a la lignine, mais egalement a un grand nombre de composes phenoliques tels que les anthocyanines, flavonols, flavones, isoflavonoi'des et proanthocyanidines, qui referent egalement aux tannins condenses (Miranda et al, 2007). Le />-coumaroyl-CoA est un intermediate metabolique ayant un role central et particulierement important puisqu'il constitue une plaque tournante entre les monomeres de lignine, les flavonoides et les autres phenols libres ou esterifies a la paroi en etant le substrat de deux enzymes, rhydroxycinnamoyl-CoA shikimate/quinate hydroxycinnamoyl transferase (HCT) d'une part, et la chalcone synthase (CHS) qui conduit a la branche des flavonoides d'autre part (Besseau et al, 2007).
Shikimale pathway t CHI Naringenin
H
Dihydrokaempferot — FLSl Isoflavonoids Stilbanes Shikimate X Naringenin ehalcone S C O * < p-Coumaroyl CoA CCR I p-Coumaraldehyde CAD I p-Coumaroyl shikimate ^CoASH + >—' Caffeoyl CoA Caffeoyl shikimate / / CCoAOMT * - Dihydroquercetlnf p-Coumaryl alcoholFLS
I J F3'H
Kaempferol
UGW I G W V ,-, . ,/vG\ * Flavonol 0
I |p-Hyd"»ypherty) lignin [ | Guaiacyl lignin| •• Conifery) alcohol
X
F5H
l
CoASH
aldehyde
F5H FeruloyI CoA 5-Hydroxy CAD 5-Hydroxy
coniferyl alcohol coniferaldehyde COMT Syrlngyl lignin I COMT I I „ „ i I CAD Sinapyl alcohol Cyanidin - Slnapaldehyde I ALDH Sinaplc acid | S G T Sinapoylglucose | S M T OH Sinapoylmalate
Figure 5. Schema de la voie metabolique des lignines et des flavonoi'des chez A.
thaliana. Abreviations: CHS, ehalcone synthase; HCT, hydroxycinnamoyl-CoA shikimate / quinate hydroxycinnamoyl transferase; ALDH, aldehyde dehydrogenase; C3H, C3-hydroxylase (CYP98A3); C4H, C4-hydroxylase (CYP73A5); F5H, ferulate 5-hydro-xylase (CYP84A1); CAD, cinnamyl alcohol dehydrogenase; CCoAOMT, caffeoyl-CoA O-methyltransferase; CCR, cinnamoyl-CoA reductase; CHI, ehalcone isom6rase; 4CL, 4-coumaroyl-CoA ligase; COMT I, acide caffeique O-methyltransferase de classe I; F3H, flavanone 3-hydroxylase; F3'H, flavonoi'de 3'-hydroxylase (CYP75B1); FLS, flavonol synthase; PAL, phenylalanine ammoniaque lyase; SGT, sinapate UDPglucose sinapoyltransferase; SMT, sinapoylglucose malate sinapoyltransferase; UGTs, "UDP sugar glycosyltransferases". Modifiee de Besseau et al. (2007).
B.3.6 Les proteines parietales
Les proteines localisees dans les parois sont omnipresentes dans les plantes et les algues vertes (Cassab, 1998). Les proteines parietales sont principalement constitutes par les proteines de structures et les proteines enzymatiques, dont les enzymes responsables de la synthese de la cellulose. Les proteines structurales sont habituellement riches en un ou deux acides amines, possedent des domaines ayant des sequences repetees et sont hautement glycosylees. Peu solubles et difficiles a extraire, leur role exact n'est pas clairement connu. D'apres leur composition en acides amines, il est possible de classer les proteines parietales en quatre categories : les glycoproteins riches en hydroxyprolines ou extensines (HRGPs), les proteines arabinogalactanes (AGPs), les proteines riches en glycines (GRPs) et les proteines riches en prolines (PRPs) faiblement glycosylees.
Certains mecanismes qui regissent la synthese de la paroi ne sont pas encore clairement elucides. En depit ou peut-etre justement en raison de son extreme complexity, la paroi des cellules vegetales possedent une grande capacite adaptative. Cette caracteristique interessante est relevee par les defis auxquels font face les agents pathogenes pour penetrer dans la plante ou encore la capacite de certaines cellules vegetales a survivre en presence d'inhibiteurs de la paroi cellulaire.
C. HABITUATION DES CELLULES VEGETALES
Depuis plus de 50 ans, il est connu qu'une variation des ratios des phytohormones cytokinine et auxine induit la formation de tissus particuliers chez les cellules vegetales, de cals indifferencies, des structures de tiges, de racines et meme de la plante entiere. La proliferation de cals indifferencies, le verdissement et la formation des structures de la tige sont tous des processus dependant des cytokinines. L'appellation "habituation" refere initialement au phenomene naturel par lequel des cultures de cals en passage continu perdent leur dependance face aux cytokinines. Plusieurs etudes ont montre un niveau anormalement eleve de cytokinines dans le contenu des cals, cette production intrinseque de la phytohormone explique possiblement leur independance vis-a-vis celle-ci (Pischke et al, 2006). Cependant, au cours des dernieres annees, le terme habituation est utilise pour d'autres processus similaires, dont l'habituation de cellules vegetales a differents inhibiteurs specifiques et ce, par contact progressif avec l'inhibiteur. En principe, l'habituation chez des cellules vegetales peut etre le resultat de mutations, de changements epigenetiques ou d'une combinaison de ces deux processus (Meins, 1989). Des etudes anterieures ont ainsi demontre la possibilite d'adapter ou d'habituer des cellules vegetales a se developper et se diviser en presence d'inhibiteurs de la synthese de la paroi, tel que le dichlobenile, l'isoxabene et le quinclorac (Alonso-Simon et al, 2008; Corio-Costet et al, 1991; Diaz-Cacho et al, 1999; Encina et al, 2001; Encina et al, 2002; Garcia-Angulo et al, 2006; Manfield et al, 2004; Shedletzky et al, 1990; Shedletzky et al, 1992). Dans l'ensemble des etudes, les differents types de cellules sont rendus resistants en ajoutant des concentrations croissantes de l'inhibiteur dans le milieu de culture pendant plusieurs generations et ce, jusqu'a des
C.l Habituation des cellules a l'isoxabene
L' isoxabene (TV- [3 -(1 -ethyl-1 -methylpropyl)-5-isoxazoryl] -2,6-dimethoxybenzamide ou ISOX) est un inhibiteur specifique de la synthese de cellulose. Deux mutants d'A thaliana resistants a l'isoxabene ixrl et ixr2 ("isoxaben resistant") (Desprez et at, 2002; Scheible et
at, 2001) ont ete caracterises, les mutations etant localisees respectivement dans les genes CESA3 et CESA6. Ces deux genes codent pour des isoformes de cellulose synthase. De
1'analyse de ces mutants, nous pouvons en deduire que l'isoxabene perturbe un site de regulation ou interfere avec l'assemblage final du complexe terminal de cellulose synthase par l'inhibition du transport des vesicules contenant les sous-unites catalytiques de CESA3 et CESA6. Afin de mieux caracteriser les effets d'une diminution du reseau de cellulose, des groupes de recherche ont egalement montre la possibilite d'habituer des cellules vegetales a de fortes concentrations d'isoxabene. L'habituation de cals de haricot et de suspensions cellulaires d'A thaliana a 1'ISOX a revele que la diminution du reseau cellulosique peut etre compensee par l'augmentation du reseau pectique (Diaz-Cacho et at, 1999; Manfield et at, 2004).
C.2 Habituation des cellules au dichlobenile
Le dichlobenile (2,6-dichlorobenzonitrile ou DCB) est connu comme un inhibiteur tres efficace de la synthese de la cellulose depuis plus de 30 ans (Hogetsu et at, 1974). Malgre
les nombreuses etudes realisees depuis, son mode d'action precis demeure inconnu.
Cependant, l'etude de DeBolt et at (2007) a apporte de nouvelles connaissances quant aux effets du DCB. Leurs experimentations ont montre que le DBC inhibe rapidement la motilite du complexe YFP::CESA6 au niveau des microtubules corticaux chez des cellules d'A
thaliana et cause une accumulation des complexes de CESA en des sites distincts dans le
pour consequence de ne plus detecter le complexe YFP::CESA6. Ainsi, le DCB semble agir sur une cible differente de PISOX. D'un autre cote, plusieurs etudes ayant habituees differents types de cellules vegetales au DCB ont revele la capacite d'adaptation impressionnante des cellules aux effets nefastes de cet inhibiteur. Parmi les premieres etudes, il y a celles de Shedletzky et al (1990 et 1992) qui ont habitue des suspensions cellulaires de tomates (Lycopersicon esculentum VF 36) a une forte concentration en DCB. Le mecanisme d'habituation de ces cellules resulterait dans leur capacite remarquable a survivre et se diviser pratiquement en l'absence de cellulose. Ensuite, Sabba et al. (1999) ont egalement note que la paroi de cellules de tabac (BY-2) habituees au DCB est grandement alteree. Ces derniers ont observe une diminution du contenu en cellulose et une augmentation des pectines, des extensines et de la callose. Plusieurs etudes realisees sur des cals et des suspensions cellulaires de haricot (Phaseolus vulgaris L.) habitues au DCB ont egalement montre que leur paroi cellulaire contient moins de cellulose et plus de pectines (Alonso-Simon et al., 2004; Alonso-Simon et al, 2007; Encina et al, 2001; Encina et al, 2002).
Ces differentes etudes et leurs analyses biochimiques de la paroi tendent a montrer que lorsque la synthese de la cellulose est modifiee progressivement par l'ajout d'inhibiteur specifique, la cellule est capable de detecter ces variations et de compenser les effets negatifs de la perturbation du reseau cellulosique. La nature dynamique de la paroi ne peut etre expliquee que par 1'existence d'un systeme pour la detection, la signalisation et la retroaction permettant le maintien d'une paroi fonctionnelle (Humphrey et al, 2007). A ce jour, les meilleurs candidats potentiels comme senseurs de l'integrite de la paroi cellulaire sont les canaux mechanosenseurs au calcium, differents recepteurs kinases localises a la membrane
plasmique, dont les "wall-associated kinases" (WAKs), les "lectin receptor kinases"
(LecRKs) et les "proline-rich extensin-like receptor kinases" (PERKs), en plus des "proteins leucin-richs extensins" (LRX) et les arabinogalactane proteines (AGPs). Bien que certaines variations soient detectees entre les especes, la reponse adaptative des cellules vegetales est habituellement la synthese d'une paroi cellulaire ayant une architecture radicalement modifiee, soit riche en pectines et en certaines proteines parietales.
D OBJECTIFS ET HYPOTHESES DE TRAVAIL
A ce jour, peu de mecanismes de resistance a la gale commune ont ete decrits au niveau moleculaire chez des cellules vegetales. Cependant, comme mentionne precedemment, 1'adaptation de cellules vegetales a des concentrations mortelles en inhibiteur de synthese de cellulose peut etre utile dans l'etude des effets a long terme de l'exposition aux herbicides et fournir de meilleures connaissances sur leurs modes d'action. Ce projet de maitrise etait done consacre a l'etude du mecanisme de resistance a la TA chez des cellules de peuplier habituees de facon progressive a cette phytotoxine. L'etude a ete debutee precedemment par Viviane Brochu, une etudiante a la maitrise dans notre laboratoire. Son projet avait pour but de caracteriser les effets de la TA chez les plantes afin d'identifier sa cible specifique chez les cellules. L'un des objectifs de son projet etait d'elaborer des cellules de peuplier habituees a la TA. Ces cellules "habituees" ont ete cultivees pendant un an en presence de concentrations croissantes de TA et depuis 4 ans, ces cellules sont maintenues en 1'absence de la phytotoxine. Nous chercnons a comprendre comment les cellules de peuplier habituees a la TA reussissent a survivre et a se diviser en presence de concentrations normalement mortelles de TA et de plus, a savoir comment elles maintiennent leur habituation dans le temps et ce, en l'absence de la phytotoxine. Les analyses realisees par Brochu (2005) ont confirme la diminution de la quantite de cellulose chez des cellules vegetales en presence de TA. Tandis que l'analyse des cellules habituees a la TA, par la spectroscopic infrarouge a transformation de Fourier (FT-IR), indique que leur paroi est enrichie en pectines et reduite en cellulose cristalline. Dans le cadre de mon projet, sachant que la paroi des cellules habituees a la TA est modifiee et que ces modifications sont maintenues pendant une longue
periode de temps, j ' a i teste Phypothese que les cellules habituees a la TA ont une expression
differentielle de certains genes par rapport a des cellules temoins, leur permettant ainsi de conserver une paroi alteree qui leur confere une resistance aux effets toxiques de la phytotoxine.
Les 3 objectifs specifiques de 1' etude etaient les suivants :
1. Caracteriser morphologiquement et physiologiquement des cellules habituees a la TA. 2. Caracteriser l'expression differentielle chez les cellules habituees a la TA par rapport
aux cellules non-habituees.
3. Identifier un traitement permettant un retour au phenotype sauvage chez les cellules habituees a la TA.
CHAPITRE 1
MATERIEL ET METHODES
1.1 EXTRACTION ET PURIFICATION DE LA TA
La TA a ete extraite et purifiee de cultures de Streptomyces scabiei souche EF-35, comme decrit par Goyer et al. (1998). S. scabiei a ete cultivee en milieu son d'avoine sous agitation a 30 °C, durant 5 a 8 jours. Une fois les cultures centrifugees, le surnageant a ete extrait deux fois avec de l'acetate d'ethyle. La TA a ete concentree par evaporation du solvant, puis purifiee par chromatographie en couches minces en utilisant des plaques enduites de 0,25 mm de gel de silice 60. Le compose jaune avec un R/de 0,27 a alors ete recupere de la silice en utilisant un melange de chloroforme : methanol (7:3). La TA a ete quantified par chromatographie liquide de haute performance (HPLC) en utilisant un chromatographe liquide Varian LC5500 equipe d'une colonne C18 (10 (am, 3,9 X 300 mm).
1.2 CULTURE DES CELLULES DE PEUPLIER
Les cellules de peuplier hybride (Populus trichocarpa X P. delto'ides HI 1-11) ont ete cultivees dans un milieu Murashige et Skoog (MS, pH 5,7), additionne de vitamines de
Gamborg (Moniz de Sa et al., 1992). Les cultures cellulaires ont ete maintenues sur un
agitateur rotatif a une vitesse de 120 rpm et a l'obscurite. Les cellules ont ete repiquees en procedant a une sous-culture, soit 1 volume de cellules a 3 volumes de milieu frais. Cette sous-culture a ete realisee chaque semaine dans le cas des cellules non-habituees ou a chaque deux semaines pour les cellules habituees a la TA.
1.3 HABITUATION DES CELLULES DE PEUPLIER A LA TA
L'adaptation des cellules de peuplier a 1,3 uM de TA a ete realisee precedemment par Brochu (2005) en ajoutant a chaque sous-culture une concentration croissante de TA pendant une periode d'un an, en commencant avec 0,1 uM de TA. De 2 a 4 sous-cultures ont ete realisees avec la meme concentration de TA avant d'en augmenter la dose. Les cellules non-habituees ont recu le meme volume en methanol que les cellules non-habituees a la TA. A partir d'une concentration de 0,4 uM de TA, les cellules en cours d'habituation a la TA avaient une croissance reduite. Par la suite, ces cellules ont ete transferees aux 14 jours. Apres une periode de 12 mois, les cellules etaient habituees a 1,3 uM de TA. Depuis 4 ans, ces cellules sont cultivees en 1'absence de TA.
1.4 TRAITEMENTS DES CELLULES
La TA, 1TSOX (Crescent Chemicals Co.) et le DCB (Sigma-Aldrich Co.) ont ete dissous dans du methanol et ajoutes aux suspensions cellulaires aux concentrations finales indiquees. Le meme volume de methanol a ete ajoute aux cellules temoins des differents traitements avec les inhibiteurs. Les traitements ont eu lieu 3 jours apres le transfert, lorsque les cellules etaient dans leur phase exponentielle de croissance. Lors du traitement a la lumiere, les suspensions cellulaires ont ete soumises a un cycle de 12 heures de lumiere a 100 uE m" s" , pendant 7 jours.
1.5 DETECTION DE LA MORTALITE CELLULAIRE
Le bleu de trypan a ete utilise comme colorant vital. Une part de cellules en suspension a ete melangee a une part de trypan bleu 0,4 % (Sigma) dilue dans 140 mM NaCl et 3,5 mM K2HPO4. Trois repetitions ont ete effectuees par condition et au moins 500 cellules ont ete comptees par echantillon. Le comptage des cellules a ete realise en les observant a l'aide d'un microscope optique DMLS (Leica).
1.6 TEST TUNEL
La degradation de l'ADN a ete detectee en utilisant la trousse de detection de la mort cellulaire in situ a la fiuoresceine (Roche) en suivant les recommandations du fabricant. Les differents types de cellules ont d'abord ete fixes pendant 45 minutes avec du paraformaldehyde 4 % dilue dans du PBS, suivi de deux lavages au PBS. Avant d'appliquer la solution de marquage des extremites 3'OH libres, les cellules fixees ont ete traitees avec de la proteinase K (20 jj,g ml"1, Sigma) pendant 15 minutes. Les noyaux ont ete colores avec du
DAPI(5 ugml"1, Sigma).
1.7 MESURE DES DIMENSIONS CELLULAIRES
Les photos de cellules, prealablement colorees au bleu de trypan, ont ete prises en microscopie optique a l'aide d'un microscope Axiolmager Zl (Zeiss). Le pourtour des cellules vivantes a ete delimite a l'aide d'un curseur. Le logiciel AxioVision LE V4.5
associe a ce microscope genere automatiquement les differentes mesures en um, soit la longueur, la largeur, le perimetre et l'aire.
1.8 COLORATION DES PECTINES
Le rouge de ruthenium a ete utilise comme colorant pour la visualisation des pectines. Une part de cellules en suspension a ete melangee a une part de rouge de ruthenium 0,02 %.
1.9 EXTRACTION ET QUANTIFICATION DES PECTINES DE LA PAROI
1.9.1 Extraction des parois cellulaires
Les cellules de peuplier agees de 5 jours apres la sous-culture ont ete filtrees, puis congelees rapidement dans l'azote liquide et conservees a -80 °C. Les parois cellulaires ont ete extraites a l'aide de la methode d'Encina et al. (2002): les cellules ont ete lavees avec du tampon phosphate de potassium, digerees avec de l'a-amylase durant 4 heures (Sigma-Aldrich Co.), lavees sequentiellement avec du tampon phosphate de potassium, de l'eau, de Pacetone, du methanol-chloroforme et du diethylether et sechees a Fair libre.