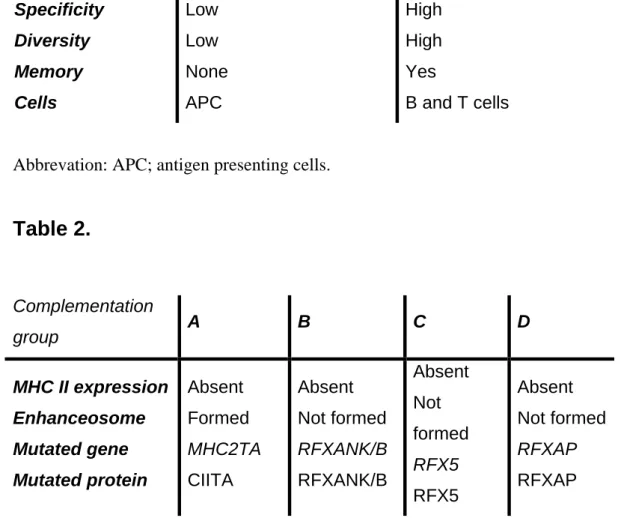
Expression of MHC II genes
Texte intégral
Figure
Documents relatifs
We searched binding sites of miRNAs in mRNAs of 157 transcription factors' genes of wild camel (Camelus ferus).. The mRNAs of 96 genes of zincfinger transcription factors' family
Keywords: Major histocompatibility complex, Family Equidae, MHC exon 2, MHC class II loci, Positive selection, Trans-species polymorphism, Selected amino acid sites.. ©
Under normal circum- stances, NK-cell activation is counteracted by inhibitory NK-cell receptors, which interact with MHC-I (major histocompatibility complex class I) molecules
To distinguish between the roles of enhanceosome binding, CIITA recruitment, PIC assembly and active transcription in promoting nucleosome eviction at the HLA-DRA gene we
effect for a 26-bp deletion in the RFXANK gene in North African major histocompatibility complex class II-deficient patients belonging to complementation group B. Wiszniewski
To investigate a dependence of Bye1 recruitment to chromatin on active Pol II transcription, we followed Bye1 occupancy at promoters and coding regions of class II genes by ChIP in
Bioinformatic prediction is a method to screen for potentially relevant T-cell epitopes based on binding to major histocompatibility complex (MHC)-I or MHC-II. Selected peptides
Increased Genome-Wide Recruitment of Maf1 to Class III Genes under Repressing Conditions The Maf1-mediated modification of the occupancy of class III genes by the Pol III
