difusion1
Texte intégral
Figure
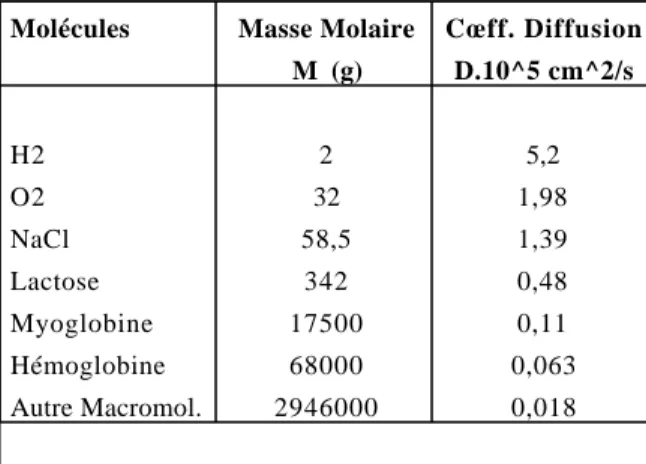
Documents relatifs
Ceci justifie l’affirmation : « On détermine le TAC si le pH d'une eau est inférieur à 8,2 car dans ce cas, l'eau contient pratiquement et uniquement des ions et ne
Sodium 136 mmol/l Potassium 5,3 mmol/l Bicarbonates 18 mmol/l Glucose 5,2 mmol/l Protéines 66 g/l Urée 34 mmol/l Créatinine 750 µmol/l Calcium 2,05 mmol/l Phosphates 1,9
Il contient des protéines très utiles en cas d’hémorragies, pour les grands brûlés, les hémophiles… Le don est réalisé soit lors d’un don de sang total (les
Il contient des protéines très utiles en cas d’hémorragies, pour les grands brûlés, les hémophiles… Le don est réalisé soit lors d’un don de sang total (les
INGRÉDIENTS : eau, sucre, sirop de glucose, rhum, farine de blé , préparation alimentaire (poudre de lactosérum, matière grasse végétale (coprah), protéines de lait , poudre de
Thème du programme : Les molécules organiques dans le domaine de la santé Savoir quels sont les principaux constituants du lait : eau, glucides, lipides,.. protéines, vitamines,
Thème du programme : Les molécules organiques dans le domaine de la santé Savoir quels sont les principaux constituants du lait : eau, glucides, lipides,.. protéines, vitamines,
D’où viennent les réactifs chez les hétérotrophes ?.. Eau, CO2, O2, glucose, sels minéraux, glucides, protéines, lipides, sève brute, sève élaborée, photosynthèse,
