Cytosolic Proteins Regulate
␣-Synuclein Dissociation from
Presynaptic Membranes
*
Received for publication, June 22, 2006, and in revised form, August 3, 2006 Published, JBC Papers in Press, August 22, 2006, DOI 10.1074/jbc.M605965200
Sabine Wislet-Gendebien1, Cheryl D’Souza, Toshitaka Kawarai, Peter St George-Hyslop, David Westaway, Paul Fraser, and Anurag Tandon2
From the Centre for Research in Neurodegenerative Diseases, University of Toronto, Toronto, Ontario M5S 3H2, Canada Intracellular accumulation of insoluble␣-synuclein in Lewy
bodies is a key neuropathological trait of Parkinson disease (PD). Neither the normal function of␣-synuclein nor the bio-chemical mechanisms that cause its deposition are understood, although both are likely influenced by the interaction of
␣-synuclein with vesicular membranes, either for a physiologi-cal role in vesicular trafficking or as a pathologiphysiologi-cal seeding mechanism that exacerbates the propensity of␣-synuclein to self-assemble into fibrils. In addition to the␣-helical form that is peripherally-attached to vesicles, a substantial portion of
␣-synuclein is freely diffusible in the cytoplasm. The mecha-nisms controlling ␣-synuclein exchange between these com-partments are unknown and the possibility that chronic dys-regulation of membrane-bound and soluble␣-synuclein pools may contribute to Lewy body pathology led us to search for cel-lular factors that can regulate␣-synuclein membrane interac-tions. Here we reveal that dissociation of membrane-bound
␣-synuclein is dependent on brain-specific cytosolic proteins and insensitive to calcium or metabolic energy. Two PD-linked mutations (A30P and A53T) significantly increase the cytosol-dependent␣-synuclein off-rate but have no effect on cytosol-independent dissociation. These results reveal a novel mecha-nism by which cytosolic brain proteins modulate␣-synuclein interactions with intracellular membranes. Importantly, our finding that␣-synuclein dissociation is up-regulated by both familial PD mutations implicates cytosolic cofactors in disease pathogenesis and as molecular targets to influence␣-synuclein aggregation.
␣-Synuclein (␣-syn)3is a member of a multigene synuclein
family that is highly abundant in presynaptic terminals of
mam-malian brain (1, 2). The function of␣-syn is poorly resolved, although it is attributed with wide ranging roles in vesicular trafficking and vesicle biogenesis and as a molecular chaperone (3).␣-Syn is also implicated in a broad spectrum of neurode-generative disorders collectively named synucleinopathies, being a primary component of Lewy bodies (4), and as a frag-ment in Alzheimer disease plaques (5). In a small number of pedigrees, autosomal-dominant inheritance of Parkinson dis-ease (PD) is linked to either multiplication of the normal␣-syn gene or to one of three missense mutations (A30P, E46K, and A53T) (6, 7).
In vitro studies suggest that ␣-syn is natively unfolded in aqueous solution, and exposure to lipids stabilizes the amino terminus in an amphipathic␣-helix that aligns polar and non-polar residues into opposing orientations (8 –11). Presumably, this secondary structure confers the lipid-binding properties for direct membrane interaction such that purified recombi-nant␣-syn can bind to small diameter artificial vesicles rich in acidic phospholipids (8, 12), to purified synaptic vesicles (13, 14), and to membranes within intact cells (15). Most studies examining␣-syn membrane binding have used either artificial phospholipids or purified membranes, without consideration to the potential regulatory function of soluble or membrane factors that may be pertinent to disease progression. Previous
investigations were equivocal on whether ␣-syn mutations
(A30P and A53T) affect membrane interactions; the A30P or A53T mutation had little or no effect on␣-syn binding (16, 17), the A53T mutation reduced (8) or increased (18, 19) the mem-brane binding, or the A30P had reduced binding to purified synaptic vesicles (14, 20, 21). Further complexity may also arise from reversible membrane interaction, akin to that of cytosolic regulators of vesicular trafficking whose direct or indirect asso-ciation with vesicle membranes is necessarily transient (e.g. exocytosis: NSF,␣/-SNAP, rab3, rabphilin, synapsin, CAPS; endocytosis: clathrin/adaptor, dynamin) (22–24). Association and dissociation is coupled to the vesicle life cycle, dictated by cation fluxes, nucleotide triphosphate cleavage, and post-trans-lational modifications. In the case of rab3, an additional mech-anism governs its membrane attachment. Another cytosolic protein, GDP-dissociation inhibitor, is essential to extract rab3 off exocytosing vesicles and deliver it to newly synthesized ves-icles (25–27).
It is unknown whether␣-synuclein exchange occurs between
soluble and membrane compartments or even whether␣-syn
can dissociate from reconstituted or biological membranes. The possibility that chronic dysregulation of subcellular ␣-synuclein pools may contribute to PD and other
synucle-*This work was supported by operating grants (to A. T.) from the Canadian
Institute of Health Research (CIHR) and the Parkinson Society of Canada. The costs of publication of this article were defrayed in part by the pay-ment of page charges. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. Section 1734 solely to indi-cate this fact.
1Postdoctoral support was provided by the Leon Frederick Foundation and
the Journal of Cell Science travel grant.
2To whom correspondence should be addressed: Centre for Research in
Neu-rodegenerative Diseases, University of Toronto, 6 Queen’s Park Crescent West, Toronto, Ontario M5S 3H2, Canada. Tel.: 8880; Fax: 416-978-1878; E-mail: [email protected].
3The abbreviations used are:␣-syn, ␣-synuclein; PD, Parkinson disease; Wt,
wild-type; ANOVA, analysis of variance; Tg, transgenic; ATP␥S, adenosine
5⬘-O-(thiotriphosphate); GTP␥S, guanosine 5⬘-3-O-(thio)triphosphate;
AMP-PNP, adenosine 5⬘-(,␥-imino)triphosphate; AMP-CP, ␣,-methylene
adenosine diphosphate.
at UNIV DE LIEGE-MME F PASLE on December 1, 2008
www.jbc.org
inopathies prompted our search for cellular factors that affect ␣-synuclein membrane interactions. To address this, we devel-oped an assay comprised of permeabilized synaptosomes and
report here that ␣-syn dissociation from presynaptic
mem-branes requires cytosolic proteins from the brain. Particularly relevant to disease pathogenesis was the finding that cytosol-dependent␣-syn dissociation was significantly increased by two PD-related mutations (A30P and A53T) suggesting a mechanism whereby cytosolic cofactors can regulate␣-syn sol-ubility and, thereby, modulate disease progression.
EXPERIMENTAL PROCEDURES
Transgene Construction and Generation of Transgenic Mice— The human␣-syn gene containing a eukaryotic Kozak initiation sequence (GCCGCCACC) (28, 29) upstream of the start codon was ligated into a linearized cos-Tet expression vector contain-ing the Syrian hamster prion protein promoter gene (30, 31). A30P and A53T␣-syn mutant transgenes were generated using
the Quick change kit (Stratagene). Following microinjection into fertil-ized oocytes of FVB/N mice, posi-tive founders were identified by amplification of genomic DNA and bred to FVB wild-type mice. To gen-erate␣-syn Tg mice lacking
endog-enous murine␣-syn, the ␣-syn Tg
mice were crossed with SNCA⫺/⫺
mice (Jackson Laboratories). F1 off-spring positive for␣-syn transgene were backcrossed and F2 progeny were selected for the presence of the neomycin resistance gene and the
absence of the endogenous ␣-syn
gene as described previously (32). All animal experiments were performed according to guidelines established in the Canadian Guide for the Care and Use of Laboratory Animals.
Synaptosome Preparation —Syn-aptosomes were prepared as de-scribed (26, 33). Briefly, brains from mice (A30P, A53T, and Wt trans-genic mice) were dissected and homogenized with 10 strokes at 500 rpm, in ice-cold buffer A (320 mM
sucrose, 1 mM EGTA, and 5 mM
HEPES (pH 7.4) and centrifuged at 3000⫻ g for 10 min. The superna-tant was spun for 10 min at 14,000⫻
gand the pellet (P2) resuspended in buffer A. The P2 fraction was loaded onto a discontinuous Ficoll gradient (13, 9, and 5% in buffer A) and cen-trifuged for 35 min at 35,000⫻ g. The 13–9% interface, containing intact synaptosomes, was resus-pended in buffer B (140 mMNaCl, 5
mM KCl, 20 mM HEPES, 5 mM
NaHCO3, 1.2 mMNa2HPO4, 1 mMMgCl2, 1 mMEGTA, and 10
mMglucose) and spun at 14,000⫻ g for 10 min. The pellet was hypotonically lysed by two washes in buffer C (10 mMHEPES,
18 mMKOAc (pH 7.2)), spun at 14,000⫻ g for 10 min, and
resuspended in buffer D (25 mMHEPES, 125 mMKOAc, and 2.5 mMMgCl2). After centrifugation (14,000⫻ g for 10 min), syn-aptosomes were resuspended in buffer D and incubated (as indicated in each figure legend) with or without␣-syn deficient brain or liver cytosol before separating membrane and super-natant by centrifugation at 14,000⫻ g for 10 min. The super-natant was centrifuged at 100,000⫻ g to remove trace contam-inating elements such as broken membranes or dissociated synaptic vesicles before quantifying soluble␣-syn by Western blotting.
Cytosol Preparation from␣-Syn Knock-out and Non-trans-genic Mice—Mouse brains were homogenized in 10 volumes of
homogenizing buffer (1 mM sucrose, 1 M KOAc, 100 mM
MgOAc, 1MHEPES (pH 7.4)). The homogenate was spun at
FIGURE 1. A,␣-syn dissociation assay. Step 1, synaptosomes and cytosol were prepared separately from brains
of Tg mice expressing the human␣-syn (Wt, A30P, or A53T) and ␣-syn-deficient mice, respectively. Step 2,
following hypotonic lysis of synaptosomes, the washed membranes (␣-syn donor) were co-incubated with
␣-syn⫺/⫺cytosol (␣-syn acceptor). Step 3, membrane and cytosolic fractions were separated by centrifugation
and the supernatants were analyzed by Western blotting. B, distribution of␣-syn in acceptor and donor
fractions. Western blots showing total synaptosomal␣-syn (detected with anti-␣-syn antibody Syn-1),
glycer-aldehyde-3-phosphate dehydrogenase (GAPDH), and synaptophysin recovered in either supernatant (S) or pellet (P) following resuspension of intact synaptosomes in isotonic buffer, hypotonic buffer, or hypotonically lysed synaptosomes (Donor fraction) washed three times and resuspended in isotonic buffer for 10, 20, or 30
min. As expected, no␣-syn was detected in brain cytosol from ␣-syn-deficient mice (acceptor fraction:
knock-out cytosol).
at UNIV DE LIEGE-MME F PASLE on December 1, 2008
www.jbc.org
15,000⫻ g for 10 min and the supernatant was then centrifuged at 100,000⫻ g for 1 h. The supernatant was dialyzed 4 times with cold dialysis buffer (1 mMKOAc, 1MHEPES (pH 7.2)) for 45 min each, centrifuged at 28,000⫻ g for 25 min, and stored at ⫺80 °C. Protein concentration was determined by BCA protein assay (Pierce).
Western Blotting—Proteins were boiled briefly in loading buffer (10% v/v glycerol; 0.05MTris (pH 6.8), 2% SDS, brom-phenol blue, and 2.5% v/v-mercaptoethanol) and separated by electrophoresis using 12% Tris-glycine polyacrylamide gels. Proteins were transferred to nitrocellulose (Life Sciences) and probed by Western blotting using: antibodies against ␣-syn (monoclonals 211 and Syn-1, Neomarkers), our own rabbit polyclonal (LWS1, 1:1000) raised to a 24-mer␣-syn-specific peptide, glyceraldehyde-3-phosphate dehydrogenase
(mono-clonal 6C5, Biodesign), or
synapto-physin (monoclonal, Biodesign).
Bound horseradish peroxidase-con-jugated anti-mouse or anti-rabbit IgG (Sigma) were revealed by chemilumi-nescence using ECL Plus (GE Health-care) and quantified with a Storm 860 fluorescent imager and ImageQuant software (GE Healthcare). Statistical comparisons were done with Graph-Pad InStat software using Student’s t test for comparisons between two groups or ANOVA (Bonferroni’s test) for multiple comparisons.
RESULTS
Cytosolic Proteins Increase␣-Syn Dissociation from Membrane—To identify regulators of␣-syn solubil-ity, we assessed the effects of cytoso-lic proteins on the interaction of ␣-syn with membranes. We meas-ured␣-syn dissociation from synap-tosomal membranes prepared from brains of transgenic mice expressing
human␣-syn as the donor of
mem-brane-bound␣-syn in the absence
or presence of brain cytosol derived
from ␣-syn-deficient mice as the
acceptor soluble fraction (Fig. 1A).
Intact synaptosomes from ␣-syn
transgenic mouse brain suspended in hypotonic buffer release⬃30% of
their total ␣-syn content, which
comprises the soluble cytoplasmic pool (Fig. 1B). The remaining␣-syn is stably associated with the mem-brane fraction following resuspension in isotonic buffer and is entirely sus-ceptible to membrane disruption by detergents. The release of mem-brane-derived␣-syn into the super-natant was markedly increased by the addition of cytosol, whereas integral membrane proteins, such as synaptophysin, were retained by the membrane pellet (Fig. 2A). Preincubation of membranes with antiserum raised to ␣-syn inhibited ␣-syn dissociation, in contrast to anti-synapto-physin antibodies or preimmune serum, neither of which had any effect (Fig. 2B). The ability to dissociate from presynaptic membranes was not exclusive to human␣-syn, because endog-enous␣-syn from non-transgenic mice also exhibited similar cytosol-dependent dissociation from presynaptic membranes (Fig. 2C).
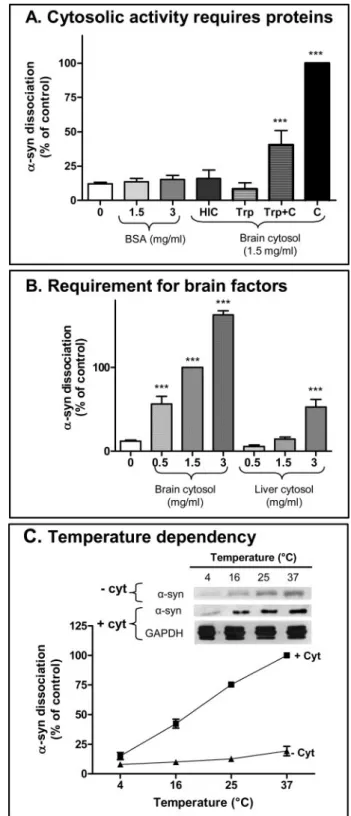
Comparable levels of a nonspecific protein, bovine serum albumin (BSA), did not affect␣-syn solubility (Fig. 3A). How-ever, the cytosolic activity was heat- and protease-labile as cytosol preincubated at 95 °C or pre-digested with trypsin failed to induce ␣-syn dissociation, suggesting that this process
FIGURE 2. A, cytosol-dependent␣-syn dissociation from synaptosome membranes. ␣-Syn Tg membranes and
brain␣-syn-deficient cytosol were co-incubated to assess the role of cytosolic proteins on ␣-syn dissociation.
Incubation for 10 min at 37 °C of hypotonically lysed synaptosome membranes (100 mg of wet synaptosomal membrane weight) with various cytosol concentrations (0 –1.5 mg/ml, total protein concentration) increased
soluble human␣-syn, as detected by human-specific antibody 211. The integral membrane protein
synapto-physin was not released into the supernatant but remained associated with the membrane pellet (Membr) solubilized with 1% Triton X-100 buffer. B, quantification of cytosol concentration on dissociation of human ␣-syn. Mean ␣-syn intensity and S.E. of four independent experiments and normalized to ␣-syn solubilized by
1.5 mg/ml cytosol.␣-Syn dissociation in the presence of 1.5 mg/ml cytosol was blocked by pretreatment of
membranes with LWS1␣-syn antibody (Student’s t test, p ⬍ 0.0001, n ⫽ 4, compared with 1.5 mg/ml cytosol
without antibody condition), but not anti-synaptophysin antibody (Student’s t test, p⬎ 0.3, n ⫽ 4) or
preim-mune serum (Student’s t test, p⬎ 0.7, n ⫽ 4). C, regulated dissociation of murine␣-syn. Western blot probed
with Syn-1 antibody showing endogenous murine␣-syn from non-Tg mice also exhibited cytosol-dependent
dissociation from synaptosome membranes upon incubation for 10 min at 37 °C with 1.5 mg/ml␣-syn⫺/⫺
cytosol.
at UNIV DE LIEGE-MME F PASLE on December 1, 2008
www.jbc.org
requires specific cytosolic proteins (Fig. 3A). To verify that pro-teolytic activity was completely inactivated by trypsin inhibitor before exposure to membranes, we showed that preincubation of trypsinized-cytosol with an equal concentration of untreated cytosol partially rescued␣-syn dissociation activity. Moreover, equivalent concentrations of liver-derived cytosol were sig-nificantly less active compared with brain cytosol, suggest-ing that the factors inducsuggest-ing␣-syn-dissociation are brain-specific (Fig. 3B). The cytosol-dependent component of ␣-syn dissociation from synaptic membranes was responsive to temperature, increasing⬃10-fold from 4 to 37 °C. In con-trast, cytosol-independent ␣-syn dissociation was insensi-tive over this range (Fig. 3C).
Cytosolic Proteins Are Rate-limiting for␣-Syn Translocation—
The kinetics of␣-syn dissociation from membranes are shown in Fig. 4A. Both the rate and the extent of␣-syn dissociation were increased by raising the cytosol concentration. The appearance of soluble␣-syn reached a plateau within 5–10 min, suggesting that cytosolic factors are rate-limiting under the conditions of our assay. Because the liberated␣-syn represents
less than 5% of the total membrane-associated ␣-syn, the
majority of the␣-syn is unaffected. Therefore, we examined whether any of the remaining bound␣-syn was susceptible to cytosol-mediated dissociation by subsequent exposure to cytosol. Fig. 4B shows that additional membrane-bound␣-syn could indeed be solubilized, although the amount in successive incubations declined somewhat, consistent with a reduction in the pool of extractable␣-syn. If cytosol was omitted during the second incubation, no additional␣-syn release was detected arguing that each dissociation event requires cytosolic factors and that␣-syn is not primed for dissociation by the prior expo-sure to cytosol.
We tested whether cytosol used during one incubation retained activity to extract additional␣-syn from fresh synaptic membranes. To distinguish between␣-syn extracted during the first and second incubation, we used synaptosomes from non-Tg mice in the first incubation and measured the
dissoci-ation of endogenous murine␣-syn using the syn-1 antibody
(Fig. 4C, lane 1). The “used” cytosol, now containing murine ␣-syn, was incubated with permeabilized synaptosomes from
human␣-syn Tg mice that are murine ␣-syn-deficient. The
anti-␣-syn antibody 211, which is specific for the human
iso-form, did not reveal any human␣-syn released from
mem-branes in the presence of used cytosol (Fig. 4C, lane 2) but did so when using fresh cytosol (Fig. 4C, lane 3). These results suggest that the availability of cytosolic proteins determines the extent
FIGURE 3. Cytosolic brain proteins regulate␣-syn dissociation from
membranes. A, cytosolic activity is heat and protease labile. Increasing
non-specific protein concentration with bovine serum albumin (BSA) did not
induce additional␣-syn dissociation compared the “no cytosol” treatment (0
mg/ml cytosol) (one-way ANOVA, p⬎ 0.9, n ⫽ 4). Pre-exposure of brain
cytosol to either heating for 3 min at 95 °C (heat-inactivated cytosol, HIC) or to trypsin (Trp) digestion completely eliminated the ability of 1.5 mg/ml cytosol
(C ) to induce␣-syn dissociation (one-way ANOVA, p ⬎ 0.8, n ⫽ 4–8). For the
trypsin experiments, proteolytic activity was terminated with trypsin inhibi-tor prior to the incubation with membranes, as shown by the partial rescue
after half-dilution with untreated brain cytosol (Trp⫹C). ***, p ⬍ 0.01 relative
to no cytosol condition (one way ANOVA, n⫽ 4; Bonferroni’s multiple
com-parison test). B,␣-syn dissociation activity is concentrated in brain cytosol.
The activities of brain and liver cytosol were compared. Liver-derived cytosol showed some weak activity only at the highest concentration tested, whereas
brain cytosol induced significant␣-syn dissociation at 6-fold lower
concen-tration 0.5 mg/ml (***, p⬍ 0.0001, one-way ANOVA, n ⫽ 4; Bonferroni’s
mul-tiple comparison test as compared with 0 mg/ml cytosol condition). C,
cytosol-dependent␣-syn dissociation is influenced by temperature. Top panel,
Western blot showing␣-syn dissociation in presence or absence of cytosol
during 10-min incubation at various temperatures (4, 16, 25, and 37 °C).
Bot-tom panel, mean␣-syn intensity and S.E. from five independent experiments
and normalized to␣-syn dissociation with 1.5 mg/ml cytosol (one-way
ANOVA, p⬍ 0.0001, n ⫽ 5; Bonferroni’s multiple comparison test revealed
that temperature significantly increased cytosol dependent␣-syn
dissocia-tion). No significant correlation was observed with temperature change in
absence of cytosol (one way ANOVA, p⬎ 0.5, n ⫽ 5). GAPDH,
glyceraldehyde-3-phosphate dehydrogenase.
at UNIV DE LIEGE-MME F PASLE on December 1, 2008
www.jbc.org
of␣-syn dissociation and that they do not regenerate under our experimental conditions.
␣-Syn Dissociation Is Not Regulated by Ca2⫹or by Nucleotide Triphosphates—Because many nerve terminal functions are subject to regulation by Ca2⫹influx and the availability of
met-abolic energy, we examined whether ␣-syn translocation is
affected by changes in these parameters. No significant differ-ences on␣-syn dissociation were observed in the presence or
absence (with EGTA) of 1 mMCa2⫹and in the presence of
increasing concentrations of cytosol, suggesting a Ca2⫹ -inde-pendent mechanism (Fig. 5A). Next, we assessed the effects of ATP and GTP, their non-hydrolyzable analogs, and their vari-ous metabolic products in our assay. We did not observe any significant changes to␣-syn dissociation in the presence of any of these compounds (Fig. 5B), suggesting that␣-syn dissocia-tion occurs independently of high energy phosphates.
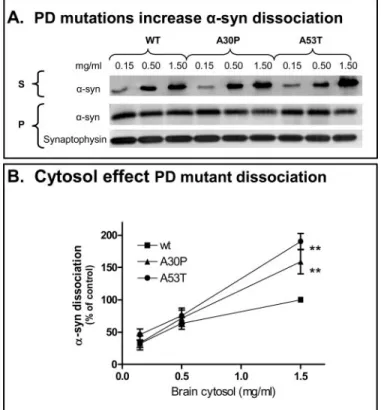
Parkinson Disease Mutations Increase Cytosol-induced␣-Syn Translocation—We analyzed whether the PD-associated muta-tions, A30P and A53T, affect␣-syn dissociation using equiva-lent amounts of synaptosomal membrane (100 mg of wet syn-aptosomal membrane weight). Fig. 6 shows that, as with Wt ␣-syn, cytosol exposure also increased the dissociation of
mutant␣-syn from synaptic membranes. However, the
disso-ciation of both A30P and A53T mutants, which was clearly cytosol-dependent, was significantly greater than that of Wt ␣-syn at the higher cytosol concentration (p ⬍ 0.01). To exclude the possibility that differences in dissociation rates of ␣-syn mutants are due to variations in expression level or sub-cellular compartmentalization, we determined the distribution of␣-syn in synaptosomes. We first compared ␣-syn expression in the cytosolic pool (released by hypotonic lysis) relative to the total synaptosomal␣-syn (solubilized by detergent). Regardless of the transgene,⬃30% of␣-syn in isolated nerve terminals is soluble (Fig. 7A); the expression levels of Wt and mutant human␣-syn were indistinguishable. The hypotonically lysed synaptosomes, which represent the␣-syn donor in our dissoci-ation assay, were centrifuged and␣-syn in the pellets measured. Again, there were no significant differences between the mem-brane fractions of Wt and mutants␣-syn (Fig. 7B). In effect, this result indicates that the clinical mutations do not inherently destabilize the␣-syn interaction with biologically derived par-ticulate fraction, since association was maintained with low
cytosol concentrations. Rather, the removal of ␣-syn from
membranes is a mediated event that is enhanced by the two missense mutations.
DISCUSSION
Current models of␣-syn lipid interaction predict that the ami-no-terminal portion of␣-synassemblesintoanamphipathic␣-he-lix that embeds partially into membrane bilayers (8, 10, 11, 16, 34). In accord with this concept, our results show that the majority of
synaptosomal ␣-syn is associated with membranes in vivo,
although substantial portion is freely diffusible within the cyto-plasm. How these bound and diffusible pools are maintained
and whether regulated exchange occurs between these␣-syn
compartments is unclear. Here, we present evidence that␣-syn stably associated with the membrane fraction can be recruited into the soluble fraction in the presence of brain cytosol. This was not due to a detergent action of cytosol because integral membrane proteins, e.g. synaptophysin, remained with the membrane pellet and the process was blocked specifically by antibodies to␣-syn. Because the antiserum alone had no affect on␣-syn distribution in the absence of cytosol, the antibodies likely prevent access of cytosolic factors to membrane␣-syn. Moreover, predigestion of brain cytosol with trypsin or pre-heating at 95 °C eliminated the activity, directly implicating a
FIGURE 4. Cytosolic activity is rate-limiting for␣-syn dissociation. A, ␣-syn
dissociation kinetics are regulated by cytosol concentration. Top panel,
rep-resentative Western blot showing the time-course of␣-syn dissociation in the
absence or presence of 1.5 mg/ml brain cytosol. The extent of␣-syn
dissoci-ation reached a plateau at within 5–10 min in presence of different
concen-trations of cytosol. Bottom panel, quantification of␣-syn dissociation kinetics
(mean and S.E.) from four to five independent experiments each. B,
mem-brane␣-syn is susceptible to multiple extractions with cytosol. Repeated
co-incubation of the hypotonically lysed synaptosomes with fresh cytosol (⫹) for
10 min at 37 °C revealed a substantial extractable pool of membrane-bound ␣-syn. There was a slight, but statistically insignificant, decrease in ␣-syn
dis-sociation with each incubation (one-way ANOVA, p⬎ 0.2, n ⫽ 4). No␣-syn
was released from membranes if fresh cytosol was omitted (⫺), indicating the
cytosolic factors must be present to induce␣-syn dissociation. C, cytosolic
activity is depleted by single exposure to synaptic membranes. KO cytosol was first incubated with non-Tg membranes to induce dissociation of murine ␣-syn as detected by monoclonal antibody Syn-1 in lane 1. The same cytosol was subsequently incubated with synaptic membranes derived from human ␣-syn Tg mice that lack endogenous murine ␣-syn. Supernatant from the
second incubation in lane 2 contained only the murine␣-syn from the first
incubation, but no human Tg␣-syn immunoreactivity was detected by the
human specific␣-syn monoclonal antibody 211. In contrast, fresh cytosol
induced dissociation of human␣-syn from the Tg membranes in lane 3.
GAPDH, glyceraldehyde-3-phosphate dehydrogenase.
at UNIV DE LIEGE-MME F PASLE on December 1, 2008
www.jbc.org
role for specific cytosolic proteins in controlling␣-syn solubil-ity. The permissive factors appear to be enriched in brain since 6-fold greater level of liver cytosol was required to achieve equivalent␣-syn dissociation as brain cytosol.
The proteins triggering ␣-syn dissociation are in limiting quantity in cytosol and are not regenerated under the condi-tions of our assay. A single exposure to synaptosome mem-branes was sufficient to deplete cytosolic capacity to extract
membrane␣-syn so that subsequent incubations with fresh
membranes yielded no additional soluble␣-syn. In contrast,
presynaptic membranes retained ample extractable ␣-syn,
which could be dissociated with subsequent applications of cytosol. While this suggests that the rate and extent of␣-syn dissociation is primarily dependent on the availability of cyto-solic factors, additional studies will be needed to determine
whether some of the␣-syn that remains membrane-bound is
biochemically distinct from the cytosol-extracted␣-syn. The cytosolic proteins in our experiments are substantially less concentrated than presynaptic cytosol in vivo (estimated at 200 –300 mg/ml)4as a result of dilution with extraction buffers
during isolation and contamination from non-neuronal cells in brain. Arguably, the portion of␣-syn that can potentially be mobilized from membranes is likely to be correspondingly greater in intact cells than the 5% shifts we detected. This is compatible with a previous report by Fortin et al. (35) showing that GFP-tagged␣-syn expressed in primary hippocampal neu-rons accumulates at synaptic boutons and that the fluorescence at photobleached boutons recovers rapidly as if␣-syn has sub-stantial steady state mobility and can be replenished from adja-cent boutons. Neuronal stimulation in the presence of extracel-lular Ca2⫹dispersed the␣-syn fluorescence from presynapses in a tetanus toxin-sensitive manner, suggesting that ␣-syn translocation occurred after exocytosis and that Ca2⫹entry per
seis an insufficient trigger. We also did not observe any Ca2⫹ dependence or a direct requirement for nucleotide triphos-phate cleavage in inducing␣-syn dissociation. Although it is likely that post-exocytotic regulatory aspects are not recapitu-lated in our assay, our observations demonstrating cytosol-me-diated␣-syn dissociation may underlie the movement of GFP-␣-syn during resting conditions or associated with exocytosis (35). Our results suggest a dynamic model of␣-syn interaction
4W. Balch, personal communication.
FIGURE 5. Calcium and phosphate high energy did not affect␣-syn dissociation. A, histogram shows mean and S.E. of ␣-syn dissociated from synaptic
membranes in the presence different concentration of brain cytosol supplemented with 1 mMCa2⫹or 1 mMEGTA. No significant difference was observed
between EGTA and Ca2⫹treatments (one-way ANOVA, p⬎ 0.5, n ⫽ 5). B, the effects of 1 mMadenosine, ATP, ADP, GTP, ATP␥S, and GTP␥S, or 3 mMof AMP-CP
(non-hydrolysable ADP analog) and AMP-PNP (the non-hydrolysable ATP analog) on␣-syn dissociation were measured in the presence or absence of cytosol
following incubation at 37 °C for 10 min. Neither nucleotide triphosphate nor their non-hydrolyzable analogs or metabolic products affected significantly basal
or cytosol-induced␣-syn dissociation (one-way ANOVA, with cytosol p ⬎ 0.1, n ⫽ 5; without cytosol p ⬎ 0.2, n ⫽ 5). CTRL, control.
FIGURE 6. Increased dissociation of A30P and A53T␣-syn compared with
Wt␣-syn. A, two familial PD-linked mutations increase ␣-syn dissociation.
Dissociation of␣-syn from equivalent amounts of synaptic membranes
(100 mg of wet synaptosomal membrane weight) derived from Wt, A53T,
or A30P transgenic␣-syn mice was measured in the presence of various
concentrations of cytosol (0.15, 0.5, and 1.5 mg/ml). The top panel shows a
representative Western blot of␣-syn in the supernatant (S) and in the
membrane pellet (P) after a 10-min incubation at 37 °C (using human
spe-cific anti-␣-syn antibody 211). Synaptophysin immunoreactivity reflects
synaptic membrane input. The bottom panel illustrates the combined
results of four independent experiments. Dissociation of␣-syn with either
PD-linked mutation was significantly higher in the presence of 1.5 mg/ml
cytosol (***, one-way ANOVA, Bonferroni’s test, p⫽ 0.0012) but not in the
presence of lower cytosol concentrations (0.15 and 0.5 mg/ml cytosol;
one-way ANOVA, p⬎ 0.7).
at UNIV DE LIEGE-MME F PASLE on December 1, 2008
www.jbc.org
with membranes, in which cytosolic proteins control the exchange of membrane-stabilized␣-syn with a diffusible form, and may explain some discrepancies in previous studies regard-ing the proportions of soluble and bound␣-syn, which ranged from equal distribution (36, 37) to virtually all soluble (13).
The fact that most of the␣-syn in synaptosomes is
mem-brane-bound during steady state conditions suggests that net cytosol-induced dissociation is impeded until activation by physiological stimuli and that the equilibrium between membrane and soluble pools is balanced by regulated re-as-sociation to membranes.
The cytosolic activity that mediates ␣-syn dissociation
clearly distinguishes between the Wt␣-syn and the PD-associ-ated mutants. Both A30P and A53T mutations doubled the cytosol-induced␣-syn off-rate but had no effect on the cytosol-independent dissociation. This was unexpected because several
studies, although not all (16), have previously suggested that the A30P mutation (but not A53T) impairs lipid binding (12, 14, 21, 38). Those studies measured the ability of
puri-fied recombinant ␣-syn to insert
spontaneously into purified or arti-ficial lipids and did not account for naturally existing cofactors or chap-erones. Indeed, we did not observe
differences in Wt or mutant␣-syn
expression in our mice or their par-titioning between soluble and mem-brane compartments, in agreement with studies on intact cultured neu-rons showing no effect on mem-brane interaction by the A30P mutation (17) and with other trans-genic lines where neither the ratio of soluble and particulate ␣-syn (39) nor its axonal transport (40) is affected by the A30P mutation. Furthermore, in our experiments, which employ biological mem-branes and include potential endog-enous factors that might participate
in ␣-syn membrane interactions,
A53T␣-syn was as likely to dissoci-ate in the presence of cytosol as
A30P ␣-syn, suggesting that the
mutations may confer a common pathogenic effect that is not evident when purified components are used to assess␣-syn membrane binding.
Our assay relies on ␣-syn-defi-cient cytosol as the acceptor frac-tion in order to detect the subtle movement that would otherwise be masked by the 30% of presynaptic ␣-syn that is freely diffusible in cyto-plasm. Although we did not address whether other peripherally associ-ated presynaptic proteins are also recruited into the cytosolic buffer along with␣-syn, this is a likely scenario since several presynaptic proteins are transiently associated with the mem-brane fraction and each has a distinct mechanism that regulates its direct or indirect membrane interaction (22–24). These are
mediated by either a membrane protein complex (e.g.
␣/-SNAP), phosphorylation (e.g. synapsin), Ca2⫹(e.g. CAPS), and nucleotide triphosphate hydrolysis (e.g. NSF, dynamin). In par-ticular, there are similarities between the dissociation of␣-syn and the behavior of rab3, a GTP-binding protein that under-goes regulated dissociation and association from vesicles with the aid of GDP-dissociation inhibitor (25–27). Rab3 lacks a transmembrane domain and its membrane-attachment is afforded by hydrophobic prenyl groups at its carboxyl terminus that insert into lipid bilayers (41). GDP-dissociation inhibitor likely shelters the prenyl moieties from aqueous cytoplasm
dur-FIGURE 7. A, proportion of cytoplasmic and total synuclein is not affected by PD mutations. Top panel,␣-Syn
expression levels and distribution in cytoplasm were compared between synaptosomes derived from Wt (lanes
1–3), A30P (lanes 4 – 6), and A53T (lanes 7–9)␣-syn Tg mice. A representative Western blot showing ␣-syn,
glyceraldehyde-3-phosphate dehydrogenase (GAPDH), and synaptophysin in the supernatant following resus-pension of 100 mg (wet tissue weight) intact synaptosomes into isotonic buffer (lanes 1, 4, and 7), hypotonic buffer (lanes 2, 5, and 8), or 1% Triton X-100 (lanes 3, 6, and 9). Bottom panel, quantification of five independent
experiments measuring the cytosolic (C; from lanes 2, 5, and 8) and total (T; from lanes 3, 6, and 9)␣-syn. No
significant differences were observed between Wt and mutant␣-syn expression (one-way ANOVA, cytosolic
fraction, p⫽ 0.51, n ⫽ 5; membrane fraction, p ⫽ 0.26, n ⫽ 5). B, left panel:␣-syn immunoreactivity was
compared between pellet fractions of the hypotonically-lysed synaptosomes from A (lanes 2, 5, and 8). Right
panel, no significant differences were observed in the amount of membrane bound␣-syn in hypotonically
lysed synaptosomes from␣-syn Wt and mutant Tg mice (one-way ANOVA, p ⬎ 0.8).
at UNIV DE LIEGE-MME F PASLE on December 1, 2008
www.jbc.org
ing extraction of rab3a from membranes. In the case of␣-syn, its amino-terminal is predicted to organize into an amphipathic ␣-helix that permits partial insertion of hydrophobic residues into the lipid bilayer (11). Regulated dissociation from mem-branes would require either unfolding of the␣-helix or segre-gation of those hydrophobic residues. Both mechanisms are compatible with a requirement for a cognate cytosolic factor that regulates␣-syn solubility. Notably, the differential effect
imparted by both PD-linked␣-syn mutations relative to Wt
␣-syn in the presence of cytosol implicates the as yet undefined cytosolic activity in the pathogenic process. Our approach described here to assess␣-syn membrane interactions can be used to identify and characterize the cytosolic activity that modulates␣-syn sequestration into and exchange between sub-cellular compartments. Importantly, the underlying mecha-nism may be amenable to experimental manipulation as a means of influencing␣-syn solubility in vivo so as to prevent its pathogenic accumulation.
REFERENCES
1. Lavedan, C. (1998) Genome Res. 8, 871– 880
2. Clayton, D. F., and George, J. M. (1999) J. Neurosci. Res. 58, 120 –129 3. Bennett, M. C. (2005) Pharmacol. Ther.105, 311–331
4. Kruger, R., Muller, T., and Riess, O. (2000) J. Neural Transm. 107, 31– 40 5. Ueda, K., Saitoh, T., and Mori, H. (1994) Biochem. Biophys. Res. Commun.
205,1366 –1372
6. Dawson, T. M., and Dawson, V. L. (2003) J. Clin. Invest. 111, 145–151 7. Zarranz, J. J., Alegre, J., Gomez-Esteban, J. C., Lezcano, E., Ros, R.,
Ampu-ero, I., Vidal, L., Hoenicka, J., Rodriguez, O., Atares, B., Llorens, V., Tor-tosa, E. G., Del Ser, T., Munoz, D. G., and de Yebenes, J. G. (2004) Ann.
Neurol. 55,164 –173
8. Jo, E., McLaurin, J., Yip, C. M., St George-Hyslop, P., and Fraser, P. E. (2000) J. Biol. Chem. 275, 34328 –34334
9. Eliezer, D., Kutluay, E., Bussell, R., Jr., and Browne, G. (2001) J. Mol. Biol.
307,1061–1073
10. Chandra, S., Chen, X., Rizo, J., Jahn, R., and Sudhof, T. C. (2003) J. Biol.
Chem. 278,15313–15318
11. Jao, C. C., Der-Sarkissian, A., Chen, J., and Langen, R. (2004) Proc. Natl.
Acad. Sci. U. S. A. 101,8331– 8336
12. Kubo, S., Nemani, V. M., Chalkley, R. J., Anthony, M. D., Hattori, N., Mizuno, Y., Edwards, R. H., and Fortin, D. L. (2005) J. Biol. Chem. 280, 31664 –31672
13. Maroteaux, L., and Scheller, R. H. (1991) Brain Res. Mol. Brain Res. 11, 335–343
14. Jensen, P. H., Nielsen, M. S., Jakes, R., Dotti, C. G., and Goedert, M. (1998)
J. Biol. Chem. 273,26292–26294
15. Fortin, D. L., Troyer, M. D., Nakamura, K., Kubo, S., Anthony, M. D., and Edwards, R. H. (2004) J. Neurosci. 24, 6715– 6723
16. Perrin, R. J., Woods, W. S., Clayton, D. F., and George, J. M. (2000) J. Biol.
Chem. 275,34393–34398
17. McLean, P. J., Kawamata, H., Ribich, S., and Hyman, B. T. (2000) J. Biol.
Chem. 275,8812– 8816
18. Sharon, R., Goldberg, M. S., Bar-Josef, I., Betensky, R. A., Shen, J., and Selkoe, D. J. (2001) Proc. Natl. Acad. Sci. U. S. A. 98, 9110 –9115 19. Lotharius, J., Barg, S., Wiekop, P., Lundberg, C., Raymon, H. K., and
Brundin, P. (2002) J. Biol. Chem. 277, 38884 –38894
20. Iwai, A., Masliah, E., Yoshimoto, M., Ge, N., Flanagan, L., de Silva, H. A., Kittel, A., and Saitoh, T. (1995) Neuron 14, 467– 475
21. Jo, E., Fuller, N., Rand, R. P., George-Hyslop, P., and Fraser, P. E. (2002) J.
Mol. Biol. 315,799 – 807
22. Lin, R. C., and Scheller, R. H. (2000) Annu. Rev. Cell Dev. Biol. 16, 19 – 49 23. Slepnev, V. I., and De Camilli, P. (2000) Nat. Rev. Neurosci. 1, 161–172 24. Sudhof, T. C. (2004) Annu. Rev. Neurosci. 27, 509 –547
25. Araki, S., Kikuchi, A., Hata, Y., Isomura, M., and Takai, Y. (1990) J. Biol.
Chem. 265,13007–13015
26. Fischer von Mollard, G., Sudhof, T. C., and Jahn, R. (1991) Nature 349, 79 – 81
27. Wu, S. K., Zeng, K., Wilson, I. A., and Balch, W. E. (1996) Trends Biochem.
Sci. 21,472– 476
28. Kozak, M. (1984) Nucleic Acids Res. 12, 857– 872 29. Kozak, M. (1991) J. Cell Biol. 115, 887–903
30. Scott, M. R., Kohler, R., Foster, D., and Prusiner, S. B. (1992) Protein Sci. 1, 986 –997
31. Citron, M., Westaway, D., Xia, W., Carlson, G., Diehl, T., Levesque, G., Johnson-Wood, K., Lee, M., Seubert, P., Davis, A., Kholodenko, D., Mot-ter, R., Sherrington, R., Perry, B., Yao, H., Strome, R., Lieberburg, I., Rom-mens, J., Kim, S., Schenk, D., Fraser, P., St George, H. P., and Selkoe, D. J. (1997) Nat. Med. 3, 67–72
32. Abeliovich, A., Schmitz, Y., Farinas, I., Choi-Lundberg, D., Ho, W. H., Castillo, P. E., Shinsky, N., Verdugo, J. M., Armanini, M., Ryan, A., Hynes, M., Phillips, H., Sulzer, D., and Rosenthal, A. (2000) Neuron 25, 239 –252 33. Tandon, A., Bannykh, S., Kowalchyk, J. A., Banerjee, A., Martin, T. F., and
Balch, W. E. (1998) Neuron 21, 147–154
34. Zhu, M., Li, J., and Fink, A. L. (2003) J. Biol. Chem. 278, 40186 – 40197 35. Fortin, D. L., Nemani, V. M., Voglmaier, S. M., Anthony, M. D., Ryan,
T. A., and Edwards, R. H. (2005) J. Neurosci. 25, 10913–10921
36. Irizarry, M. C., Kim, T. W., McNamara, M., Tanzi, R. E., George, J. M., Clayton, D. F., and Hyman, B. T. (1996) J. Neuropathol. Exp. Neurol. 55, 889 – 895
37. Kahle, P. J., Neumann, M., Ozmen, L., Muller, V., Odoy, S., Okamoto, N., Jacobsen, H., Iwatsubo, T., Trojanowski, J. Q., Takahashi, H., Wakaba-yashi, K., Bogdanovic, N., Riederer, P., Kretzschmar, H. A., and Haass, C. (2001) Am. J. Pathol. 159, 2215–2225
38. Bussell, R., and Eliezer, D. (2004) Biochemistry 43, 4810 – 4818
39. Kahle, P. J., Neumann, M., Ozmen, L., Muller, V., Jacobsen, H., Schindz-ielorz, A., Okochi, M., Leimer, U., van Der, P. H., Probst, A., Kremmer, E., Kretzschmar, H. A., and Haass, C. (2000) J. Neurosci. 20, 6365– 6373 40. Li, W., Hoffman, P. N., Stirling, W., Price, D. L., and Lee, M. K. (2004)
J. Neurochem. 88,401– 410
41. Goody, R. S., Rak, A., and Alexandrov, K. (2005) Cell Mol. Life Sci. 62, 1657–1670
at UNIV DE LIEGE-MME F PASLE on December 1, 2008
www.jbc.org