© Julien Chaillot, 2019
Étude de l'homéostasie de la taille chez la levure
opportuniste Candida albicans
Thèse
Julien Chaillot
Doctorat en biologie cellulaire et moléculaire
Philosophiæ doctor (Ph. D.)
III
Résumé
L’homéostasie de la taille est un processus important de la prolifération cellulaire mais les mécanismes moléculaires sont mal compris. Les cellules eucaryotes doivent atteindre une taille seuil avant la division, ce qui permet de maintenir une taille constante sur le long terme. Ce processus est régulé à un point de contrôle, à la fin de la phase G1, appelé START chez les levures. Le contrôle de la taille cellulaire a été étudié chez la levure modèle Saccharomyces cerevisiae mais n’a jamais été étudié chez les levures pathogènes. Dans cette thèse, nous avons utilisé Candida albicans comme organisme modèle pour étudier la régulation de la taille chez les levures opportunistes. Nous avons criblé des collections de mutants de délétions hétérozygotes et homozygotes de C. albicans afin d’identifier des gènes régulateurs de la taille.
Nous avons analysé la distribution de taille de 279 mutants homozygotes et 4 348 mutants hétérozygotes (recouvrant 90% du génome). Nous avons comparé nos résultats à différents criblages effectués sur la levure modèle S. cerevisiae. Ces comparaisons montrent que peu de régulateurs sont conservés entre C. albicans et S. cerevisiae et que la régulation de la taille est processus très plastique.
Par exemple, le mutant dot6 a un phénotype petit chez C. albicans mais n’a pas de phénotype de taille chez S. cerevisiae. Nous avons montré que Dot6 est un facteur de transcription nécessaire pour l’activation des gènes de la biogénèse des ribosomes. Dot6 est également un régulateur de START et joue un rôle dans l’adaptation de la taille suivant les sources de carbone disponibles.
Nous avons également mis en évidence un nouveau rôle pour la protéine kinase Hog1/p38 dans la régulation de la taille chez C. albicans en
IV
absence de stress. Ce rôle n’a jamais été démontré chez S. cerevisiae. Nous avons montré que Hog1, ainsi que toute la voie HOG, sont des régulateurs négatifs de START. Nous avons mis en évidence que Hog1 régule à la fois la croissance cellulaire via Sfp1, un régulateur majeur de la biogénèse des ribosomes et des protéines ribosomales, et le cycle cellulaire via le complexe SBF (Swi4/Swi6), des facteurs de transcription nécessaires pour la transition G1/S.
Nous avons également découvert qu’Ahr1, un facteur de transcription n’ayant pas d’orthologue chez S. cerevisiae, est nécessaire pour la régulation de la taille et aussi requis pour l’adaptation de la taille en fonction des acides aminés disponibles. Nous avons montré qu’Ahr1 agit dans la voie Tor1-Sch9 et régule négativement START.
En conclusion, notre travail a permis de découvrir de nouveaux régulateurs de START, de caractériser leur fonction et de les placer dans différentes voies. Comme la dérégulation de la voie Hog1/p38 est associée à des pathologies humaines, nous proposons C. albicans comme organisme modèle pour l’étude de cette voie et son implication dans l’homéostasie de la taille chez les organismes eucaryotes.
V
Abstract
Cell size homeostasis is an important process of cell proliferation but the molecular mechanisms are poorly understood. Eukaryotic cells must reach a threshold size before entering the cell cycle, which helps to maintain a constant size over the long term. This process is regulated at the end of the G1 phase, a check point called START. Cell size control has been studied in the model yeast Saccharomyces cerevisiae but has never been studied in pathogenic fungi. In this thesis, we used Candida albicans as a model organism to study the regulation of size in pathogenic yeasts. We have screened heterozygous and homozygous deletion collections of C. albicans to identify genes that control cell size.
We analyzed the size distribution of 279 homozygous mutants and 4,348 heterozygous mutants (covering 90% of the genome). We compared our results with different screens performed on the model yeast S. cerevisiae. These comparisons showed that few regulators were conserved between S. cerevisiae and C. albicans and suggesting that the cell size regulation is evolutionary plastic.
For example, dot6 mutant has a small phenotype in C. albicans but has no size phenotype in S. cerevisiae. We have shown that Dot6 is a transcriptional factor necessary for the activation of ribosome biogenesis genes. Dot6 is also a regulator of START and plays a critical role in adapting size according to the carbon sources available in the medium.
We also uncovered a novel stress-independent role of the Hog1/p38 MAPK in size regulation in C. albicans a role that has never been demonstrated in S. cerevisiae. We have shown that Hog1, as well as the entire HOG pathway, are negative regulators of START. We have shown that Hog1 regulates both growth via Sfp1, a major transcriptional regulator of
VI
ribosomal biogenesis and ribosomal proteins, and the cell cycle via the SBF complex (Swi4/Swi6), transcriptional factors necessary for the G1/S transition.
We also found that Ahr1, a transcription factor with no obvious ortholog in S. cerevisiae, has a role for the adaptation of the size according to the amino acids available in the medium. We have shown that Ahr1 is a negative START regulator and is controlled by the Tor1-Sch9 pathway.
In conclusion, our work has permitted to discover new regulators of START, to characterise their function and to map them in different pathways. As the Hog1/p38 pathway is linked to many human pathologies, we think that C. albicans is a useful model to study of this pathway and dissect its role in size control in eukaryotes.
VII
Table des matières
Résumé ... III Abstract ... V Table des matières ... VII Liste des figures ... X Liste des tableaux ... XI Liste des abréviations et des sigles ... XII Remerciements ... XVI Avant-propos ... XVIII
Introduction ... 1
Chapitre 1 - Genome-wide screen for haploinsufficient cell size genes in the opportunistic yeast Candida albicans. ... 40
1.1 - Résumé ... 40
1.2 - Article ... 41
1.2.1 - Abstract ... 42
1.2.2 - Introduction ... 43
1.2.3 - Materials and methods ... 45
1.2.4 - Results and discussion ... 50
1.2.5 - Acknowledgments ... 56
1.2.6 - References ... 57
1.2.7 - Figures ... 61
Chapitre 2 - The p38/HOG stress-activated protein kinase network couples growth to division in Candida albicans. ... 63
2.1 - Résumé ... 63 2.2 - Article ... 64 2.2.1 - Abstract ... 65 2.2.2 - Introduction ... 66 2.2.3 - Results ... 70 2.2.4 - Discussion ... 80 2.2.5 - Methods ... 86 2.2.6 - Acknowledgments ... 90 2.2.7 - References ... 91 2.2.8 - Figures ... 102
VIII
Chapitre 3 - Integration of growth and cell size via the TOR pathway and
the Dot6 transcription factor in Candida albicans. ... 114
3.1 - Résumé ... 114
3.2 - Article ... 115
3.2.1 - Abstract ... 116
3.2.2 - Introduction ... 117
3.2.3 - Materials and Methods ... 120
3.2.4 - Results ... 126
3.2.5 - Discussion ... 133
3.2.6 - Acknowledgments ... 137
3.2.7 - References ... 138
3.2.8 - Figures ... 143
Chapitre 4 - Caractérisation d’un nouveau régulateur de la taille : Ahr1 .. ... 155
4.1 - Le mutant ahr1 présente un phénotype de petite taille ... 155
4.2 - Ahr1 est un régulateur négatif de START ... 158
4.3 - Ahr1 interagit génétiquement et physiquement avec Sch9 ... 160
4.4 - Ahr1 régule la croissance suivant les acides aminés disponibles.... ... 162
4.5 - La localisation d’Ahr1 est régulée par la voie TOR... 165
4.6 - Discussion ... 168
4.7 - Matériels et Méthodes ... 171
Chapitre 5 - Discussion générale et perspectives ... 175
5.1 - Conservation des mécanismes du contrôle de la taille cellulaire .... ... 177
5.2 - La régulation de la taille cellulaire est un processus évolutif plastique ... 179
5.2.1 - Rôle de Hog1 dans le contrôle de la taille cellulaire ... 180
5.2.2 - Rôle de Dot6 dans le contrôle de la taille cellulaire ... 182
5.2.3 - Rôle de Ahr1 dans le contrôle de la taille cellulaire ... 185
5.3 - Lien entre nutriments et taille cellulaire ... 186
5.4 - Lien entre virulence et taille cellulaire ... 187
5.5 - C. albicans – Organisme modèle ... 189
Conclusion ... 191
IX
Annexe 1 - The monoterpene carvacrol generates endoplasmic reticulum
stress in the pathogenic fungus Candida albicans ... 212
Résumé ... 212
Article ... 213
Abstract ... 214
Introduction ... 215
Materials and methods... 217
Results ... 221
Discussion ... 227
Acknowledgments ... 229
References ... 230
Figures ... 236
Annexe 2 - pH-dependant antifungal activity of valproic acid against the Human fungal pathogen Candida albicans ... 244
Résumé ... 244
Article ... 245
Abstract ... 246
Introduction ... 247
Materials and methods... 250
Results ... 255
Discussion ... 261
References ... 265
X
Liste des figures
Figure 1 - Modèles de régulation de la taille cellulaire 3
Figure 2 - Principe du Coulter Counter 4
Figure 3 - Modèle de régulation de la taille bactérienne par DnaA 6 Figure 4 - Régulation des gènes RiBi et RP 10 Figure 5 - Cycle cellulaire de S. cerevisiae 11 Figure 6 - Processus de la transition G1/S 12 Figure 7 - Corrélation négative entre la taille et le temps passé en G1 13 Figure 8 - Régulation de la taille chez S. pombe 17 Figure 9 - Diversité de la taille de Cryptococcus neoformans 21 Figure 10 - Phylogénie des clades Candida et Saccharomyces 24 Figure 11 - Formes morphologiques de C. albicans 28 Figure 12 - C. albicans en forme pseudohyphe, levure et hyphe 30 Figure 13 - Formes phénotypiques de C. albicans 32 Figure 14 - Distribution de la taille du mutant ahr1 156 Figure 15 - Courbes de croissance du mutant ahr1 et du WT 157 Figure 16 - Taille des hyphes du mutant ahr1 et du révertant 158 Figure 17 - Budding index du mutant ahr1 et du WT 159 Figure 18 - Expression de RNR1 et PCL2 en fonction de la taille 159 Figure 19 - Intéraction génétique entre SCH9 et AHR1 161 Figure 20 - Coimmunoprécipitation entre Ahr1 et Sch9 162 Figure 21 - Analyse transcriptionnelle du mutant ahr1 163 Figure 22 - Temps de doublement des mutants ahr1 et sch9 164 Figure 23 - Taille cellulaire en fonction du temps de doublement 165 Figure 24 - Photos de microscopie d’une souche exprimant Ahr1-GFP 166 Figure 25 - Modèle de la régulation de START 169 Figure 26 - Interaction génétique entre Nrm1 et le complexe SBF 178 Figure 27 - Effet de la doxycycline sur la taille du mutant dot6 185
XI
Liste des tableaux
Tableau 1 - Oligonucléotides utilisées dans cette étude 173 Tableau 2 - Souches utilisées dans cette étude 174
XII
Liste des abréviations et des sigles
ADN Acide désoxyribonucléiqueAhr1 Adhesion and Hyphal Regulator AmB Amphotéricine B
ARN Pol Acide Ribonucléique Polymerase ARNm Acide Ribonucléique messager ATP Adenosine Triphosphate
bp base pair
Ca Candida albicans
CDK Cyclin-Dependant Kinase CFU Colony-Forming Unit
CGD Candida Genome Database CHI Complex happloinsuffisiency ChIP Chromatin immunoprecipitation
CRISPR Clustered Regularly Interspaced Short Palindromic Repeats CSP Caspofungine
CTZ Clotrimazole CWI Cell Wall Integrity
DAPI 4',6-diamidino-2-phénylindole DBC Double Barcode
DMSO diméthylsulfoxyde
DNA Deoxyribose Nucleic Acid ds Double Strand
dNTP DeoxyNucleotide TriPhosphate DO Densité Optique
Dot6 Disruptor Of Telomeric silencing DTT Dithiothréitol
ER Endoplasmic Reticulum
F Forward
XIII FCZ Fluconazole
FDR False Discovery Rate
FIC Fractional Inhibitory Concentration FISH Fluorescence In Situ Hybridization FITC Fluorescein isothiocyanate
fL femtolitre G1 Gap 1 G2 Gap 2
G3 Genes, Genomes, Genetics GFP Green Fluorescent Protein
GRACE Gene Replacement And Conditional Expression GO Gene Ontology
GSEA Gene Set Enrichment Analysis GTP Guanosine Triphosphate
GUT Gastrointestinally induced Transition HA human influenza hemagglutinin
HCGP Haploid deletion Chemical-Genetic Profiling
HDAC Histone Deacetylase
HIV Human Immunodeficiency Virus Hog1 High Osmolarity Glycerol response IGF Insulin-like Growth Factor
ITZ Itraconazole
Kog1 Kontroller Of Growth
Lge Désigne un phénotype de grande taille LDH Lactate Deshydrogénase
M Mitosis
MAPK Mitogen-activated protein kinases Mb Megabase
MBF MCB-binding factor MCF Micafungine
XIV mg milligramme
MIC Minimum Inhibitory Concentration Min minutes
mL millilitre mM millimolaire MM Milieu Minimum MoA Mechanism of Action
mTOR mechanistic Target Of Rapamycin ND Non determined
ng nanogramme NS Non Significatif
ORF Open Reading Frame
OriC Origine de réplication d’E. coli PAC Polymerase A and C
PalmC palmitoylcarnitine
PBS Phosphate Buffered Saline PCR Polymerase Chain Reaction pg picogramme
pH Potentiel Hydrogène PKA Protein Kinase A PI Propidium Iodide
PI3K phosphatidylinositol 3-kinase PMSF Fluorure de phénylméthylsulfonyle PVDF PolyVinyliDene Fluoride
R Reverse
Rb Retinoblastoma protein RiBi Ribosome Biogenesis RNA Ribonucleic acid RP Ribosomal Protein
RPMI Roswell Park Memorial Institute medium S Synthesis
XV
SARM Staphylococcus aureus Résistant à la Méticilline SBF SCB-binding factor Sc Saccharomyces cerevisiae SC Synthetic Complete SCB SBF cell-cycle box SD Standard deviation SDS dodécylsulfate de sodium Sec Seconde
SGD Saccharomyces Genome Database
SIDA Syndrome d'Immuno Déficience Acquise
SILAC Stable Isotope Labeling by Amino acids in Cell culture Tco89 Tor Complex One
TCZ Teroconazole
TGF Transforming Growth Factor Tod6 Twin Of Dot6
TOR Target Of Rapamycin TRB Terbinafine
µg microgramme µM micromollaire
UPR Unfolded Protein Response VPA Valproic Acid (Acide valproïque) VVC Vulvovaginal Candidiasis
WGD Whole Genome Duplication
Whi Désigne un phénotype de petite taille WT Wild Type
XTT tetrazolium salt
XVI
Remerciements
Je tiens à remercier mon directeur de thèse, Dr Adnane Sellam, de m’avoir accueilli dans son laboratoire et pour son soutien tout au long de mon doctorat.
Je remercie également tous les membres de l’équipe avec qui j’ai travaillé pendant toutes ces années : Anaïs Burgain, Faiza Tebbji, Carlos Garcia, Emilie Pic et Inès Khemiri.
Merci à Julie-Christine Lévesque pour son aide en microscopie et en cytométrie.
Je remercie les coauteurs qui ont participé aux projets publiés dans cette thèse : Mike Tyers (Université de Montréal), Jaideep Mallick (Université de Montréal), Michael A. Cook (Samuel Lunenfeld Research Institute) et Jacques Corbeil (Université Laval) ; ainsi que tous les collaborateurs qui nous ont fourni des souches et des plasmides : Catherine Bachewich (Université Concordia), Joachim Ernst (Université de Düsseldorf), Joseph Heitman (Université de Duke), Julia Köhler (Boston Children's Hospital), Daniel Kornitzer (Technion), Christian Landry (Université Laval), Robbie Loewith (Université de Genève), Aaron Mitchell (Université Carnegie Mellon), André Nantel (National Research Council Canada), Suzanne Noble (Université de Californie à San Francisco), Janet Quinn (Université de Newcastle), Dominique Sanglard (Université de Lausanne), Ana Traven (Université Monash), Jonathan Thorner (Université de Californie Riverside) et Malcolm Whiteway (Université Concordia).
Je remercie également les membres du jury, Christian Landry, Hugo Wurtele et Yves Bourbonnais d’avoir accepté d’évaluer mon manuscrit et ma soutenance de thèse.
XVII
Enfin, je remercie mes parents, Jean-Yves et Marie Dominique, mon frère David ainsi que ma conjointe Carolina d’être présents pour moi.
XVIII
Avant-propos
L’article du chapitre 1 est intitulé “Genome-Wide Screen for Haploinsufficient Cell Size Genes in the Opportunistic Yeast Candida albicans”. Il a été publié le 9 février 2017 dans le journal G3 (Genes, Genomes, Genetics). Je suis le premier auteur de cet article. J’ai effectué les vérifications de taille pour valider le criblage et j’ai participé à l’analyse des résultats. Michael Cook et Jacques Corbeil ont participé aux analyses bioinformatiques du séquençage. Adnane Sellam a déterminé le « Budding Index », a participé à l’analyse des résultats, a rédigé l’article et a supervisé le projet.
L’article du chapitre 2 est intitulé “The p38/HOG stress-activated protein kinase network couples growth to division in Candida albicans. Cet article a été publié en mars 2019 dans le journal Plos Genetics. J’ai effectué une partie du criblage et les tests d’interaction génétique. Adnane Sellam et Julien Richard Albert ont réalisé le criblage. Jaideep Malick a réalisé les co-immunoprécipitations. Michael A Cook a réalisé les analyses bio-informatiques. Faiza Tebbji a réalisé les ChIP-chip. Adnane Sellam est le premier auteur de cet article. Il a réalisé les puces à ADN, les ChIP-chip, les ChIP-qPCR, a rédigé l’article et a supervisé le projet avec Mike Tyers.
L’article du chapitre 3 est intitulé “Integration of Growth and Cell Size via the TOR Pathway and the Dot6 Transcription Factor in Candida albicans”. Il a été publié en février 2019 dans le journal Genetics. Je suis le premier auteur de cet article. J’ai créé les souches nécessaires pour l’étude, j’ai fait les tests d’interactions génétiques, les expériences de microscopie, les courbes de croissance, déterminer la distribution de taille des souches et j’ai participé à la rédaction de l’article. Jaideep Malick et Faiza Tebbji ont réalisé les Western-Blot. Adnane Sellam a réalisé les puces à ADN, les
RT-XIX
qPCR et la détermination des « Budding Index ». Il a également rédigé l’article et supervisé le projet.
Le chapitre 4 est intitulé « Caractérisation d’un nouveau régulateur de la taille : Ahr1 ». Ce chapitre présente des résultats non publiés. Adnane Sellam a réalisé les puces ADN et la détermination du « Budding Index ». Jaideep Malick a réalisé l’expérience de co-immunoprécipitation. J’ai créé les mutants utilisés dans l’étude, fait les analyses de cytométrie en flux, de microscopie ainsi que les courbes de croissance et déterminé la distribution de taille des souches.
L’article de l’annexe 1 est intitulé « The Monoterpene Carvacrol Generates Endoplasmic Reticulum Stress in the Pathogenic Fungus Candida albicans ». Il a été publié le 26 mai 2015 dans le journal Antimicrobial Agents & Chemotherapy. Je suis co-premier auteur de l’article avec Faiza Tebbji. J’ai réalisé les expériences de microscopie, les RT-PCR et les tests de synergisme. Faiza Tebbji a réalisé les tests de sensibilité au carvacrol. Charles Boone, Mohamed Bellaoui et Grant Brown ont réalisé les essais chemogénétiques sur les souches haploïdes de Saccharomyces cerevisiae. Mohamed Bellaoui et Adnane Remmal sont les premiers à avoir observé l’effet du carvacrol sur S. cerevisiae et C. albicans. Adnane Sellam a supervisé le projet et a rédigé l’article.
L’article de l’annexe 2 est intitulé « pH-Dependant Antifungal Activity of Valproic Acid against the Human Fungal Pathogen Candida albicans ». Il a été publié le 9 octobre 2017 dans le journal Frontiers in Microbiology. Je suis le premier auteur de cet article. J’ai réalisé les tests de sensibilité à l’acide valproïque, les tests de synergismes et la microscopie confocale. Faiza Tebbji a réalisé les expériences testant l’effet protecteur du VPA sur des cellules épithéliales humaines. Carlos Garcia a évalué l’efficacité du VPA sur les souches cliniques résistantes et sur les biofilms. René Pelletier
XX
a fourni les souches cliniques. Adnane Sellam a supervisé le projet et a rédigé l’article.
J’ai également participé à l’article « A phenotypic small-molecule screen identifies halogenated salicylanilides as inhibitors of fungal morphogenesis, biofilm formation and host cell invasion.” Les auteurs sont Carlos Garcia, Anaïs Burgain, Émilie Pic, Inès Khemiri et Adnane Sellam. L’article a été publié le 1er aout 2018 dans le journal Scientific Reports. Cet article ne sera pas discuté dans cette thèse (Garcia et al. 2018).
1
Introduction
L’homéostasie de la taille
Généralités
Les organismes vivants ont une différence de masse allant de moins de 0,1pg (par exemple les mycoplasmes) à plus de 1 000 tonnes (par exemple les séquoias). Il y a donc une différence de l’ordre de 1022. Parmi les espèces unicellulaires, la différence de masse est de l’ordre de 109, les plus petites étant les mycoplasmes et les plus grandes les amibes. Dans un même organisme, la taille de différents types de cellules varient grandement, de quelques micromètres pour les cellules sanguines chez l’Homme, jusqu’à un mètre pour les neurones, par exemple.
La taille d’une cellule est limitée par sa surface. Plus une cellule est grande, plus le rapport surface/volume diminue, ce qui est un désavantage pour les échanges entre la cellule et le milieu extérieur. La taille est également limitée par la diffusion passive. En effet, la diffusion passive (des nutriments ou de l’oxygène), est inefficace sur des longues distances (Schulz and Jorgensen 2001), une taille optimale semble donc être nécessaire pour assurer les échanges entre la cellule et le milieu extérieur ainsi que pour le transport intracellulaire. Par exemple, certains processus comme la transcription ou la traduction se réalisent généralement proche du centre de la cellule, ce qui peut créer un gradient de métabolites ou de nutriments dans les cellules de grande taille.
Dans une population de cellules en division, il y a habituellement peu de variation de taille. Si la distribution de la taille est modifiée, par exemple par un changement de nutriment, la distribution de taille revient à celle précédent le changement. Ces observations indiquent que les cellules en division contrôlent leurs tailles de façon active et en réponse à
2
l’environnement. Ces observations montrent aussi que les cellules adaptent leurs tailles suivant l’environnement, probablement pour optimiser le rapport surface/volume afin de favoriser les échanges avec le milieu extérieur. De plus, ceci suggère qu’il y a une coordination entre les processus contrôlant la croissance (biomasse) et la division afin de maintenir la taille.
La taille cellulaire dépend également de la ploïdie. Par exemple la levure Saccharomyces cerevisiae est plus petite en phase haploïde qu’en phase diploïde. Ce phénomène pourrait être expliqué par le fait que l’augmentation de la ploïdie augmente également le taux de transcription, puis le taux de traduction et ainsi la cellule produit plus de masse. L’augmentation de la ploïdie est une stratégie utilisée chez l’Homme pour former des cellules géantes. Par exemples, les mégacaryocytes augmentent la ploïdie pour former des cellules de 100µm de diamètre. Ensuite, ces cellules se fragmentent pour former des thrombocytes (Mazzi et al. 2018).
Conceptuellement, l’homéostasie de la taille peut être régulée de différentes façons (Figure 1) :
-par un mécanisme « Timer » dans lequel la division se produit après un temps fixe depuis la naissance de la cellule fille.
-par un mécanisme « Adder » dans lequel la cellule ajoute un volume constant après chaque division (Taheri-Araghi et al. 2015).
-par un mécanisme « Sizer » dans lequel la cellule fille doit atteindre une taille seuil pour se diviser.
3
Figure 1 – Modèles de régulation de la taille cellulaire. Dans le modèle « Timer », la
cellule grossit pendant un temps fixe avant de se diviser. Dans le modèle « Adder », la cellule ajoute un volume constant avant de se diviser, indépendamment de la taille à la naissance. Dans le modèle « Sizer », la cellule doit atteindre une taille seuil avant de se diviser, indépendamment de la taille de la cellule à la naissance (Varsano, Wang, and Wu 2017).
Techniques pour l’analyse de la taille cellulaire
Coulter Counter
Le Coulter Counter est un appareil inventé par Wallace H. Coulter dans les années 1940 et breveté en 1953. Il permet de quantifier des particules ou des cellules et de mesurer leur volume. L’appareil est utilisé pour la quantification de microbes et de cellules mammifères. Il est
4
particulièrement utilisé en biologie médicale pour réaliser des hémogrammes. Pour mesurer le volume des cellules, elles sont mises en suspension dans un électrolyte et sont aspirées par une pompe. Les cellules passent entre des électrodes et n’étant pas conductrices, elles génèrent une variation de l’impédance qui est proportionnelle au volume de la cellule (Figure 2).
Figure 2 - Principe du Coulter Counter. Les cellules sont en suspension dans un électrolyte.
La solution est aspirée par une pompe et des électrodes mesurent l’impédance provoquée par les cellules. L’impédance est proportionnelle au volume des cellules. L’ordinateur produit un graphique avec la fréquence en fonction du volume, en femtolitre (fL). Ici, par exemple le WT a un volume médian d’environ 50 fL et le mutant de délétion sfp1 a un volume médian d’environ 25 fL.
Microscopie
La taille cellulaire peut être quantifiée par microscopie optique. Cette méthode a été utilisée par Navarro et Nurse pour mesurer la longueur et la
5
largeur de S. pombe (Navarro and Nurse 2012). Cependant, cette méthode a le désavantage d’être fastidieuse.
Calcul de la biomasse
Certains auteurs estiment la taille en mesurant la quantité de protéines ou d’ARN dans la cellule. Par exemple, la quantité de protéines peut être déterminée par marquage au FITC. Le marquage par le FITC peut être analysé par cytométrie de flux (Cipollina et al. 2005).
Régulation de la taille des organismes procaryotes
La taille des bactéries varie énormément, elle va de 10-4 fL pour les bactéries marines du genre Candidatus (ce volume représente environ 1% d’une bactérie E. coli) à 108 fL pour Thiomargarita namibiensis (8 fois plus volumineuse qu’E. coli) (Young 2006). Cette bactérie est plus grande que l’œil d’une drosophile et peut être visible à l’œil nu (Schulz et al. 1999).
Régulation de la taille des Bacilles – Division symétrique
La taille des cellules bactériennes est influencée par le milieu (Schaechter, Maaloe, and Kjeldgaard 1958; Trueba and Woldringh 1980) et elle est corrélée au taux de croissance (Monds et al. 2014). Dans un milieu riche, le temps de génération est moins élevé qu’en milieu pauvre, alors que la taille des cellules est plus élevée en milieu riche qu’en milieu pauvre.
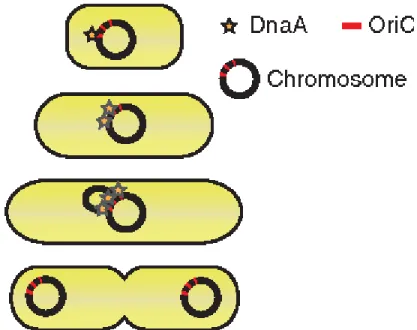
L’existence d’une molécule capable de mesurer la taille de la cellule est débattue (Robert 2015). DnaA, une molécule activatrice de la réplication de l’ADN, a été proposée comme une protéine contrôlant la taille cellulaire (Lobner-Olesen et al. 1989) : l’augmentation de son expression réduit la taille cellulaire, suggérant que la concentration en DnaA influence la taille cellulaire. Dans ce modèle, les cellules grossissent et il y a accumulation de la molécule DnaA à un niveau suffisant pour initier la réplication de l’ADN (Figure 3). Donc, les cellules de petites tailles devraient grossir plus
6
longtemps afin d’accumuler assez de DnaA, ce qui permet de maintenir l’homéostasie de la taille.
Figure 3 – Accumulation de DnaA, dépendamment de la taille, sur l’origine de réplication
du chromosome jusqu’à ce qu’un seuil soit atteint pour initier la réplication de l’ADN. (Amodeo and Skotheim 2016)
Une autre protéine candidate est FtsZ, une GTPase essentielle dans la formation du septum et permettant la cytocinèse. La réduction de l’expression de cette protéine engendre des cellules de grande taille (Palacios, Vicente, and Sanchez 1996). FtsZ contrôle la division de façon spatiale et temporelle. Cette protéine est inhibée par MinC, qui se situe aux extrémités de la cellule et crée un gradient jusqu’au centre. Pendant l’élongation, la concentration en MinC devient plus faible au centre de la cellule, ce qui permet l’activation de FtsZ (Rothfield, Taghbalout, and Shih 2005; Lutkenhaus 2008).
Des études sur B. subtilis, P. aeruginosa et E. coli ont montré que la taille ajoutée entre la cellule fille (sb) et la division (sd) est constante : Δ= sd- sb
7
(Taheri-Araghi et al. 2015; Deforet, van Ditmarsch, and Xavier 2015). Le volume ajouté varie suivant les conditions de cultures. Ceci signifie que les bactéries régulent la taille en ajoutant un volume constant à chaque génération, donc il s’agit d’un mécanisme « Adder ». Du fait que ces bacilles se divisent de façon symétrique, ce mécanisme permet de maintenir la taille sur le long terme.
Archéobactéries
Les archéobactéries ont une taille comparable aux bactéries. Les mécanismes contrôlant le cycle cellulaire partagent des similarités avec les procaryotes et avec les eucaryotes. Par exemple, la protéine FtsZ est conservée chez les bactéries et les archéobactéries. Comme chez les eucaryotes, le cycle cellulaire est divisé en quatre phases : G1, S, G2 et M (Lindås and Bernander 2013).
Halobacterium salinarum, une archéobactérie halophile, maintient sa taille en ajoutant un volume constant entre chaque division. Le modèle « Adder » semble donc être partagé entre les bactéries et les archéobactéries (Eun et al. 2018).
Régulation de la taille des eucaryotes unicellulaires
Parmi les eucaryotes unicellulaires, S. cerevisiae et S. pombe ont été utilisés pour l’étude du cycle cellulaire et la régulation de la taille (Sveiczer and Horvath 2017; Wood and Nurse 2015; Jorgensen et al. 2002). S. cerevisiae est une levure qui se divise par bourgeonnement alors que S. pombe se divise de façon symétrique. Due à cette différence morphologique, la taille semble être régulée à la transition G1/S pour S. cerevisiae et à la transition G2/M pour S. pombe.
Saccharomyces cerevisiae Croissance cellulaire
8
La croissance, c’est-à-dire la production de biomasse, est importante dans le contrôle de la taille cellulaire. La croissance est assurée par l’accumulation de masse cellulaire via les macromolécules : protéines, acides nucléiques, sucres et lipides. La synthèse des protéines est un processus indispensable pour la croissance cellulaire. Ce processus est réalisé par les ribosomes. En phase de croissance, la cellule doit assurer une synthèse abondante des ribosomes d’environ 2 000 par minute (Warner 1999). La biogénèse des ribosomes est corrélée aux conditions environnementales et est régulée essentiellement par la voie TOR (Target Of Rapamycin).
De nombreuses expériences indiquent que la traduction joue un rôle fondamental dans la régulation de la taille cellulaire. Premièrement, le traitement des cellules avec des inhibiteurs de la traduction, comme la cycloheximide, provoque un défaut de taille (Popolo, Vanoni, and Alberghina 1982). Ensuite, il a été montré que les souches déficientes en gènes de la biogénèse des ribosomes et de protéines ribosomales ont un phénotype de petite taille (Jorgensen et al. 2002; Soifer and Barkai 2014). Les plus petites souches identifiées sont des mutants de délétions de Sfp1 et Sch9, qui sont deux régulateurs de la biogénèse des ribosomes, suggérant que ce processus est important pour la croissance et le contrôle de la taille. De plus, il a été montré que muter des gènes codants la petite sous unité des ribosomes provoque un phénotype de grande taille, alors que les mutants de la grande sous unité présentent un phénotype de petite taille. Ces observations suggèrent que l’initiation de la traduction ainsi que l’élongation ont un rôle dans la régulation de la taille cellulaire (Soifer and Barkai 2014). Enfin, les souches déficientes en facteurs d’élongation de la traduction ainsi qu’en facteurs d’initiation ont également un défaut de taille (Jorgensen et al. 2002).
9
TOR est un complexe protéique situé au niveau des vacuoles (Urban et al. 2007; Sturgill et al. 2008). Chez S. cerevisiae, il existe deux complexes TOR : TORC1 et TORC2. TORC1 est composée des protéines Tor1 ou Tor2, Kog1, Tco89 et Lst8. TORC2 est composé de Tor2, Lst8, Avo1, Avo2, Avo3, Bit2 et Bit61 (Loewith et al. 2002; Wedaman et al. 2003; Reinke et al. 2004). TOR permet de percevoir les nutriments dans le milieu extérieur (azote, carbone) et aussi les facteurs de stress (De Virgilio and Loewith 2006). TOR favorise la croissance via l’augmentation de la traduction, la transcription des gènes nécessaires pour la biogénèse des ribosomes et de la glycolyse (Averous and Proud 2006; Ma and Blenis 2009; Laplante and Sabatini 2012). TOR contrôle la biogénèse des ribosomes via Sch9, Sfp1, Rrn3 et Maf1 (Rohde et al. 2008).
Sch9 est une kinase qui contrôle la biogénèse des ribosomes en régulant les ARN polymérases I, II et III (Lee, Moir, and Willis 2009; Huber et al. 2011). La régulation de la biogénèse des ribosomes se fait également via les facteurs de transcriptions Dot6 et Tod6 (Figure 4) (Huber et al. 2011). Sch9 régule aussi l’initiation de la traduction en phosphorylant la protéine ribosomale Rps6 et le facteur d’initiation eIF2 (Urban et al. 2007).
Sfp1, un facteur de transcription, régule la transcription des protéines ribosomales et des protéines nécessaire à la biogénèse des ribosomes (Figure 4) (Marion et al. 2004; Blumberg and Silver 1991; Jorgensen et al. 2002).
Rrn3 est un facteur de transcription requis pour la transcription de l’ADN ribosomal, régulé par l’ARN Pol I (Claypool et al. 2004).
10
Figure 4 Régulation des gènes de la biogénèse des ribosomes (RiBi) et les protéines
ribosomales (RP). Sfp1 se fixe sur les séquences RRPE pour activer les gènes RiBi et RP. Dot6 et Tod6 se fixent sur les séquences PAC pour inhiber les gènes RiBi. (Loewith and Hall 2011).
Cycle cellulaire
Le cycle cellulaire eucaryote est une succession de processus qui permet à une cellule de donner naissance à deux cellules filles identiques. Il est composé de la phase G1, dans laquelle la cellule grossit et prépare la phase S, qui est la phase dans laquelle la cellule duplique son génome. Ensuite, il y a la phase G2 et enfin la phase M, phase dans laquelle la cellule se divise.
Le cycle cellulaire est régulé par des CDK (Cyclin Dependant Kinase) dont l’activité est régulée par des cyclines. Chez S. cerevisiae, neuf cyclines sont connues : Cln1, Cln2, Cln3 qui contrôlent la transition G1/S ; Clb5 et Clb6 qui contrôlent la phase S et Clb1, Clb2, Clb3 et Clb4 qui contrôlent
11
les phases G2 et M (Figure 5). Cdc28 (ou CDK1) est la seule CDK qui contrôle le cycle cellulaire chez S. cerevisiae, ainsi que chez C. albicans.
Figure 5 – Cycle cellulaire de S. cerevisiae. La Cdk1 est la seule Cdk qui contrôle le cycle
cellulaire. Les Cln1, 2 et 3 régulent la transition G1/S. Clb5 et 6 régulent la synthèse de l’ADN (phase S). Les cyclines B 1 à 4 régulent la transition G2/M. Schéma adapté de https://studentreader.com/KF645/mitosis-factors-cyclin-cdks/
Les cyclines contrôlent l’activité kinase de la protéine Cdc28 et permettent l’activation de facteurs de transcriptions dans l’ordre nécessaire pour la progression du cycle cellulaire (Haase and Wittenberg 2014).
Les complexes Cln3/Cdc28 et Cln1-2/Cdc28 permettent l’activation des complexes SBF (Swi4/Swi6) et MBF (Mbf1/Swi6) en inhibant les inhibiteurs Whi5 et Nrm1 (Figure 6). SBF et MBF sont positionnés sur les promoteurs des gènes permettant l’entrée en phase S (Ferrezuelo et al. 2010; Simon et al. 2001; de Bruin et al. 2006) .
Clb5-6/Cdc28 initient la synthèse de l’ADN (Schwob et al. 1994) et inhibent l’activité des complexes Cln1-3/Cdc28 (Basco, Segal, and Reed 1995).
12
Clb1-2/Cdc28 activent Mcm1 et Fkh2 qui régulent la transition G2/M. Ceci permet l’expression d’environ 35 gènes, dont Ace2, Swi5 et Cdc5, afin d’activer la mitose (Cho et al. 1998).
La transition M/G1 est régulée par Mcm1 qui est lui-même réprimé par Yox1 et Yhp1. Mcm1 permet la synthèse de Swi4 et Cln3 et des gènes de pré-réplication de l’ADN (Cdc6, Mcm2-7).
Figure 6 - Processus de la transition G1/S (Haase and Wittenberg 2014). Le complexe SBF
est inhibé par Whi5. Cln3/Cdk1 inhibe Whi5, ce qui permet au complexe SBF de transcrire les cyclines 1 et 2. Cln3/Cdk1 active également le complexe MBF. Cln1-2/Cdk1 activent à leurs tours les complexes SBF et MBF. Le complexe MBF transcrit Nrm1, qui est un inhibiteur de MBF et permet la sortie de la phase G1. Clb2/Cdk1 inhibe les complexes SBF et MBF.
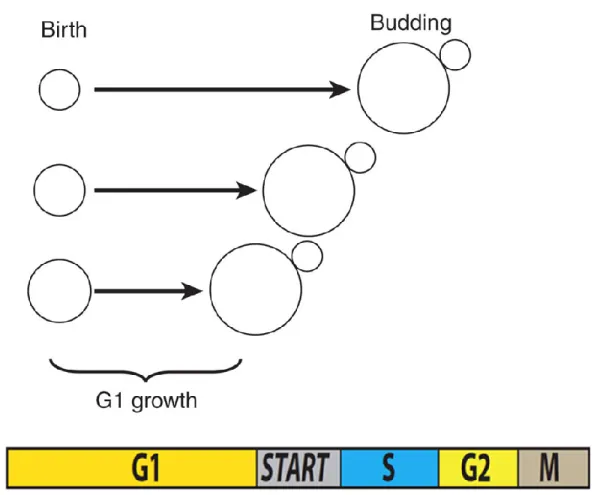
START : modèle « Sizer »
En 1974, Leland Hartwell a proposé que la croissance et la division soient couplées à START, un point de contrôle situé en fin de phase G1 (Hartwell et al. 1974). À START, les cellules arrêtent le cycle cellulaire en réponse aux hormones, à des stress ou à une carence nutritionnelle. De plus, les cellules doivent atteindre une taille seuil pour pouvoir passer START et se
13
diviser. Comme S. cerevisiae se divise de façon asymétrique, la cellule fille est plus petite que la cellule mère et doit passer plus de temps en phase G1 pour passer START (Figure 7). Ce point de contrôle permet d’assurer le maintien de la taille cellulaire sur le long terme. START est également modulé par le milieu extracellulaire : les cellules cultivées en milieu riche ont une plus grande taille qu’en milieu pauvre.
Figure 7 – Corrélation négative entre la taille cellulaire à la naissance et le temps passé en
G1. Figure adaptée de (Turner, Ewald, and Skotheim 2012).
La cycline 3 (Cln3) est une protéine importante de START. Cln3 est nécessaire pour la régulation temporelle du cycle cellulaire, mais n’est pas nécessaire à la viabilité chez S. cerevisiae, contrairement à l’homologue Cln3 chez C. albicans (Chapa y Lazo, Bates, and Sudbery 2005). Cette
14
observation suggère qu’il existe une autre voie chez S. cerevisiae qui permet de passer START. Cette fonction redondante serait controlée par Bck2 (Wijnen and Futcher 1999).
La concentration de Cln3 augmente pendant la phase G1 et se fixe à la CDK Cdc28. Ce complexe permet la phosphorylation de Whi5 (Tyers, Tokiwa, and Futcher 1993; Dirick, Bohm, and Nasmyth 1995; Stuart and Wittenberg 1995). Cette phosphorylation lève l’inhibition de Whi5 sur le complexe SBF (Figure 6) (facteurs de transcription Swi4/Swi6) (Costanzo et al. 2004; de Bruin et al. 2004; Schaefer and Breeden 2004). Whi5 est par conséquent délocalisé du noyau vers le cytoplasme par la protéine Msn5 (Taberner, Quilis, and Igual 2009). Cln3/Cdc28 phosphoryle également le complexe SBF, mais cette phosphorylation ne semble pas être nécessaire pour l’activation de SBF (Geymonat et al. 2004).
L’activation de SBF et MBF permet la synthèse d’environ 200 gènes nécessaire pour la synthèse de l’ADN et pour le bourgeonnement. Parmi ces gènes, les cyclines CLN1 et CLN2 sont transcrites afin d’assurer la poursuite du cycle cellulaire en s’associant avec Cdc28 et en continuant l’inactivation de Whi5 (Eser et al. 2011; de Bruin et al. 2004), ce qui forme une boucle d’activation. Cette boucle d’activation permet l’engagement irréversible du cycle cellulaire (Skotheim et al. 2008; Charvin et al. 2010; Doncic and Skotheim 2013). Cln1-Cln2/Cdc28 phosphorylent également Sic1, un inhibiteur des complexes Clb/Cdc28 (Verma, Feldman, and Deshaies 1997; Feldman et al. 1997). La phosphorylation de Sic1 permet son ubiquitination et sa dégradation, permettant l’activation de Clb5-6/Cdc28 et activant la réplication de l’ADN (Nash et al. 2001; Schneider, Yang, and Futcher 1996; Tyers 1996). Clb5 a également un rôle dans la phosphorylation de Sic1, ce qui permet encore d’amplifier la boucle d’activation (Yang et al. 2013).
15
Comme les levures sont capables de contrôler activement leurs tailles, ceci suggère qu’il existe un ou plusieurs senseurs. Le senseur qui permet d’indiquer à la cellule la taille n’est pas encore connu. Néanmoins, Cln3 et Whi5 sont les deux principaux candidats.
Cln3 est séquestrée dans le réticulum endoplasmique par la protéine Ydj1 (Verges et al. 2007). En fin de la phase G1, la quantité d’Ydj1 devient limitante et Cln3 est libéré et s’accumule par la suite dans le noyau pour enclencher la phase S. D’autres auteurs ont émis l’hypothèse que c’est l’accumulation de l’activité de Cln3/Cdc28 qui permet d’activer la transition G1/S (Schneider et al. 2004).
Whi5 joue également un rôle essentiel dans la régulation de START. La concentration nucléaire en Whi5 diminue pendant la phase G1. La protéine étant diluée, elle ne joue plus son rôle d’inhibition de SBF et MBF (Schmoller et al. 2015). Il a également été montré que la concentration en Swi4 augmente pendant la phase G1, ce qui fait diminuer le ratio Whi5/Swi4 (Dorsey et al. 2018). Donc, la quantité de Swi4 augmente au cours de la phase G1 et permet le passage de START.
Modèle « Adder »
Le couplage entre la croissance et la division à START permet de contrôler la taille sur le long terme. Cependant, une cellule fille née à une taille plus petite passera plus de temps en G1 mais bourgeonnera tout de même à une taille plus petite. START semble donc être un mécanisme imparfait et suggère qu’un ou plusieurs autres mécanismes permettent de contrôler la taille sur le long terme. Soifer et al ont montré que les levures ajoutent un volume constant entre deux bourgeonnements (Soifer, Robert, and Amir 2016; Chandler-Brown et al. 2017). Au niveau moléculaire, l’hypothèse de dilution de Whi5 au cours de la phase G1 est cohérent avec le modèle « Adder » (Soifer, Robert, and Amir 2016; Schmoller et al. 2015).
16
Le mécanisme « Adder » serait donc conservé chez les bactéries, les archéobactéries et les eucaryotes.
Schizosaccharomyces pombe
S. pombe se divise de façon symétrique. La progression du cycle cellulaire est régulée par Cdc2, la seule Cdk retrouvée chez cette levure. Cdc2 contrôle l’entrée en mitose et contrôle la longueur de la phase G2 (Nurse 1990). Contrairement à S. cerevisiae, la régulation de la taille se fait majoritairement en phase G2/M (Jorgensen and Tyers 2004).
La taille serait contrôlée par la protéine Pom1, qui est localisée aux extrémités de la cellule. Pom1 est un inhibiteur des kinases Cdr1 et Cdr2, qui sont eux-mêmes des inhibiteurs de Wee1. Ce dernier inhibe la Cdk Cdc2 par phosphorylation sur la Tyrosine 15 (Figure 8a) (Gould and Nurse 1989). Cdr1 et Cdr2 localisent au centre de la cellule (Deng and Moseley 2013). Pendant la croissance, la concentration de la protéine Pom1 diminue au centre de la cellule. L’inhibition de Cdr1 et Cdr2 est par conséquent levée et Wee1 est inhibé, ce qui permet l’activation de Cdc2 (Figure 8b) (Martin and Berthelot-Grosjean 2009; Moseley et al. 2009). Cependant, une mutation de la Tyrosine 15 de Cdc2 par un acide aminé non phosphorylable ne perturbe pas la taille cellulaire des levures, suggérant que d’autres mécanismes contrôlent la taille cellulaire (Coudreuse and Nurse 2010).
Une autre hypothèse est que la concentration de Cdr2 augmente proportionnellement à la croissance de la cellule et permet l’inhibition de Wee1 (Pan et al. 2014; Russell and Nurse 1987).
17
Figure 8 - Régulation de la taille chez S. pombe. (Marshall et al. 2012). a) Pom1 inhibe
l’activité de Cdr1 et Cdr2. Wee1 inhibe Cdc2. b) Pendant la croissance cellulaire, la concentration de Pom1 diminue au centre de la cellule et Pom1 ne peut plus inhiber Cdr1 et Cdr2. Cdr1 et Cdr2 inhibent Wee1, ce qui permet l’activation de Cdc2.
Métazoaires
La taille des cellules au sein d’un même tissu est homogène (Ginzberg, Kafri, and Kirschner 2015; Lloyd 2013). La régulation cellulaire est un processus extrêmement important pour former des organes et des organismes de taille physiologique. La taille d’un animal dépend à la fois du nombre de cellules et de la taille des cellules. La différence de taille des organismes selon les espèces animales est principalement une différence de nombre de cellules plutôt que de la taille des cellules. Par exemple, l’Homme possède 3 000 fois plus de cellules qu’une souris (Conlon and Raff 1999). Nous avons vu que la régulation de la taille est influencée par l’environnement extérieur. Chez les métazoaires, l’environnement physique peut également contrôler la taille cellulaire. Par exemple, les neurones arrêtent de grandir quand ils touchent leurs cibles (Guthrie 2007). La croissance et la prolifération sont différentiellement contrôlées selon le
18
type de cellule et de tissus, plusieurs niveaux de contrôles sont donc nécessaires. De plus, la croissance et la division ne sont pas toujours couplées. Par exemple, les neurones peuvent grossir sans se diviser. Au contraire, les ovocytes fécondés peuvent se diviser sans grossir.
L’équivalent de START est appelé le « Restriction Point » chez les métazoaires (Wells 2002; Blagosklonny and Pardee 2002; Zetterberg, Larsson, and Wiman 1995). Cependant, cette coordination semble être principalement régulée par des facteurs extracellulaires plutôt que des facteurs intracellulaires (Conlon et al. 2001; Conlon and Raff 2003). La croissance et la division nécessitent des signaux distincts. Des molécules mitogènes sont nécessaires pour favoriser la division cellulaire et des facteurs de croissances sont nécessaires pour la croissance (Conlon et al. 2001; Rathmell et al. 2000).
La voie de régulation contrôlant la croissance la mieux caractérisée est la voie IGF/PI3K/AKT/mTORC1 (Laplante and Sabatini 2012; Tumaneng, Russell, and Guan 2012).
Mammifères
Chez les mammifères, la croissance est contrôlée par la voie TOR, qui contrôle principalement la traduction via le facteur de transcription C-Myc et la protéine kinase S6K. C-Myc augmente la production des ARN ribosomiques et la taille du nucléole afin de favoriser la synthèse des protéines (Grewal et al. 2005; Saucedo and Edgar 2002; Wang, Dillon, et al. 2011). Myc est considéré comme l’analogue fonctionnel de Sfp1 (Cook and Tyers 2007). Les facteurs extracellulaires, comme les facteurs de croissances et les molécules mitogènes semblent être importants pour la régulation de la taille (Conlon and Raff 2003). Des criblages chez l’Homme ont permis d’identifier différents régulateurs de la taille, comme le gène PRR16/Largen, qui est un activateur de la traduction et contrôle l’activité mitochondriale (Yamamoto et al. 2014). Un autre criblage a permis
19
d’identifier la p38 comme un régulateur de la taille chez l’Homme (Liu et al. 2018).
Drosophile
Chez la drosophile, l’inactivation de mTOR par la rapamycine provoque un délai du cycle cellulaire et une diminution de la taille cellulaire (Zhang et al. 2000). S6K (l’analogue fonctionnel de Sch9) est un substrat de mTOR dont l’inactivation génétique ou l’inactivation par traitement à la rapamycine provoque des ailes de petites tailles (Montagne et al. 1999; Chung et al. 1992). mTOR contrôle la croissance cellulaire via la traduction et la biogénèse des ribosomes et augmente également la production de nucléotides afin de favoriser la production d’ARN (ribosomiques et messagers) ainsi que la traduction et produire des dNTP nécessaire à la réplication de l’ADN.
Les nutriments sont également importants pour la régulation de la taille. Une carence nutritive peut provoquer une diminution de la taille des ailes de 15% (Edgar 2006) alors que des nutriments en excès n’influence par la taille des drosophiles.
Nématodes
Les nématodes Caenorhabditis elegans et Ascaris lumbricoides ont été longuement étudiés. Ces deux espèces ont la même taille après l’éclosion, mais à la taille adulte, A. lumbricoides est 109 fois plus gros que C. elegans. La différence entre C. elegans et A. lumbricoides est que ce dernier produit des cellules de plus grande taille afin de produire un organisme allant jusqu’à 40 cm de long.
La taille cellulaire peut influencer la taille des organes ainsi que la taille de l’organisme. Par exemple, le nombre de cellules de C. elegans est fixe, mais des perturbations peuvent modifier la taille des cellules et ainsi modifier la
20
taille de l’organisme adulte (Irle and Schierenberg 2002; Cook and Tyers 2007). Différents criblages pour identifier les gènes régulant la taille ont permis d’identifier les voies TGF-β et MAPK sma-5 (Small Body Size). Les mutations dans ces voies peuvent produire des vers 10 fois plus petits qu’un ver sauvage (Savage-Dunn et al. 2003; Watanabe, Ishihara, and Ohshima 2007). La voie TGF-β semble également avoir un rôle dans la croissance des cellules en favorisant l’augmentation de la ploïdie et a pour conséquence d’augmenter la taille de l’organisme (Lozano et al. 2006).
Régulation de la taille et virulence fongique
De nombreux champignons sont pathogènes pour l’Homme. Les plus importants sont Candida, Aspergillus, Cryptococcus et Histoplasma. Il n’est pas encore établi s’il y a un lien direct entre la régulation de la taille des champignons et la virulence. Cependant, plusieurs études suggèrent que la taille d’un pathogène fongique est un déterminant important de la virulence.
Cryptococcus neoformans
En 2009, une forme « géante » de C. neoformans a été mise en évidence dans un modèle d’infection de souris (Figure 9) (Zaragoza et al. 2010). Ces cellules géantes possèdent une capsule et la cellule elle-même (sans la capsule) a aussi une grande taille. Ces cellules sont plus résistantes au stress osmotique et aux radiations gamma et les macrophages ne peuvent pas les phagocyter (Okagaki et al. 2010). La grande taille et la capsulation sont favorisées par l’environnement de l’hôte, notamment la température à 37°C (Garcia-Rodas et al. 2011). La diversité des cellules (avec et sans capsule) permet l’invasion de l’hôte et l’échappement au système immunitaire. Ces cellules ont également une ploïdie élevée, indiquant qu’elles dupliquent leur génome sans se diviser.
21
Figure 9 - Diversité de la taille de Cryptococcus neoformans. A. Cellules sans capsule. B.
Cellules avec capsule. (Zaragoza et al. 2010) Histoplasma capsulatum
H. capsulatum est une levure pathogène dimorphique, mais contrairement à C. albicans, cette levure produit des hyphes à 25°C et elle se propage sous forme levure à 37°C à l’intérieure de l’Homme (Wang and Lin 2012). À 37°C, ce champignon produit deux types de propagules, des microconidies et des macroconidies. Les microconidies, qui sont les formes infectieuses, ont une petite taille qui leur confère l’avantage de se loger dans les alvéoles pulmonaires et de persister dans les cellules phagocytaires (Seider et al. 2010). Ce champignon produit donc des cellules de différentes formes et taille suivant l’environnement.
Paracoccidioides brasiliensis
Cette levure est caractéristique par sa diversité de forme et de taille au sein d’une population. Une mutation du gène CDC42 provoque une diminution de la taille et de sa variabilité. Ce mutant est plus facilement phagocyté par les macrophages et il est non virulent dans des modèles d’infection de souris. Ceci suggère que la diversité de la taille, ainsi que la grande taille de cette levure sont des attributs importants pour le pouvoir pathogène de ce champignon (Almeida et al. 2009). Cependant, il n’est pas exclu que CDC42 régule d’autres processus nécessaire pour la virulence pendant l’infection.
22 Mucor circinelloides
Mucor circinelloides présente une diversité de taille qui dépend du groupe de compatibilité sexuelle de la cellule (+ ou -). Les cellules produites par des souches (–) sont plus grandes et peuvent provoquer la lyse des cellules de l’hôte à l’inverse des cellules (+) qui ont une petite taille et sont non-pathogènes (Li et al. 2011).
23
C. albicans et les candidoses
C. albicans est une levure de la division des Ascomycètes et de l’ordre des Saccharomycetales. Le nom de classification a été proposé par Christine Marie Berkhout dans sa thèse de doctorat de l’Université d’Utrecht, aux Pays-Bas, en 1923 (Barnett 2004). Candida provient du mot latin « candidus », signifiant « blanc ». Albicans est le participe présent du mot latin « albicō », signifiant « blanc » également.
C. albicans est une levure opportuniste retrouvée chez l’Homme et différents animaux comme les oiseaux, les bovins, les chevaux, les chats (Edelmann, Kruger, and Schmid 2005)… La levure fait partie de la flore commensale humaine de la bouche, du tractus digestif et du vagin. C. albicans est maintenu sous sa forme commensale par d’autres microorganismes et par le système immunitaire de l’hôte. En cas de perturbation de la flore commensale, par prise d’antibiotiques ou d’immunosuppresseurs, à cause du tabagisme (Akram et al. 2018), ou suite à une immunodéficience (SIDA, chimiothérapie), la levure peut devenir virulente et provoquer une candidose. C. albicans est un pathogène très polyvalent et est devenu un agent pathogène majeur. En Amérique du Nord, 10% des septicémies sont causées par des levures du genre Candida, généralement par l’espèce C. albicans (C. albicans représente 90% des septicémies à levure) (Marchetti et al. 2004; Edmond et al. 1999; Bille, Marchetti, and Calandra 2005). Les Candida sont la troisième cause d’infection nosocomiale aux États-Unis (Wisplinghoff et al. 2004), après Escherichia coli et Staphylococcus aureus.
Quelques espèces de Candida, dont C. albicans, utilisent un code génétique non standard. En effet, le codon CUG code généralement pour une leucine mais code pour une sérine chez certaines espèces de Candida
24
(Santos et al. 1997), d’où le nom du clade CTG de C. albicans et les espèces proches (Figure 10).
Figure 10 - Phylogénie des clades Candida et Saccharomyces. Le Clade Candida est
caractérisé par l’utilisation du codon CTG qui code pour une sérine. S. cerevisiae se trouve dans le clade WGD = Whole Genome Duplication. (Butler et al. 2009)
Épidémiologie et manifestations cliniques des candidoses
Parmi les 200 espèces du genre Candida, 14 sont pathogènes pour l’Homme : C. albicans, C. auris, C. dubliensis, C. glabrata, C. kefyr, C. krusei, C. lusitaniae, C. parapsilosis, C. tropicalis, C. guilliermondii, C. famata, C. lipolytica, C norvengensis, C. rugosa (Lopez-Martinez 2010; Satoh et al. 2009; Pfaller et al. 2006). C. albicans est l’espèce la plus fréquemment isolée.
25
Les candidoses sont en augmentation partout dans le monde. Elles sont favorisées par l’âge, la prise d’antibiotiques, de stéroïdes et d’antidépresseurs, par les transplantations d’organe et de moelle osseuse, par le diabète, divers cancers, le SIDA et la malnutrition (Lopez-Martinez 2010).
Les candidoses peuvent avoir différentes formes :
-Candidose buccale, ou muguet, caractérisée par des plaques blanchâtres sur la langue, le palais, les joues et le pharynx. Cette infection touche jusqu’à 90% des patients atteints du SIDA, dans ce cas, l’infection peut se propager dans le système digestif. C’est la forme la plus fréquente de candidose et peut être très agressive pour les prématurés, les femmes allaitantes et les personnes âgées. (Lalla, Patton, and Dongari-Bagtzoglou 2013). Les autres facteurs de risque sont le cancer, l’utilisation d’une prothèse dentaire ou la prise d’antibiotiques.
-Candidose vulvovaginal, caractérisée par une leucorrhée et des démangeaisons. 50 à 75% des femmes ont au moins une candidose vaginale au cours de leur vie, à tout âge (Sobel 2007). Ce type d’infection est favorisé par la prise de contraceptifs ou d’antibiotiques, par l’obésité, chez les femmes enceintes et par les thérapies hormonales (Fidel 2004). -Candidose balano-préputial, caractérisée par une douleur, des pustules, une irritation, parfois par un ulcère et une sécrétion sur le gland du pénis et sur le prépuce. Ce type d’infection est favorisé par le manque d’hygiène, le diabète et l’immunodéficience.
-Onychomycose, caractérisée par une décoloration de l’ongle, ou au contraire une couleur verte-jaune, une séparation à l’extrémité de l’ongle, l’apparition d’un œdème et une douleur. C’est une infection fréquente chez les diabétiques.
-Candidose mucocutanée, caractérisée par une douleur sur la surface infectée, une hyperkératose et des ulcères de la peau. Cette infection peut
26
se propager dans les tissus profonds puis provoquer une septicémie et le décès.
-Candidose invasive, caractérisée par une septicémie, une endocardite, une méningite ou une endophtalmie (Pappas 2006). La septicémie à Candida est la 4ème infection du sang la plus courante. Elle est responsable de 50 000 décès par an dans le monde. Les Candidoses invasives sont favorisées par la prise d’antibiotique, la chirurgie et l’immunodéficience. Traitements des candidoses
Le traitement d’une candidose dépend de la zone d’infection. Un guide est publié pour aider les cliniciens à choisir le traitement approprié. (Pappas et al. 2016; Lopez-Martinez 2010). Dans un premier temps, il est nécessaire d’éliminer les facteurs de prédisposition : arrêter la prise d’antibiotiques, de stéroïdes ou d’immunosuppresseurs ; traiter l’humidité localement ou ajuster le pH vaginal. La prise de probiotiques peut également être efficace dans le cas d’une infection vaginale (Jurden et al. 2012; Abad and Safdar 2009). En revanche, quand le facteur de prédisposition est une pathologie, comme le SIDA, le diabète ou bien un cancer, la prise d’antifongique est nécessaire.
Il existe trois principales classes d’antifongiques utilisées contre les candidoses : les polyènes, les azoles et les échinochandines.
Les polyènes et les azoles sont utilisés pour les traitements oraux, vaginaux, cutanés et en intraveineuse pour les infections systémiques (Moosa et al. 2004; Silverman, Pories, and Caro 1989).
Les échinochandines sont utilisées en intraveineuses et sont recommandées pour le traitement des candidoses systémiques (Bassetti et al. 2018; Pappas et al. 2016).
Ces antifongiques ont plusieurs limitations. Ils peuvent provoquer des effets secondaires, comme des allergies, des insuffisances rénales, des nausées et des maux de tête (Sawant and Khan 2017). De plus, des
27
résistances à ces antifongiques apparaissent. Il est donc nécessaire de trouver de nouvelles molécules pour traiter les candidoses (Perfect 2017).
Génome
C. albicans est un diploïde et possède 8 chromosomes allant de 3,3 à 0,95 Mb (chromosomes 1 à 7 et le chromosome R) (Olaiya and Sogin 1979). Un premier séquençage a été achevé en 2004 (Jones et al. 2004). Un haut niveau d’hétérozygotie, de recombinaison intrachromosomique et d’aneuploïdie a été observé dans différentes souches. Cette plasticité génomique a un effet sur l’adaptation de la levure à différents stress, sur la résistance aux antifongiques (Wertheimer, Stone, and Berman 2016) et génère de la variation phénotypique (Holmes et al. 2006). La fréquence de perte d’hétérozygotie et d’aneuploïdie est 1 000 fois plus élevée in vivo qu’in vitro et l’instabilité génomique a un impact sur la virulence de cette levure (Satpati et al. 2017; Forche et al. 2018). En effet, l’instabilité génétique influence le taux de croissance de la levure, sa morphologie, la résistance aux stress ou aux antifongiques (Braunsdorf and LeibundGut-Landmann 2018; Schonherr et al. 2017).
La taille du génome est d’environ 29Mb (forme diploïde) et le génome possède environ 6 200 ORF (6 600 ORF pour S. cerevisiae), dont 4 400 (70%) ne sont pas caractérisés (http://www.candidagenome.org ; consulté en novembre 2018). S. cerevisiae et C. albicans ont environ 75 % de gènes orthologues et 20% des gènes de C albicans n’ont pas d’homologue chez S. cerevisiae, C. albicans et l’Homme (Odds, Brown, and Gow 2004; Jones et al. 2004). Bien que la levure ait été considérée comme asexuée, le génome contient les gènes permettant une reproduction sexuée (Hull and Johnson 1999). Les levures peuvent former des formes tétraploïdes et revenir sous forme diploïde par perte aléatoire de chromosomes excédentaires (Miller
28
and Johnson 2002; Wu et al. 2005). La méiose n’a jamais été observée chez cette levure (Bennett and Johnson 2005).
Morphologie
C. albicans est une levure pléomorphique, c’est-à-dire qu’elle présente différentes morphologies selon l’environnement dans laquelle elle se trouve (Figure 11). C. albicans a la possibilité de passer d’une forme levure à une forme hyphe ou pseudohyphe. De plus, la levure peut faire des « transitions phénotypiques » nommées White, Opaque, GUT (Gastrointestinally induced Transition) et Gray. Récemment, la forme Goliath a été décrite, cette forme est observée dans des milieux carencés en zinc (Malavia et al. 2017).
Figure 11 - Représentation des différentes formes morphologiques de C. albicans (Gow
and Yadav 2017).
29
C. albicans est capable de se diviser sous forme levure ou bien sous forme filamenteuse (hyphe ou pseudohyphe) (Figures 11 et 12). La forme levure prolifère par bourgeonnement avec une forme ellipsoïdale. La forme filamenteuse croît par élongation, la cellule fille ne se sépare pas de la cellule mère et forme des filaments. Dans le cas d’un pseudo-hyphe, il y a formation d’une paroi transversale appelée septum. La croissance des hyphes est assurée par le Spitzenkörper (Berman 2006), un complexe dans la région apicale de l’hyphe qui est riche en vésicules sécrétoires et permettant la polymérisation des constituants de la paroi. La forme levure est favorisée par un pH acide et une température inférieure à 25°C. La forme hyphe est favorisée par un pH alcalin, une température de 37°C, une haute concentration en CO2, une carence nutritive, l’hypoxie, la croissance sur une surface solide et la présence de sérum dans le milieu. La forme pseudo-hyphe est favorisée à 35°C, pH=6 et en carence azotée. Le dimorphisme levure/hyphe semble nécessaire pour la virulence. De nombreux mutants incapables de faire la transition entre les formes levures et hyphes perdent leur capacité à coloniser et envahir l’hôte, suggérant que les deux formes sont nécessaires pour la virulence (Lo et al. 1997; Saville et al. 2003). La forme levure serait nécessaire pour la colonisation et la dissémination (Saville et al. 2003) alors que la forme hyphe serait nécessaire pour l’invasion des tissus et l’échappement au système immunitaire (Berman and Sudbery 2002; Malinverni et al. 1985).
30
Figure 12 - Photos de C. albicans en forme pseudohyphe (Pseudohyphae), levure (Yeast)
et hyphe (Hyphae). La barre d’échelle représente 5 µm (Sudbery 2011).
Transitions phénotypiques
C. albicans est capable d’exprimer des phénotypes différents suivant les conditions environnementales. La transition phénotypique est définie comme la capacité de subir spontanément et de manière réversible des transitions de morphologie (Soll 1992). Certains phénotypes semblent être les formes commensales et d’autres les formes pathogènes. Quatre formes phénotypiques ont été décrites chez C. albicans : White, Gray, Opaque et GUT (Gastrointestinally-IndUced Transition).
Dans les conditions de laboratoire, la forme la plus commune est la forme White. Les colonies White sont blanches, lisses et rondes. La transition White-Opaque a été décrite en 1985 (Slutsky et al. 1987). Les colonies Opaques sont plus grosses, plus rugueuses et plus grises que les formes White. Les cellules Opaques sont compétentes pour la reproduction