Saurons-nous construire une bactérie synthétique ?
Texte intégral
Figure
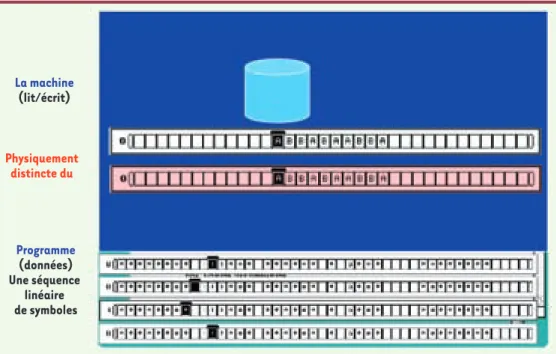
![Figure 3. Le groupement des gènes « persistants » en clusters assure la survie de la cellule [10].](https://thumb-eu.123doks.com/thumbv2/123doknet/12239223.319055/5.891.85.553.83.374/figure-groupement-gènes-persistants-clusters-assure-survie-cellule.webp)
Documents relatifs
Pour permettre à chacune et à chacun de prendre une réelle place dans le développement de son territoire, ID innove pour assurer une participation citoyenne la plus
[r]
après libération ,les toxines vont diffuser dans l’organisme et venir agir sur les cellules- cible elles peuvent donc agir à distance du foyer infectieux où elles sont
En ouvrant largement ses pinces philosophiques comme pour mieux saisir l'esprit de l'opération en cours, le Géomètre tel un "Crabe terrestre" au regard tout
Ces compétences, même si nous avons du mal à nous les recon- naître, il est nécessaire de les transmettre à de nouveaux venus à la pédagogie Freinet et à bien
Toutes les équations précédentes, jointes à l'hypothèse émise concernant la l i néarité, permettent de construire un hydrogramme synthétique résultant d'une pluie nette de
Le triangle qui a été ainsi construit est un triangle rectangle, et cela est vrai pour tous les triangles qui sont faits avec
Etayer les compétences managériales et notamment dans le domaine du management participatif... Plan de
