MicroARN et translocations chromosomiques dans les hémopathies malignes
Texte intégral
Figure

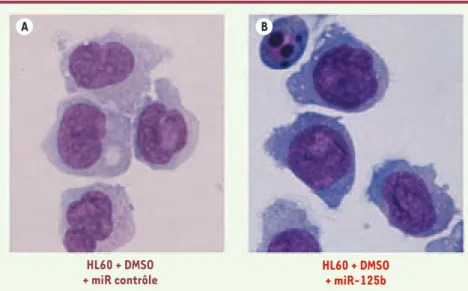
Documents relatifs
Ce sont des proliférations de petites cellules B avec souvent une différentiation plasmocytaire. Leur évolution est
L’archive ouverte pluridisciplinaire HAL, est destinée au dépôt et à la diffusion de documents scientifiques de niveau recherche, publiés ou non, émanant des
Although the catalytic mechanism of MtNAS turned out to be quite different from both those of spermine and spermidine synthase, the E81Q mutant was nevertheless free of tNA in
Dans les anomalies chromosomiques des néoplasies de type T, 1 'examen des chromosomes remaniés confirme que les points de cassure localisés dans les bandes
which may result from either incomplete pairing in the interstitial regions of the quadrivalent at early pachytene or desynapsis at mid-late pachytene starting in these
Le mécanisme de la trans location serait donc une recombinaison aberrante, probablement catalysée par la recombz'nase, entre des segments situés sur des chromosomes
Dans la dernière partie nous avons mis en œuvre des modèles de niche pour prédire l’impact potentiel des changements globaux sur la distribution des deux
Les translocations réciproques sont dues à des échanges de segments chromosomiques entre deux chromosomes, les points de cassure s'étant produits ailleurs que dans les
