HAL Id: hal-02806696
https://hal.inrae.fr/hal-02806696
Submitted on 6 Jun 2020HAL is a multi-disciplinary open access
archive for the deposit and dissemination of sci-entific research documents, whether they are pub-lished or not. The documents may come from teaching and research institutions in France or abroad, or from public or private research centers.
L’archive ouverte pluridisciplinaire HAL, est destinée au dépôt et à la diffusion de documents scientifiques de niveau recherche, publiés ou non, émanant des établissements d’enseignement et de recherche français ou étrangers, des laboratoires publics ou privés.
Détermination des domaines vitaux et taux de contact
chez le blaireau, Meles meles en présence de zones à
bovins dans un contexte de tuberculose bovine.
Lucile Millet
To cite this version:
Lucile Millet. Détermination des domaines vitaux et taux de contact chez le blaireau, Meles meles en présence de zones à bovins dans un contexte de tuberculose bovine.. [Stage] Université de Bourgogne (UB), FRA. 2012, 52 p. �hal-02806696�
MEMOIRE DE STAGE
Détermination des domaines vitaux et taux
de contact chez le blaireau, Meles meles en
présence de zones à bovins dans un contexte
de tuberculose bovine.
Maîtres de stage:
Ariane PAYNE
Loïc BOLLACHE
Tuteur pédagogique:
Lucile MILLET
Paul ALIBERT 2011-2012
Remerciements :
Je tiens à remercier Ariane Payne, qui m’a permis de réaliser ce stage enrichissant et qui a toujours été rassurante et de très bon conseil.
Je tiens à remercier Loïc Bollache pour son entrain sans limite à travers le stage. Merci à l’ONCFS de m’avoir accueilli au sein de leur organisme.
Organisme d’accueil
:
L’Office National de la Chasse et de la Faune Sauvage est un établissement public sous la double tutelle des Ministères chargés du Développement Durable et de l’Agriculture. Elle remplit cinq missions principales qui s’inscrivent dans les objectifs gouvernementaux du Grenelle de l’Environnement.
Créé en 1972, l’Office dispose d’un budget de 120 millions d’euros pour remplir ses missions sur l’ensemble du territoire (métropole et DOM).
Créée en 2001, la Délégation Interrégionale Bourgogne Franche-Comté se compose de deux régions administratives, la Bourgogne et la Franche-Comté, et de huit départements : la Côte d’Or (21), la Nièvre (58), l’Yonne (89), la Saône-et-Loire (71), pour la Bourgogne ; le Doubs (25), le Jura (39), la Haute-Saône (70) et le Territoire de Belfort (90) pour la Franche-Comté. Cette entité couvre une superficie totale de 47 784 km² (soit près de 8,8 % du territoire national), dont 31 582 km² pour la Bourgogne.
La Délégation regroupe environ une centaine d’agents techniques et techniciens de l’Environnement affectés dans les 6 Services Départementaux, le Service Inter-Départemental et la Brigade Mobile d’Intervention (BMI), et six agents basés au siège de la Délégation à Dijon (Délégué Régional et son adjoint, deux Ingénieurs en Cellule Technique, un correspondant informatique et une secrétaire). La délégation de Dijon se situe au 57 rue de Mulhouse.
Sommaire :
PARTIE I : Introduction Bibliographique ... 1
1. Contexte ... 1
2. Biologie du blaireau Meles meles ... 2
3. Domaines vitaux-Généralités ... 3
a. Définition ... 3
b. Méthodes d’estimation des domaines vitaux ... 5
4. Utilisation de l’habitat et Taux de contact ... 6
a. Utilisation de l’habitat ... 6
b. Taux de contact ... 7
5. Télémétrie ... 8
PARTIE II : Matériel et Méthode... 9
1. Sites d’étude ... 9
2. Capture et manipulation des blaireaux ... 10
3. Acquisition des données de localisation ... 11
4. Télémétrie ... 12
5. Estimation des domaines vitaux ... 12
6. Estimation de l’utilisation de l’habitat et taux de contact ... 13
7. Analyses statistiques ... 14
PARTIE III : Résultats et Critiques ... 14
1. Résultats ... 14 a. Suivi télémétrique ... 14 b. Domaines Vitaux ... 20 c. Utilisation de l’habitat ... 27 d. Taux de contact ... 30 2. Discussion et critiques ... 31 Liste bibliographique: ... 36 ANNEXES : ... 42
Liste des figures :
Figure 1 : Carte représentative de la Côte d’Or et les différentes vallées où ont été effectués les captures (*représentent les endroits précis des captures)
Figure 2 : Représentation de l’utilisation de chaque type d’habitat pour chaque blaireau au sein de l’ensemble de leur domaine vital
Figure 3 : Représentation de l’utilisation de chaque type d’habitat pour chaque blaireau au sein du domaine vital à 50% d’UD pour chaque blaireau
Figure 4 : Représentation de l’utilisation de chaque type d’habitat pour chaque blaireau au sein du domaine vital à 30% d’UD pour chaque blaireau
Figure 5 : Représentation de l’utilisation de chaque type d’habitat en fonction de chaque saison chez le blaireau Alhambra
Figure 6 : Représentation de la corrélation entre l’utilisation des pâtures et les distances terriers-pâtures
Figure 7 : Représentation du pourcentage d’utilisation de la lisière en pâture et des bosquets en pâture contre le pourcentage d’utilisation de la pâture exclusivement
Liste des images :
Image 1 : Représentation du domaine vital à 95% d’UD avec une localisation toutes les demi-heures (en rouge) et une localisation toutes les deux demi-heures (en bleu) chez Adélie à L’Oiserolle
Image 2 : Représentation du domaine vital à 95% d’UD avec une localisation toutes les demi-heures (en rouge) et une localisation toutes les deux demi-heures (en bleu) chez Alhambra à La Roche Vanneau
Image 3 : Représentation du domaine vital à 95% d’UD avec une localisation toutes les demi-heures (en rouge) et une localisation toutes les deux demi-heures (en bleu) chez Brownette à Braux Image 4 : Représentation du domaine vital à 95% d’UD avec une localisation toutes les demi-heures (en rouge) et une localisation toutes les deux demi-heures (en bleu) chez Brownie à Braux Image 5 : Représentation du domaine vital à 95% d’UD avec une localisation toutes les demi-heures (en rouge) et une localisation toutes les deux demi-heures (en bleu) chez Eddy à La Roche Vanneau
Image 6 : Représentation du domaine vital à 95% d’UD avec une localisation toutes les demi-heures (en rouge) et une localisation toutes les deux demi-heures (en bleu) chez Flavie à Grenant Image 7 : Représentation du domaine vital à 95% d’UD avec une localisation toutes les demi-heures (en rouge) et une localisation toutes les deux demi-heures (en bleu) chez Johnny à Grenant Image 8 : Représentation du domaine vital à 95% d’UD avec une localisation toutes les demi-heures (en rouge) et une localisation toutes les deux demi-heures (en bleu) chez Galou à Jaugey Image 9 : Représentation des domaines vitaux calculés avec la méthode du kernel fixe LSCV (en rouge) à 95% d’UD et du MCP (en bleu turquoise) (a) et ceux calculés avec la méthode du kernel fixe LSCV à 50% et 30% d’UD (b) de la blairelle Adélie avec ses localisations (*) et son terrier ( )
Image 10 : Représentation des domaines vitaux calculés avec la méthode du kernel fixe LSCV (en rouge) à 95% d’UD et du MCP (en bleu turquoise) (a) et ceux calculés avec la méthode du kernel fixe LSCV à 50% et 30% d’UD (b) de la blairelle Alhambra avec ses localisations (*) et son terrier ( )
Image 11 : Représentation des domaines vitaux calculés avec la méthode du kernel fixe LSCV (en rouge) à 95% d’UD et du MCP (en bleu turquoise) (a) et ceux calculés avec la méthode du kernel fixe LSCV à 50% et 30% d’UD (b) de la blairelle Brownette avec ses localisations (*) et son terrier ( )
Image 12 : Représentation des domaines vitaux calculés avec la méthode du kernel fixe LSCV (en rouge) à 95% d’UD et du MCP (en bleu turquoise) (a) et ceux calculés avec la méthode du kernel fixe LSCV à 50% et 30% d’UD (b) du blaireau Brownie avec ses localisations (*) et son terrier ( )
Image 13 : Représentation des domaines vitaux calculés avec la méthode du kernel fixe LSCV (en rouge) à 95% d’UD et du MCP (en bleu turquoise) (a) et ceux calculés avec la méthode du kernel fixe LSCV à 50% et 30% d’UD (b) du blaireau Eddy avec ses localisations (*) et son terrier ( )
Image 14 : Représentation des domaines vitaux calculés avec la méthode du kernel fixe LSCV (en rouge) à 95% d’UD et du MCP (en bleu turquoise) (a) et ceux calculés avec la méthode du kernel fixe LSCV à 50% et 30% d’UD (b) de la blairelle Flavie avec ses localisations (*) et son terrier ( )
Image 15 : Représentation des domaines vitaux calculés avec la méthode du kernel fixe LSCV (en rouge) à 95% d’UD et du MCP (en bleu turquoise) (a) et ceux calculés avec la méthode du kernel fixe LSCV à 50% et 30% d’UD (b) du blaireau Johnny avec ses localisations (*) et son terrier ( )
Image 16 : Représentation des domaines vitaux calculés avec la méthode du kernel fixe LSCV (en rouge) à 95% d’UD et du MCP (en bleu turquoise) (a) et ceux calculés avec la méthode du kernel fixe LSCV à 50% et 30% d’UD (b) du blaireau Galou avec ses localisations (*) et son terrier ( )
Image 17 : Représentation des domaines vitaux d’Alhambra calculés avec la méthode du kernel fixe LSCV pour chaque saison (en blanc pour l’hiver, en rose pour le printemps, en jaune pour l’été et en bleu pour l’automne).
Image 18 : Représentation des domaines vitaux de Galou calculés avec la méthode du kernel fixe LSCV pour chaque saison (en blanc pour l’hiver, en rose pour le printemps, en jaune pour l’été et en bleu pour l’automne).
Liste des tableaux :
Tableau 1 : Récapitulatif des méthodes d’estimation des domaines vitaux (Pellerin, 2005) Tableau 2 : Variables individuelles de chaque blaireau capturé
Tableau 3 : Récapitulatif des distances choisies pour les différents types de milieu Tableau 4 : Représentation des localisations pour chaque individu
Tableau 5 : Représentation des données non exploitables à l’analyse sur les données totales Tableau 6 : Représentation des localisations exploitables et non exploitables (en pourcentage) en fonction du milieu (ouvert et fermé)
Tableau 7 : Représentation de la taille des domaines vitaux de chaque blaireau calculés avec des méthodes différentes et les moyennes correspondantes
Tableau 8 : Représentation du pourcentage des aires d’intense activité à 50% d’UD sur la superficie totale du domaine vital
Tableau 9 : Représentation de la taille des domaines vitaux correspondant aux saisons des deux blaireaux Alhambra et Galou
Tableau 10 : Représentation du pourcentage d’utilisation de la pâture et des bâtiments accueillant des cheptels bovins pour chaque blaireau
Liste des annexes :
Annexe 1 : Exemple d’une feuille Excel provenant d’un collier GPS
Annexe 2 : Résultats concernant les distances choisies pour les polygones sous logiciel MapInfo
1
PARTIE I : Introduction Bibliographique
1. Contexte
Le caractère généraliste des pathogènes contagieux pour l’homme et leurs bétails les rend capable d’infecter de multiples espèces d’hôte. 77 % des pathogènes du bétail infectent plus d’une espèce hôte (Cleaveland et al., 2001). Quand la maladie existe dans des systèmes d’hôtes multiples, on parle de transmission inter-spécifique de la maladie (Holt et al., 2003 ; Fenton & Pedersen, 2005).
C’est notamment le cas dans la transmission de la tuberculose bovine (ANSES, 2011). Cette maladie, causée par le bacille gram + acido-alcoolique Mycobacterium bovis appartenant au complexe Mycobacterium tuberculosis, est très contagieuse pour la faune et l’homme. Dans la plupart des cas, la maladie est chronique et d’évolution lente. Chez certaines espèces elle peut être asymptômatique. Les bovins sont considérés comme l’hôte véritable et originel de M. bovis. La maladie a aussi été signalée chez beaucoup d’animaux domestiques et sauvages. A l’origine, la tuberculose bovine est transmise aux animaux sauvages par des bovins infectés. Néanmoins, la bactérie à l’origine de l’infection peut chez certaines espèces et dans certaines conditions démographiques et environnementales se développer et se maintenir dans les populations sauvages, qui peuvent alors constituer un réservoir primaire, source d’infection pour les bovins et/ou l’Homme (Hars et al., 2011).
La transmission de la tuberculose bovine par la faune sauvage au bétail est présente dans de nombreuses zones agricoles à travers le monde (Porphyre et al., 2008 ; Reilly & Courtenay, 2008). Par exemple, en Afrique, le buffle (Synceros caffer) constitue un réservoir primaire de la tuberculose bovine et en Nouvelle-Zélande, c’est l’opossum (Trichosurus
vulpecula). Malgré un siècle de recherche, la voie précise de cette transmission reste floue
pour de nombreuses espèces (Briscoe, 1912 ; Phillips et al., 2003). Ce problème se pose notamment au Royaume-Uni et en Irlande où les populations de blaireaux (Meles meles) sont connus pour être réservoirs primaires de M. bovis, capables d’entretenir l ‘infection en l’absence d’autres hôtes et de transmettre la maladie aux bovins (Krebs et al., 1997).
En France, les premiers blaireaux porteurs de la bactérie ont été découverts en 2009 en Côte d’Or à proximité de cheptels bovins récemment infectés (Hars et al., 2010). Des mesures d’éradication et des contrôles réguliers existaient depuis 1995 dans les cheptels bovins et la France a acquis le statut indemne de tuberculose en 2001 (Comité OMS, 1954). Cependant,
2 lorsque des blaireaux infectés par M. bovis ont été trouvés, une enquête destinée à connaître la prévalence de la tuberculose chez le blaireau a été mise en place dans le département par les services vétérinaires (Direction Départementale de la Protection des Populations de Côte d’Or, 2012). La prévalence chez les blaireaux entre 2009 et 2011 a été évaluée à 6%. Les voies précises de transmission et la mesure dans laquelle les blaireaux contribuent à l’infection des bovins sont cependant méconnues (Hutchings & Harris, 1997 ; Garnett et al., 2002 ; Kaneene et al., 2002). Deux grands axes ont été proposés dans la littérature comme moyen d’infection entre la faune et le bétail : une transmission indirecte par le biais des pâturages partagés, du foin, du grain, ou de l’ensilage contaminés par la salive, l’urine ou les matières fécales (Hutchings & Harris, 1997) ou une transmission directe par les aérosols (Garnett et al., 2002).
A partir de 2011, un programme de recherche a été mis en place par l’unité sanitaire de la faune de la direction des études et de la recherche de l’Office Nationale de la Chasse et de la Faune Sauvage. L’étude s’intègre dans un travail de thèse qui a pour buts de déterminer les conditions d’émergence de la tuberculose bovine dans la faune sauvage et d’évaluer le risque de transmission entre les populations sauvages (ciblant les blaireaux et les sangliers) et les bovins. Dans ce rapport, l’objectif est de mieux comprendre les aspects spatio-temporels des interactions entre blaireaux et bovins pouvant conduire au risque de transmission de la tuberculose bovine via l’environnement.
2. Biologie du blaireau Meles meles
Le blaireau Meles meles fait partie de la famille des Mustélidés. Il est présent en Europe sous un climat tempéré sous 2000m d’altitude (Griffiths & Thomas, 1998). C’est une espèce nocturne et crépusculaire. De nuit, son activité se concentre sur la recherche et la prise de nourriture. Le blaireau est omnivore opportuniste, il cueille sa nourriture plus qu’il ne la prédate. Son régime alimentaire est principalement constitué de vers de terre (Lumbricus
terrestris). Il se nourrit aussi de micro-mammifères, de baies sauvages, d’insectes,
d’amphibiens et de céréales (Cleary et al., 2009). La recherche de nourriture s’effectue par la fréquentation de différents types de milieux : ouverts tels que la pâture, les cultures et fermés comme les endroits boisés. De jour, il occupe son terrier profond et complexe qu’il creuse à l’aide de ses pattes courtes et puissantes et de ses longues griffes non rétractiles (Do Linh San, 2002). Leur terrier est généralement situé en lisière de forêt proche des milieux ouverts (Do
3 Linh San, 2002). Le terrier principal accueille les individus tout au long de l’année. Il permet aussi l’accueil des jeunes entre février et mars. Les terriers secondaires servent occasionnellement et ne sont pas occupés à temps complet. Lorsque la densité de blaireaux au km2 est élevée, les territoires des individus sont délimités activement en déposant les défécations et les sécrétions des glandes anales dans des pots nommés « latrines » (Do Linh San, 2002 ; Tuyttens et al., 2001). Lorsque la densité est faible (moins de 2 individus au km2), les latrines sont réparties de manière irrégulière au sein de leur domaine vital (Balestrieri et al., 2011). La taille de leurs domaines vitaux a été évaluée entre 50 et 150 ha (Do Linh San, 2002). L’activité des blaireaux est intense en printemps et en été. C’est à cette période qu’ils s’occupent de l’élevage des jeunes (Kowalczyk et al., 2006). Lorsque les conditions environnementales et météorologiques sont moins favorables (novembre à février), ils hivernent et sortent très peu de leur terriers. En Angleterre, où la densité des blaireaux est élevée, plusieurs constats ont été montrés. Il a été prouvé que la fréquentation des bâtiments par les blaireaux est élevée et que les blaireaux infectés visitent plus les fermes et les bâtiments d’élevage que les blaireaux non infectés (Tolhurst et al., 2009).
3. Domaines vitaux-Généralités
a. Définition
En 1943, Burt fut le premier à donner une définition du domaine vital d’un animal. Il le décrivit comme la zone qu’un animal traverse lors de ses activités normales comme la recherche et le partage de la nourriture, la reproduction et le soin aux jeunes. Burt n’a pas considéré les sorties occasionnelles comme faisant partie du domaine vital d’un animal car elles sont plus de nature exploratoire.
Déterminer la taille, la forme et la structure interne d’un domaine vital, c’est-à-dire la structure spatio-temporelle des activités d’un animal, nécessite de localiser un individu au cours de ses diverses activités. Ces localisations doivent être réalisées sans interférence avec le comportement de l’animal en le localisant à distance grâce à un émetteur (système VHF « Very High Frequency » ou GPS « Global Positioning System »). Les localisations sont réalisées uniquement au cours des phases d’activité de l’animal considéré, le repos n’étant pas considéré comme une activité.
4 Les localisations obtenues par GPS d'un animal correspondent à un échantillon de l’ensemble des emplacements occupés formant son domaine vital. La caractérisation statistique du domaine vital doit donc reposer en principe sur des localisations indépendantes (ou non autocorrélés) (Bodin, 2005). L'autocorrélation entre localisations successives se manifeste lorsque le délai entre deux mesures est insuffisant. Lors du calcul d’un domaine vital, un nombre insuffisant de données autocorrélés engendre un biais sur l’emplacement du centre d'activité et une sous-estimation de la taille du domaine vital. Ce problème disparaît dès que l'échantillon atteint un effectif suffisant.
Les méthodes qui permettent de mesurer les domaines vitaux doivent avoir une signification biologique pour l’animal et être objectives. Pour se faire, lors de l’analyse des données, un estimateur des domaines vitaux doit être utilisé pour vérifier et tester les hypothèses ainsi que les données (Powell, 1987). Le choix de cet estimateur est très important. Les domaines vitaux fournissent de riches données sur l’organisation sociale et les activités associées, et sur toutes les interactions liées comme la sélection d’un habitat par l’animal (Powell, 2000). Cet estimateur devrait pouvoir déterminer avec un certain niveau de prévisibilité où se trouve l’animal dans son domaine vital. En effet, dans les années 70, la définition donnée par Burt a été reformulée comme la probabilité de trouver un animal à une localisation particulière d’un plan (Anderson, 1982). Cette probabilité est une fonction de densité appelée Distribution d’Utilisation (UD) et correspond à la distribution des positions d’un animal dans le plan.
Le domaine vital ne doit pas être limité à une notion de surface, mais être à même de mettre en évidence la distribution des activités de l’animal dans l’espace, c’est-à-dire le pourcentage de temps que l’animal consacre aux différentes zones de son domaine vital. Le domaine vital est caractérisé par des sites d’intense activité entourés de zones moins souvent utilisées. En effet, il est important de savoir si l’animal passe plus de temps dans certaines parties de son domaine. Elles peuvent avoir une signification biologique et/ou être liées à des conditions environnementales. Chez certains mammifères terrestres, jusqu’à la moitié de la surface globale peut correspondre à des zones où l’animal passe moins de 5% de son temps (Benhamou, 1998). Cela signifie que 95% de ses déplacements sont concentrés sur une surface minime. Le domaine vital peut être défini par une aire minimale utilisée par un animal au cours d’une période donnée et contenant un pourcentage donné (généralement 95%) de son
5 activité globale (Jennrich & Turner, 1969). Les zones d’intense activité correspondraient à 50% du temps d’activité de l’animal (Kaufmann, 1962 ; Bodin, 2005).
Ce rapport permettra de comprendre et de déterminer les différentes méthodes d’estimation et de quantification du domaine vital pour mieux répondre à nos objectifs.
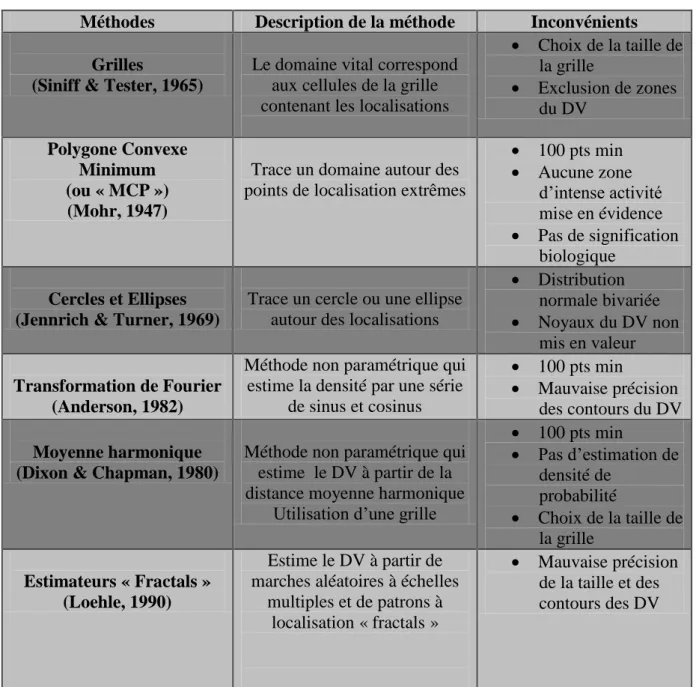
b. Méthodes d’estimation des domaines vitaux
Il existe de nombreuses méthodes pour estimer et mesurer un domaine vital (Powell, 2000).
Méthodes Description de la méthode Inconvénients Grilles
(Siniff & Tester, 1965)
Le domaine vital correspond aux cellules de la grille contenant les localisations
Choix de la taille de la grille Exclusion de zones du DV Polygone Convexe Minimum (ou « MCP ») (Mohr, 1947)
Trace un domaine autour des points de localisation extrêmes
100 pts min Aucune zone d’intense activité mise en évidence Pas de signification biologique Cercles et Ellipses (Jennrich & Turner, 1969)
Trace un cercle ou une ellipse autour des localisations
Distribution normale bivariée Noyaux du DV non mis en valeur Transformation de Fourier (Anderson, 1982)
Méthode non paramétrique qui estime la densité par une série
de sinus et cosinus
100 pts min
Mauvaise précision des contours du DV
Moyenne harmonique (Dixon & Chapman, 1980)
Méthode non paramétrique qui estime le DV à partir de la distance moyenne harmonique
Utilisation d’une grille
100 pts min Pas d’estimation de densité de probabilité Choix de la taille de la grille Estimateurs « Fractals » (Loehle, 1990) Estime le DV à partir de marches aléatoires à échelles
multiples et de patrons à localisation « fractals »
Mauvaise précision de la taille et des contours des DV
6 Il existe une autre méthode, qui n’est pas décrite dans ce tableau, qui permet de pallier les problèmes majeurs de l’estimation des domaines vitaux. C’est la méthode du kernel développée par Worton en 1989. C’est une méthode statistique non paramétrique qui fournit une estimation de densité non biaisée directement à partir des données. Une fonction de densité de probabilité (le kernel) est placée sur chaque point de localisation et l’estimateur est construit en additionnant les différentes composantes. Plus les points sont concentrés, plus la densité de probabilité est forte. Le paramètre de lissage (h) contrôle la quantité de variabilité dans chaque composante de l’estimation. Le choix du paramètre de lissage est très important car il influence fortement la forme et la surface du domaine vital estimé. Cette méthode fait l’hypothèse que tous les points sont indépendants (ou non autocorrélés). Dans le cas de localisations GPS, pour éviter l’autocorrélation spatiale des points, il faut donc des intervalles de temps suffisamment éloignés.
Deux approches ont été développées pour la méthode kernel : le kernel fixe (« fixed kernel ») et le kernel adaptatif (« adaptative kernel ») (Worton, 1989). Le kernel est appelé « fixe » lorsque les paramètres de lissage sont fixés à travers le plan. Au contraire, la méthode adaptative fait varier le paramètre dans le plan pour que les aires avec une faible concentration de points aient une plus grande valeur de h que les aires avec une grande concentration de points.
Dans la méthode du kernel fixe, il existe trois méthodes pour déterminer le paramètre de lissage: le h de référence (« ad hoc ») fixe, le h « Least-Squares Cross-Validation » (LSCV), et le h fixé à une valeur donnée par l’opérateur (Silverman, 1986 ; Worton, 1989). La méthode de kernel fixe avec l’utilisation de la méthode LSCV est celle qui comporte le moins de biais et permet d’obtenir jusqu’à 95% d’UD (Seaman & Powell, 1996). Pour cette méthode, un échantillon de 30 localisations minimum est nécessaire mais il serait préférable d’utiliser au moins 50 localisations par animal (Seaman et al., 1999).
4. Utilisation de l’habitat et Taux de contact
a. Utilisation de l’habitatEn 1993, Block et Brennan ont défini l’habitat comme l’ensemble des facteurs physiques environnementaux qu’une espèce utilise pour sa survie et sa reproduction. Ce sont
7 les ressources et les conditions présentes dans une zone qui permettent l’occupation, la survie et la reproduction, par un organisme donné (Hall et al., 1997).
L’utilisation de l’habitat décrit comment une espèce ou un individu utilise son environnement pour subvenir à ses besoins. Pour mesurer l’utilisation de l’habitat chez une espèce, on étudie les domaines vitaux. Le domaine vital permet d’obtenir la zone d’activité de l’animal. La superposition du domaine vital avec l’environnement nous donne l’habitat utilisé par l’animal. L’environnement peut alors être classé en différentes catégories, choisies en fonction des objectifs de l’étude. L’utilisation de l’habitat correspondra au taux de localisations (ou à la probabilité de présence si on utilise le kernel) pour chaque classe d’habitat considéré.
b. Taux de contact
Définition
Un individu donné peut fréquenter certains milieux plus que d’autres car ils sont attrayants d’un point de vue nourriture, site de reproduction…L’habitat des espèces domestiques peut être attractif pour les espèces sauvages notamment quand elles peuvent y trouver une source de nourriture et d’abreuvement (ex : point d’eau, zone de nourrissage, pierre à sel, pâture, paysages bocagers) (Garnett et al., 2002). Ces lieux favorisent de ce fait les contacts entre les deux espèces et sont d’autant plus nombreux que le paysage est fragmenté (entremêlement de forêts et de pâtures ou cultures). En 2001, Courtenay et al., ont définit le taux de contact ci d’un individu i avec un élément paysager donné (en l’occurrence
100m autour des habitations humaines) comme la somme des localisations d’un individu i trouvées sur cet élément paysager à travers un nombre de villages j divisé par le nombre total de localisations N obtenus par l’individu : Ci
=∑c
ij= (∑
j=1
n ij
)/N.
On distingue alors deux grands types de taux contact. Il y a le taux de contact direct et indirect. Par exemple, en 2006, Richomme et al., ont travaillé sur la transmission de la pasteurellose et de la brucellose entre le bétail et les ongulés sauvages. Ils parlent de taux de contact indirect lorsque l’ongulé sauvage pâture dans une zone utilisé par le bétail. Au contraire, ils parlent de taux de contact direct lorsque les ongulés sauvages se trouvent à une distance de moins de 20m d’un troupeau domestique. Ils ont alors calculé le taux de contact direct et indirect de manière analogue à Courtenay et al., 2012.
8 Dans l’étude, le calcul des domaines vitaux et l’utilisation de l’habitat va fournir des informations sur la présence du blaireau sur les zones de présence des bovins (pâtures et bâtiments d’élevage). La distribution du blaireau dans son environnement va donc permettre de mesurer les interactions entre les deux espèces. Le taux de contact revient finalement à estimer l’utilisation de l’habitat correspondant aux zones de présence des bovins.
Méthodes d’estimation du taux de contact
Pour étudier les possibilités de taux de contact indirect à partir d’un élément paysager, différentes approches sont décrites.
Premièrement, on peut étudier la fréquence de visite de l’élément paysager. On peut le faire sur l’ensemble des localisations d’un animal équipé d’un GPS pour savoir combien de fois il est observé à proximité ou au niveau de l’élément paysager. En 2009, Wickoff et al., ont utilisé cette méthode pour déterminer la fréquence des contacts entre sangliers et porcs domestiques présents dans les fermes. On peut également déterminer quelles variables environnementales ou individuelles influencent la fréquence des visites. Une analyse de ce type est réalisée dans l’article de Tolhurst et al., 2009, sur la fréquentation des fermes de bovins par les blaireaux. Cela lui permet d’étudier la fréquentation de la ferme en fonction du sexe du blaireau et de la saison.
Deuxièmement, on peut calculer le domaine vital de l’animal et le comparer avec l’élément paysager comme les zones de pâture d’animaux domestiques (cartographiée sous SIG). Cette approche est adoptée dans l’article de Clifford et al., 2009.
5. Télémétrie
Il existe de nombreux biais dans les études de télémétrie GPS, qui peuvent être classés en deux grands types d’erreurs (Frair et al., 2004). La première concerne le manque de précision des localisations. Le deuxième type d’erreur concerne les données manquantes. Elles comportent un plus grand biais que le manque de précision parce qu’elles représentent un manque d’information au sein de l’analyse (Johnson et al., 1998). Ces erreurs sont majoritairement dues au degré d’hétérogénéité du paysage (Frair et al., 2004). Des études ont montré que la couverture végétale, la densité et la hauteur des arbres peuvent influencer l’acquisition des localisations (Rumble & Lindzey, 1997). En 1997, Rumple et Rodgers, ont montré qu’un collier GPS est 3.8 fois moins susceptible d’acquérir une localisation en forêt
9 (>15m de hauteur) qu’en zone dépourvue d’arbres. La période de la journée où les localisations sont enregistrées peut être un biais car le satellite change durant la journée (Moen et al., 1997). Frair et al., 2004, ont aussi montré que la saison et le comportement de l’animal jouaient un rôle dans l’acquisition des données. En effet, la météorologie peut entraîner une perte d’information. De plus, si un animal est errant ou explorateur et surtout si le paysage est très fragmenté, il peut traverser sur une courte durée des habitats très variés et donc une couverture végétale très hétérogène. Enfin, l’année de fabrication et la marque du collier utilisé affecte aussi la performance de l’acquisition des données télémétriques (Rempel & Rodgers, 1997).
La plupart des scientifiques ont souvent ignoré ces biais lors des analyses statistiques sur l’utilisation et la sélection de l’habitat. Cependant, il existe différentes méthodes pour contrer ces erreurs. En ce qui concerne le manque de précision, il faut réechantilloner ou accroître l’échantillon (Kenow et al., 2001). On peut aussi mesurer l’imprécision et la prendre en compte dans l’exploitation des données. Pour les données manquantes, il faut tester si leur nombre est important et si elles se répartissent équitablement entre les différents milieux traversés par l’animal.
PARTIE II : Matériel et Méthode
1. Sites d’étude
Les données analysées dans cette étude ont été récoltées sur des individus provenant de la Vallée de l’Ouche et de la Vallée de l’Auxois en Côte d’Or (Figure 1). La Vallée de l’Ouche s’étend de Pont-de-Pany à Bligny-sur-Ouche. Certaines captures ont été effectuées près de Jaugey (47,252559Lat ; 4,728929Long) et au Château de l’Oiserolle (47,256997Lat ; 4,688050Long). Les autres captures ont été effectués en Vallée de l’Auxois à Grenant-les-Sombernon (47,271351Lat ; 4,710148Long), à Braux (47,408115Lat ; 4,424252Long) et à La Roche Vanneau (47,474060Lat ; 4,524380Long). Les blaireaux ont été capturés à différents endroits avec différents degrés d’éloignement aux pâtures (entre 10m et 500m) dans le but de voir si la distance entre le terrier et la pâture la plus proche influence l’utilisation de la pâture par le blaireau. Le climat est tempéré et les deux zones comportent des prairies, des bocages, des forêts de feuillus, des zones agricoles. La végétation est constituée de chênaie-charmaie (Carpinus betulus et Quercus pubescens) en plateaux et d’hêtraies (Fagus sylvatica) en pentes.
10 Figure 1 : Carte représentative de la Côte d’Or et les différentes vallées où ont été effectuées les captures (*représentent les endroits précis des captures).
2. Capture et manipulation des blaireaux
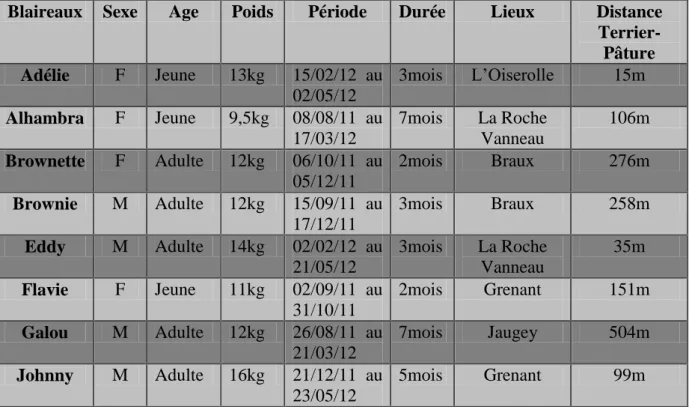
Pour la capture, les individus ont été appâtés avec du maïs dans des cages proche de leur terriers. Les individus ont été anesthésiés dans la cage à la sarbacane en utilisant un mélange de kétamine et de médéthomidine. Lorsque les animaux sont stables, le vétérinaire présent sur les lieux réalise un suivi anesthésique et différents prélèvements (écouvillons laryngés, fécal et prise de sang) dans le but de connaître le statut du blaireau vis-à-vis de M. bovis et de déterminer les voies d’excrétion de la bactérie. Chaque animal est identifié par la présence de boucles numérotées sur les oreilles (rouge pour les mâles et jaune pour les femelles). Cela permet de savoir si l’animal a déjà été capturé et d’identifier le groupe. De plus, cela permet de montrer aux chasseurs et aux agriculteurs les animaux qui sont capturés pour éviter de possibles pertes. Plusieurs données ont été relevées lors des captures : le sexe, l’âge, le poids, la dentition et un nom leur a été attribué (Tableau 2). La pesée s’effectue avec une balance
11 manuelle. Seuls les individus de plus de 10 kg sont équipés de colliers GPS. Sur les autres sont juste posées les boucles auriculaires et tous sont prélevés.
3. Acquisition des données de localisation
L’objectif était d’équiper 10 blaireaux si possible de groupes différents et avec un équilibre entre mâles et femelles. Les captures ont débuté le 1er août 2011 et à ce jour, 38 blaireaux ont été capturés et 9 colliers ont pu être posés. Les colliers sont des colliers GPS de marque Lotek, (modèle Small wildcell) pesant 400g. Ils émettent également en VHF (afin de retrouver le collier une fois que le GPS ne fonctionne plus). Ils transmettent les localisations au fur et à mesure par SMS grâce à une carte SIM intégrée au collier et à une base de réception qui se connecte à l’ordinateur. Les paramètres mesurés à chaque point sont la date et heure, la position GPS en mode différentiel (latitude, longitude, hauteur,) le nombre de satellites, une mesure de la qualité du point (DOP= « Dilution Of Precision ») et la température (Annexe 1). Les colliers ont été programmés pour prendre une localisation toutes les 1/2h pendant la nuit (de 19h ou 20h à 8h ou 9h selon les saisons). La durée de vie théorique de batterie indiquée par le fournisseur était de 5 à 7 mois (selon le temps moyen d’acquisition du point GPS) mais en pratique seuls 2 colliers ont eu cette durée de fonctionnement, les autres ayant duré moins longtemps (Tableau 2).
Blaireaux Sexe Age Poids Période Durée Lieux Distance Terrier-Pâture Adélie F Jeune 13kg 15/02/12 au 02/05/12 3mois L’Oiserolle 15m Alhambra F Jeune 9,5kg 08/08/11 au 17/03/12 7mois La Roche Vanneau 106m Brownette F Adulte 12kg 06/10/11 au 05/12/11 2mois Braux 276m Brownie M Adulte 12kg 15/09/11 au 17/12/11 3mois Braux 258m Eddy M Adulte 14kg 02/02/12 au 21/05/12 3mois La Roche Vanneau 35m Flavie F Jeune 11kg 02/09/11 au 31/10/11 2mois Grenant 151m Galou M Adulte 12kg 26/08/11 au 21/03/12 7mois Jaugey 504m Johnny M Adulte 16kg 21/12/11 au 23/05/12 5mois Grenant 99m
12
4. Télémétrie
Pour chaque individu, un certain nombre de localisations ne sont pas prises en compte dans l’analyse soit parce qu’elles sont trop imprécises (données non exploitables dues à une « DOP » élevé, à un nombre de satellites insuffisant) soit parce le point n’a pas pu être pris du tout (données manquantes dues à la position de l’animal, au couvert végétal et nuageux…). La proportion de localisations non validées et manquantes a été calculée pour chaque individu. Afin d’identifier un éventuel biais pour l’analyse, la répartition des points non exploitables en milieu ouvert et fermé a été testée. Ce test n’a pu être fait pour les données manquantes par manque de temps.
5. Estimation des domaines vitaux
Les domaines vitaux des blaireaux ont été estimés à partir des localisations GPS par la méthode de Kernel fixe (Worton, 1989). Celle-ci fournit une valeur des Distributions d’Utilisation (UD) permettant ensuite d’effectuer une estimation des domaines vitaux. En 1996, Seaman et Powell, ont montré que cette méthode en utilisant la méthode du LSCV pour déterminer le paramètre de lissage h, est celle qui comportait le moins de biais et permettait d’obtenir jusqu’à 95% d’UD. Le choix s’est donc porté sur cette méthode. La détermination de h par la méthode du LSCV n’a pas été possible pour deux blaireaux (Brownie et Brownette). En effet, l’algorithme mathématique utilisé par le logiciel n’a pas pu converger et aucun résultat n’est ressorti. Pour calculer leurs domaines vitaux, la méthode du kernel fixe avec le paramètre de lissage de référence ad hoc a été utilisée. Les domaines vitaux ont aussi été calculés à partir de la méthode du MCP afin de pouvoir comparer nos résultats avec d’autres études.
Deux échelles ont été développées dans cette analyse: tout d’abord une estimation des domaines vitaux à 95% d’UD qui a permis de travailler sur l’ensemble du domaine vital, puis une estimation des centres des domaines vitaux ou « core area » à 50% et 30% d’UD. L’ensemble du domaine vital concerne la plupart des déplacements des individus et les centres représentent des zones fortement exploitées.
Afin de s’assurer que les points pris toutes les 1/2h n’étaient pas autocorrélés et ainsi ne n’avait pas d’impact sur la taille et la forme des domaines vitaux, ils ont été comparés aux domaines vitaux calculés avec une période de deux heures entre chaque localisation (dans ce cas, chaque localisation était indépendante).
13 Les domaines vitaux ont été calculés sur la période totale de capture des individus. Certains domaines ont pu être calculés et affinés en fonction des saisons lorsque la période le permettait (pour Alhambra et Galou) :
• « Hiver » : 10 décembre au 23 février : baisse d’activité du aux températures, gestation et mise bas,
• « Printemps » : 24 février au 23 mai : reprise de l’activité après l’hiver, élevage des jeunes au terrier,
• « Eté » : 08 aout au 13 octobre : sevrage des jeunes,
• « Automne » : 14 octobre au 09 décembre : accumulation de réserves de graisse.
6. Estimation de l’utilisation de l’habitat et taux de contact
L’utilisation de l’habitat a été estimée grâce aux localisations GPS. Elle consiste à connaître le nombre de localisations par milieu (forêt, pâture, culture, bâtiment et lisière) sur le nombre de localisation total. Le taux d’utilisation obtenue pour chaque milieu correspondra aux pourcentages d’utilisation des différents types d’habitat du blaireau au sein de son domaine vital. L’utilisation de l’habitat a été affinée par saison pour Alhambra et Galou.
Le taux de contact entre les blaireaux et les bovins a été estimé en calculant pour chaque blaireau, le rapport du nombre de localisations en pâture et en bâtiment d’élevage sur le nombre total de localisations. Cela permet d’obtenir l’utilisation par les blaireaux de l’habitat où se trouvent les bovins. Un deuxième calcul a été effectué à l’intérieur des pâtures afin d’affiner les résultats. Les types d’habitat pris en compte sont alors les bosquets, la lisière et la pâture exclusive. Pour chaque blaireau, une couche a été crée à partir de leur situation géographique et a été capturée par image via GoogleEarth puis géoréférencé sur MapInfo. A partir des différentes couches « Jaugey », « Oiserolle », « Braux », « Roche Vanneau », « Grenant », des polygones pour chaque type de milieux ont été construits.
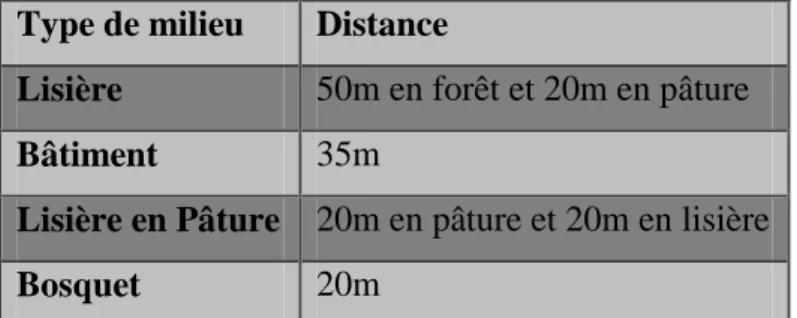
Pour contrer le manque de précision sur les données de localisation, l’imprécision a été mesurée et les données ont été traitées en fonction de cette imprécision. Ainsi, la limite de chaque milieu a été entourée d’une zone tampon dont la largeur correspond à l’imprécision mesurée pour chaque milieu par des tests préalables sur les colliers (Annexe 2). L’imprécision a été estimée à 20m en milieu fermé et à 10 m en milieu ouvert. Ainsi, par exemple, pour la lisière, définie d’après la littérature par 10m en milieu ouvert et 30m en milieu fermé (Do Linh San, 2002), une zone tampon de 20 m en milieu fermé et de 10 m en milieu ouvert a été
14 ajoutée pour la constitution des polygones correspondant à cet habitat. Le tableau 3 récapitule les largeurs des zones tampons pour chaque type de milieu.
Type de milieu Distance
Lisière 50m en forêt et 20m en pâture
Bâtiment 35m
Lisière en Pâture 20m en pâture et 20m en lisière Bosquet 20m
Tableau 3 : Récapitulatif des distances choisies pour les différents types de milieu
7. Analyses statistiques
Afin de tester la répartition des points validés entre le milieu ouvert et le milieu fermé, un test du Chi-deux a été réalisé. Lorsque les données étaient inférieures ou égales à 5, un test exact de Fisher a été utilisé. Deux tests du Chi-deux ont été utilisé pour les deux individus Alhambra et Galou afin de comparer la taille des domaines vitaux par saison. Pour vérifier s’il existait une relation entre l’utilisation des pâtures et la distance entre le terrier et la pâture, un test de corrélation de Spearman a été réalisé. Pour chaque blaireau, un test du Chi-deux a été utilisé afin d’investiguer une différence dans l’utilisation de l’habitat pour l’ensemble des localisations. Un test de Kruskal-Wallis a été utilisé pour comparer l’utilisation de l’habitat en fonction des saisons pour le blaireau Alhambra avec pour facteur la saison. Un test du Chi-deux a été utilisé pour Alhambra afin de visualiser une préférence d’habitat au sein de chaque saison. Avant chaque test, la normalité des données et/ou l’homoscédasticité des variances ont été vérifiées. Toutes ces analyses statistiques ont été réalisées sous le logiciel R avec un seuil α de 5% et le package adehabitat pour le calcul des domaines vitaux. Le nombre de réplicats par individu étant insuffisant, l’apport de statistiques descriptives a semblé être une bonne alternative. Les différents traitements SIG ont été réalisés sous le logiciel MapInfo.
PARTIE III : Résultats et Critiques
1. Résultats
a. Suivi télémétrique
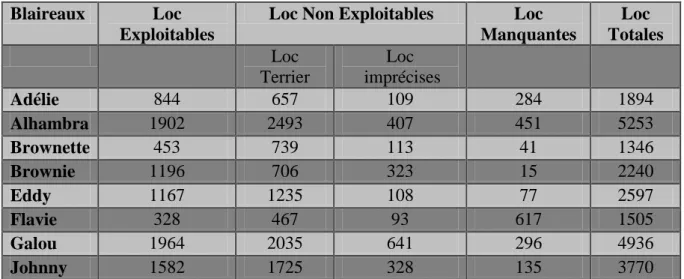
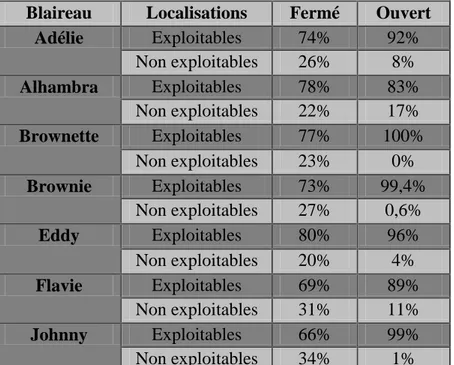
Le nombre de localisations exploitables et non exploitables pour chaque blaireau a été calculé (Tableau 4). Le nombre de localisations non exploitables à l’analyse ont été calculées
15 et le pourcentage de localisations donc la perte d’information par rapport aux localisations totales ont été mesurés (Tableau 5). La répartition des localisations exploitables et non exploitables en milieu ouvert et fermé a été mesurée pour chaque individu (Tableau 6). La répartition des données entre milieux est statistiquement différente pour l’ensemble des blaireaux (Pour Adélie : =49,684 ; p= 1,806e-12, pour Alhambra : =5,872 ; p=0,015, pour Brownette : F=22,594; p=4,476e-09, pour Brownie : F=102,552 ; p=4,476e-09, pour
Eddy : = 79,574 ; p< 2,200e-16 , pour Flavie : =23,169 ; p=1,484e-06, pour Johnny : =358,012 ; p=< 2,200e-16).
Blaireaux Loc Exploitables
Loc Non Exploitables Loc Manquantes Loc Totales Loc Terrier Loc imprécises Adélie 844 657 109 284 1894 Alhambra 1902 2493 407 451 5253 Brownette 453 739 113 41 1346 Brownie 1196 706 323 15 2240 Eddy 1167 1235 108 77 2597 Flavie 328 467 93 617 1505 Galou 1964 2035 641 296 4936 Johnny 1582 1725 328 135 3770
Tableau 4 : Représentation des localisations pour chaque individu
Blaireaux Pourcentage Loc non exploitables (terrier) Pourcentage Loc non exploitables (imprécises) Pourcentage Loc manquantes Pourcentage total Adélie 35% 6% 15% 56% Alhambra 47% 8% 9% 64% Brownette 56% 8% 3% 67% Brownie 32% 14% 0.7% 47% Eddy 48% 4% 3% 55% Flavie 31% 6% 40% 77% Galou 41% 13% 6% 60% Johnny 46% 9% 4% 59%
16
Blaireau Localisations Fermé Ouvert Adélie Exploitables 74% 92% Non exploitables 26% 8% Alhambra Exploitables 78% 83% Non exploitables 22% 17% Brownette Exploitables 77% 100% Non exploitables 23% 0% Brownie Exploitables 73% 99,4% Non exploitables 27% 0,6% Eddy Exploitables 80% 96% Non exploitables 20% 4% Flavie Exploitables 69% 89% Non exploitables 31% 11% Johnny Exploitables 66% 99% Non exploitables 34% 1%
Tableau 6 : Représentation des localisations exploitables et non exploitables imprécises (et non celles du à la présence du blaireau dans son terrier) en pourcentage en fonction du milieu (ouvert et fermé)
Pour vérifier l’autocorrélation des données, le domaine vital mesuré avec une localisation toutes les demi-heures a été comparé à celui mesuré avec une localisation toutes les deux heures (Image 1 à Image 8).
Image 1 : Représentation du domaine vital à 95% d’UD avec une localisation toutes les demi-heures (en rouge) et une localisation toutes les deux demi-heures (en bleu) chez Adélie à L’Oiserolle
17 Image 2 : Représentation du domaine vital à 95% d’UD avec une localisation toutes les demi-heures (en rouge) et une localisation toutes les deux demi-heures (en bleu) chez Alhambra à La Roche Vanneau
Image 3 : Représentation du domaine vital à 95% d’UD avec une localisation toutes les demi-heures (en rouge) et une localisation toutes les deux demi-heures (en bleu) chez Eddy à La Roche Vanneau
18 Image 4 : Représentation du domaine vital à 95% d’UD avec une localisation toutes les demi-heures (en rouge) et une localisation toutes les deux demi-heures (en bleu) chez Brownette à Braux
Image 5 : Représentation du domaine vital à 95% d’UD avec une localisation toutes les demi-heures (en rouge) et une localisation toutes les deux demi-heures (en bleu) chez Brownie à Braux
19 Image 6 : Représentation du domaine vital à 95% d’UD avec une localisation toutes les demi-heures (en rouge) et une localisation toutes les deux demi-heures (en bleu) chez Johnny à Grenant
Image 7 : Représentation du domaine vital à 95% d’UD avec une localisation toutes les demi-heures (en rouge) et une localisation toutes les deux demi-heures (en bleu) chez Flavie à Grenant
20 Image 8 : Représentation du domaine vital à 95% d’UD avec une localisation toutes les demi-heures (en rouge) et une localisation toutes les deux demi-heures (en bleu) chez Galou à Jaugey
b. Domaines Vitaux
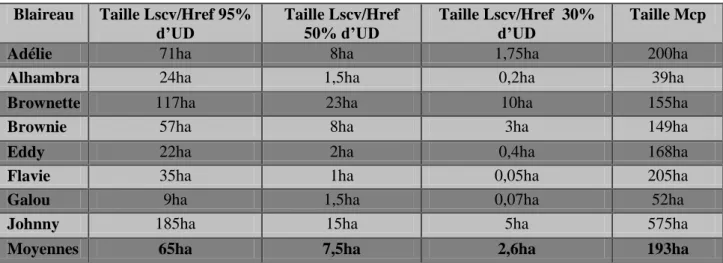
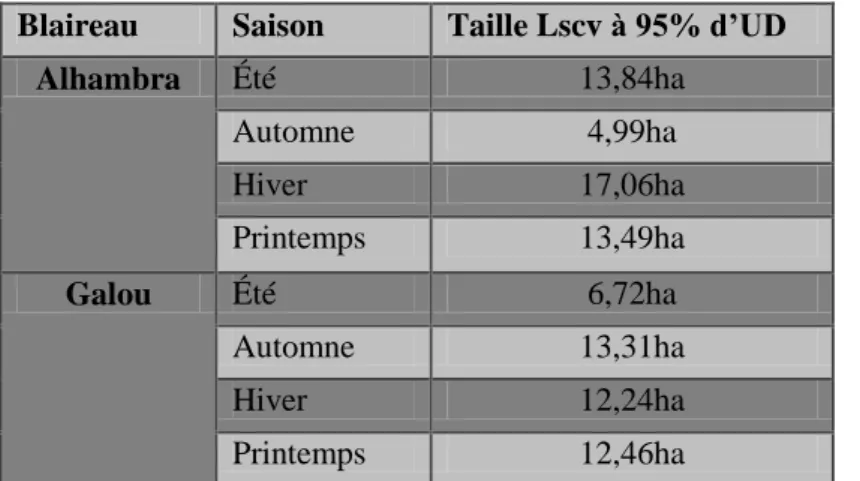
Les domaines vitaux ont été mesurés pour chaque blaireau avec la méthode du kernel fixe et du paramètre de lissage LSCV pour 95% d’UD (ou le h de référence dans le cas de Brownie et Brownette) et de la méthode du MCP (Image 11a à Image 16a). Ils ont aussi été mesurés pour 50% et 30% d’UD avec la méthode du kernel fixe (Image 11b à Image 16b). La moyenne des tailles des domaines vitaux sur l’ensemble des individus a été calculée pour chaque méthode utilisée (Tableau 7). Les domaines vitaux d’Alhambra et de Galou ont aussi été mesurés en fonction des saisons (Tableau 8). Pour ces deux blaireaux, il n’y a pas de différence concernant la taille du domaine vital en fonction de la saison (Pour Alhambra : =0,903 ; p=0,342 et pour Galou : =0,594 ; p=0,441) (Tableau 9).
21 Image 9: Représentation des domaines vitaux calculés avec la méthode du kernel fixe LSCV (en rouge) à 95% d’UD et du MCP (en bleu turquoise) (a) et ceux calculés avec la méthode du kernel fixe LSCV à 50% et 30% d’UD (b) de la blairelle Adélie avec ses localisations (*) et son terrier ( )
Image 10: Représentation des domaines vitaux calculés avec la méthode du kernel fixe LSCV (en rouge) à 95% d’UD et du MCP (en bleu turquoise) (a) et ceux calculés avec la méthode du kernel fixe LSCV à 50% et 30% d’UD (b) de la blairelle Alhambra avec ses localisations (*) et son terrier ( )
22 Image 11 : Représentation des domaines vitaux calculés avec la méthode du kernel fixe LSCV (en rouge) à 95% d’UD et du MCP (en bleu turquoise) (a) et ceux calculés avec la méthode du kernel fixe LSCV à 50% et 30% d’UD (b) de la blairelle Brownette avec ses localisations (*) et son terrier ( )
Image 12: Représentation des domaines vitaux calculés avec la méthode du kernel fixe LSCV (en rouge) à 95% d’UD et du MCP (en bleu turquoise) (a) et ceux calculés avec la méthode du kernel fixe LSCV à 50% et 30% d’UD (b) du blaireau Brownie avec ses localisations (*) et son terrier ( )
23 Image 13 : Représentation des domaines vitaux calculés avec la méthode du kernel fixe LSCV (en rouge) à 95% d’UD et du MCP (en bleu turquoise) (a) et ceux calculés avec la méthode du kernel fixe LSCV à 50% et 30% d’UD (b) du blaireau Eddy avec ses localisations (*) et son terrier ( )
Image 14 : Représentation des domaines vitaux calculés avec la méthode du kernel fixe LSCV (en rouge) à 95% d’UD et du MCP (en bleu turquoise) (a) et ceux calculés avec la méthode du kernel fixe LSCV à 50% et 30% d’UD (b) de la blairelle Flavie avec ses localisations (*) et son terrier ( )
24 Image 15 : Représentation des domaines vitaux calculés avec la méthode du kernel fixe LSCV (en rouge) à 95% d’UD et du MCP (en bleu turquoise) (a) et ceux calculés avec la méthode du kernel fixe LSCV à 50% et 30% d’UD (b) du blaireau Johnny avec ses localisations (*) et son terrier ( )
Image 16 : Représentation des domaines vitaux calculés avec la méthode du kernel fixe LSCV (en rouge) à 95% d’UD et du MCP (en bleu turquoise) (a) et ceux calculés avec la méthode du kernel fixe LSCV à 50% et 30% d’UD (b) du blaireau Galou avec ses localisations (*) et son terrier ( )
25
Blaireau Taille Lscv/Href 95%
d’UD Taille Lscv/Href 50% d’UD
Taille Lscv/Href 30%
d’UD Taille Mcp
Adélie 71ha 8ha 1,75ha 200ha
Alhambra 24ha 1,5ha 0,2ha 39ha
Brownette 117ha 23ha 10ha 155ha
Brownie 57ha 8ha 3ha 149ha
Eddy 22ha 2ha 0,4ha 168ha
Flavie 35ha 1ha 0,05ha 205ha
Galou 9ha 1,5ha 0,07ha 52ha
Johnny 185ha 15ha 5ha 575ha
Moyennes 65ha 7,5ha 2,6ha 193ha
Tableau 7: Représentation de la taille des domaines vitaux de chaque blaireau calculés avec des méthodes différentes et les moyennes correspondantes
Blaireau % de la superficie du domaine vital
Adélie 11% Alhambra 6% Brownette 20% Brownie 13% Eddy 8% Flavie 3% Galou 15% Johnny 8% Moyenne 10.5%
Tableau 8 : Représentation du pourcentage des aires d’intense activité à 50% d’UD sur la superficie totale du domaine vital
26 Image 17 : Représentation des domaines vitaux d’Alhambra calculés avec la méthode du kernel fixe LSCV pour chaque saison (en blanc pour l’hiver, en rose pour le printemps, en jaune pour l’été et en bleu pour l’automne)
Image 18 : Représentation des domaines vitaux de Galou calculés avec la méthode du kernel fixe LSCV pour chaque saison (en blanc pour l’hiver, en rose pour le printemps, en jaune pour l’été et en bleu pour l’automne)
27
Blaireau Saison Taille Lscv à 95% d’UD
Alhambra Été 13,84ha Automne 4,99ha Hiver 17,06ha Printemps 13,49ha Galou Été 6,72ha Automne 13,31ha Hiver 12,24ha Printemps 12,46ha
Tableau 9 : Représentation de la taille des domaines vitaux correspondant aux saisons des deux blaireaux Alhambra et Galou
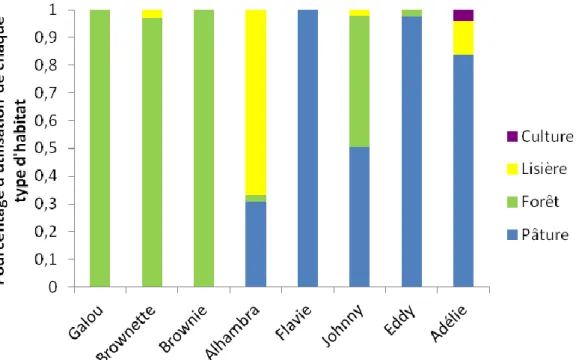
c. Utilisation de l’habitat
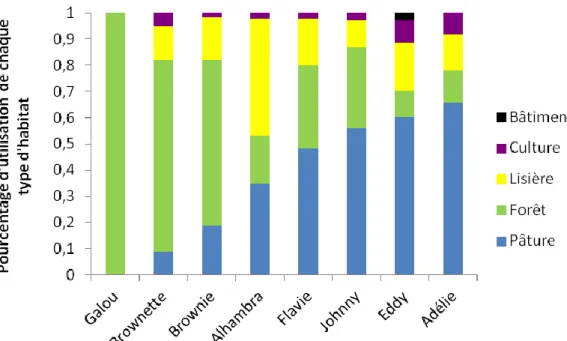
Le pourcentage d’utilisation de chaque habitat pour chaque blaireau a été mesuré pour l’ensemble des localisations (Figure 2), pour les localisations présentent au sein du domaine vital à 50% d’UD (Figure 3) et pour les localisations présentes au sein du domaine vital à 30% d’UD (Figure 4). Sur l’ensemble des localisations, chaque blaireau utilise ces habitats de manière statistiquement différente (Pour Adélie : =166,541 ; p<2.200e-6, pour Alhambra :
=743,696 ; p<2.200e-6, pour Brownette : =13,527 ; p=2,351e-4, pour Brownie :
=19,946; p=7,964e-6, pour Eddy : = 297,415 ; p< 2,200e-6 , pour Flavie : =50,120 ;
p=1,111e-12, pour Johnny : =143,322 ; p=< 2,200e-6). Les calculs n’ont pas été effectués pour le blaireau Galou car il utilise exclusivement la forêt.
Il n’existe pas de différence statistique dans l’utilisation de l’habitat par le blaireau Alhambra en fonction du facteur saison (Kruskal-Wallis : K=3,154 ; p=0,368). A chaque saison, elle utilise les habitats de manière statistiquement différente (Pour l’été : =284,912 ; p< 2,200e-16, pour l’automne : =136,464 ; p< 2,200e-16, pour l’hiver : =35,722 ; p=2,275e-9, pour le printemps : =260,585 ; p< 2,200e-16) (Figure 5).
28 Figure 2: Représentation de l’utilisation de chaque type d’habitat pour chaque blaireau au sein de l’ensemble de leur domaine vital
Figure 3: Représentation de l’utilisation de chaque type d’habitat au sein du domaine vital à 50% d’UD pour chaque blaireau
29 Figure 4: Représentation de l’utilisation de chaque type d’habitat au sein du domaine vital à 30% d’UD pour chaque blaireau
Figure 5: Représentation de l’utilisation de chaque type d’habitat en fonction de chaque saison chez le blaireau Alhambra
30 d. Taux de contact
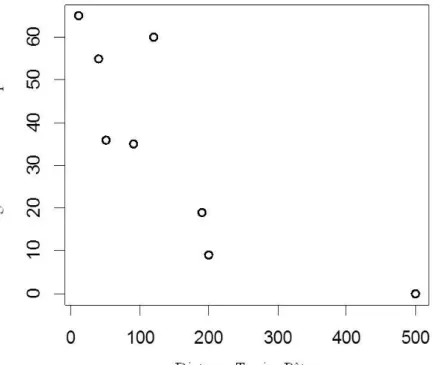
Il existe une corrélation entre le facteur distance terrier aux pâtures et l’utilisation de ces dernières (Coefficient de corrélation=0,857 ; p=0,011) : plus les terriers sont proches, plus l’utilisation de ces dernières est importante (Figure 6).
Figure 6 : Représentation de la corrélation entre l’utilisation des pâtures et les distances terrier-pâture
Le taux de contact entre les blaireaux et les bovins ont été calculés pour tous les individu à partir de l’ensemble de leur localisations (Tableau 10). L’utilisation de la pâture exclusive par rapport à l’utilisation des bosquets et de la lisière en pâture a été mesurée (Figure 7).
Blaireaux Pâture Bâtiment Total
Galou 0% 0% 0% Brownette 9% 0% 9% Brownie 19% 0% 19% Alhambra 35% 0% 35% Flavie 48% 0% 48% Johnny 56% 0% 56% Eddy 60% 3% 63% Adélie 66% 0% 66%
Tableau 10: Représentation du pourcentage d’utilisation de la pâture et des bâtiments accueillant des cheptels bovins pour chaque blaireau
31 Figure 7: Représentation du pourcentage d’utilisation de la lisière en pâture et des bosquets en pâture contre le pourcentage d’utilisation de la pâture exclusivement
2. Discussion et critiques
Dans cette étude, les colliers des blaireaux ont permis d’obtenir un minimum de 100 localisations, ce qui a permis d’utiliser la méthode du kernel fixe avec le paramètre de lissage « LSCV » lors de la mesure des domaines vitaux (Seaman et al., 1999). Néanmoins, à l’analyse des colliers GPS de chaque individu, on trouve un certain nombre de localisations non exploitables. Elles représentent en moyenne 60% des localisations acquises par le collier GPS. Les localisations non exploitables qui correspondent à l’emplacement du blaireau dans son terrier sont élevées. Elles représentent en moyenne 42% de perte de localisations. Ce qui peut paraître normal puisque le blaireau peut passer jusqu’à 70% de son temps nocturne dans son terrier (Roper, 2010). Finalement, ces localisations ne sont donc pas une grosse perte pour l’étude puisqu’elles ont un sens biologique. Les localisations non exploitables qui sont du à un manque de précision représentent en moyenne 8.5% sur l’ensemble des localisations. Ce manque de précision peut être du à la couverture végétale. Si l’on regarde les données fournies par les colliers de Brownie et de Galou, ils ont respectivement 14% et 13% de localisations imprécises. Or, d’après les résultats d’utilisation d’habitat, ces deux blaireaux
32 passent le plus clair de leur temps en fôret. Les résultats montrent que le nombre de localisations non exploitables en milieu ouvert est significativement plus faible par rapport à celles présentes en milieu fermé. Ce résultat est cohérent avec l’étude de Rumple & Rodgers, 1997, qui ont mis en évidence qu’un collier GPS est 3.8 fois moins susceptible d’acquérir une localisation en forêt (>15m de hauteur) qu’en zone dépourvue d’arbres. De plus, Brendel et
al., 2012, ont montré que le succès de localisation diminuait lorsque le blaireau sortait de son
terrier. Il existe un retard dans l’obtention des données de localisations à la sortie du terrier correspondant au temps recquis par le GPS pour mettre à jour les éphémérides (Brendel et al., 2012). Le comportement du blaireau peut aussi jouer un rôle dans l’acquisition des données. Les griffures et les toilettes des individus peuvent entraîner la rotation du collier, déplaçant ainsi l’antenne GPS de l’horizontale à la verticale, ce qui diminue les performances du collier (D'eon & Delparte, 2005).
Concernant le nombre de données manquantes, sur huit blaireaux, six d’entre eux ont moins de dix pourcent de localisations manquantes. Les blaireaux Adélie et Flavie ont 15% et 40% de données manquantes respectivement. Or, il s’avère que ces deux blaireaux possèdaient le même collier à des périodes différentes. Flavie portait le collier du 02 septembre au 31 octobre et Adélie le portait du 15 février au 02 mai. Le manque d’information est sûrement du à un problème dans la réception du collier. Ce résultat coïncide avec l’étude de Frair et al., 2004 qui ont montré qu’un collier GPS défectueux peut s’avérer être la cause du manque de données au sein d’une étude télémétrique. Pour les autres blaireaux, 10% de localisations manquantes est un chiffre faible même si il aurait fallu prendre en compte ce manque d’information en mesurant l’effet sur la taille. Par manque de temps, cette étude n’a pas pu le tester. Cela reste un problème très courant dans les études télémétriques (Jonhson et al., 1998). Dans cette étude, le fait d’avoir plus de localisations imprécises, et donc non prises en compte en forêt plutôt qu’en milieu ouvert, a du engendrer une sous-estimation des localisations en milieu fermé pour le calcul du domaine vital ou de l’utilisation de l’habitat.
En ce qui concerne l’autocorrélation des données, un intervalle de temps trop faible entre les localisations cause une sous-estimation du domaine vital. La moitié des individus présente un domaine vital inférieur à celui calculée avec un intervalle de deux heures entre chaque localisation. De plus, pour certains individus, les patchs créés par la méthode du kernel fixe ne sont pas au même endroit et sont parfois plus nombreux. Chez certains individus, les données sont donc probablement autocorrélées. En effet, un animal qui se
33 déplace à une vitesse finie relativement lente par rapport à l’acquisition du GPS peut décaler l’acquisition des données et toute localisation tend alors naturellement à être proche de la précédente (Bodin, 2005). Certains auteurs n’ont pas réussi à éliminer l'autocorrélation entre localisations quel que soit l’intervalle de temps choisi (Rooney et al., 1998, De Solla et al., 1999). En effet, l’indépendance des données est influencée par le mode d’utilisation de l’espace de l’espèce considérée.
Cette étude a tenu à mesurer les domaines vitaux des blaireaux en utilisant deux méthodes différentes : la méthode du kernel fixe et la méthode du MCP. Pour l’ensemble des blaireaux, la méthode du kernel fixe est bien plus précise que celle du MCP. En effet, le domaine vital calculé avec le kernel fixe a la forme de patchs entourant minutieusement les localisations des animaux prenant en compte l’intensité de l’utilisation de l’espace. Le MCP, donne un domaine vital beaucoup plus large en prenant juste les extrémitès des localisations. Dans la plupart des cas, de nombreuses zones sans localisations des individus sont prises en compte avec cette méthode. Elle sur-estime donc le domaine réel de l’individu. La méthode du kernel reflète donc mieux la structure interne du domaine vital de l’animal car elle prend en compte la distribution des localisations. Lors de l’utilisation de la méthode du kernel fixe, deux paramètres de lissage ont été utilisés même si le choix s’était porté sur le « LSCV ». Pour les deux blaireaux Brownette et Brownie, le calcul du domaine vital a été mesuré avec le paramètre de lissage h de référence. La méthode du kernel fixe avec pour paramètre de lissage le « LSCV » donne les estimations les plus précises de surfaces de domaines vitaux et permet une meilleure estimation des formes de la distribution d’utilisation (Seaman & Powell, 1996 ; Seaman et al., 1999). Alors que le paramètre de lissage de référence ad hoc surestime la surface de la distribution et semble sensible à l’auto-corrélation dans les données, il permet tout de même des estimations de valeurs raisonnables (Seaman & Powell, 1996 ; Seaman et
al., 1999).
La superficie des domaines vitaux individuels des 8 blaireaux suivis par radio-télémétrie varie de 9 hectares à 185 hectares et elle est en moyenne de 65 hectares avec la méthode du kernel fixe. Les domaines vitaux de nos blaireaux sont plus petits en moyenne comparé à ceux des Ardennes qui allaient de 100 à 450 hectares (Bodin, 2005). En Suisse, une étude mesure des domaines vitaux allant de 50 à 150 hectares (Do Linh San, 2002). Cette fourchette est cohérente avec nos résultats même si chez certains de nos individus les domaines vitaux sont très petits (Galou par exemple avec 9ha). La taille des domaines vitaux
34 des blaireaux varie considérablement d’une région à l’autre (Do Linh San, 2002). En effet, plusieurs études ont montré que les domaines vitaux étaient beaucoup plus grand et plus vaste lorsque l’abondance et la distribution de la ressource alimentaire du milieu était pauvre (320ha en moyenne dans les montagnes du Jura (Ferrari, 1997), 414ha en moyenne dans les forêts boréales (Broseth, 1997) et 406 à 780 ha en moyenne dans les milieux arides méditerranéens (Revilla & Palomares, 2002)). Au contraire, lorsque le milieu est riche, la taille des domaines vitaux est faible, 20 à 50 ha en moyenne pour l’Angleterre (Cheeseman et
al., 1987). La recherche de nourriture peut alors avoir un lien direct avec la distribution
spatiale et la densité des populations de blaireau.
La superficie des aires d’intense activité (déterminées à partir des 50% d’UD) varie de 1 à 15 hectares selon les individus, avec une moyenne de 7,5 hectares. Ces zones représentent en moyenne 10,5% de la superficie des domaines vitaux individuels. La moitié de l’activité des blaireaux est donc située dans à peine plus de 10% de l’ensemble des lieux qu’ils utilisent. Selon les individus, le domaine vital contient une à onze aires d’intense activité et, même lorsqu’il n’en existe qu’une, celle-ci n’est pas nécessairement centrée autour du terrier principal comme pour le domaine vital dans son ensemble. Cela implique que les blaireaux concentrent leurs déplacements routiniers dans des sites particuliers et qu’ils n’utilisent pas tout l’espace contenu dans leur domaine vital de manière homogène.
Il n’existe pas de différence statistique dans la superficie des domaines vitaux d’Alhambra et de Galou selon les saisons. Cependant, le domaine vital d’Alhambra en hiver est le plus grand, quant à Galou, c’est celui en Automne. Ces résultats sont en contradiction avec la littérature car en hiver les blaireaux sont en repos et leur activité est plus faible. Ces résultats peuvent être à l’origine d’une période hivernale relativement douce cette année. Il se peut aussi que les blaireaux sortent moins souvent mais suivent un plus grand parcours pour trouver de la nourriture.
Sur l’ensemble de leurs localisations, certains individus privilégient l’utilisation des pâtures (Adélie, Eddy et Johnny), d’autres le milieu forestier (Galou de manière exclusive, Brownie et Brownette), et d’autres encore utilisent l’habitat de manière relativement homogène (Alhambra et Flavie). La culture est très peu utilisée par l’ensemble des blaireaux et les bâtiments utilisés par Eddy uniquement représentent une faible proportion par rapport aux différents habitats. Sur les localisations correspondant aux aires d’intenses activités, certains habitats sont devenus exclusifs, comme la pâture et la forêt. Les pâtures peuvent être