Effets des protéines S100A8 et S100A9 dans la
différenciation cellulaire dans la leucémie myéloïde
aiguë
Thèse
Malika Laouedj
Doctorat en biologie cellulaire et moléculaire
Philosophiae Doctor (Ph. D.)
Québec, Canada
Effets des protéines S100A8 et S100A9 sur la
différenciation cellulaire dans la leucémie myéloïde
aiguë
Thèse
Malika Laouedj
Sous la direction de :
Frédéric Barabé, directeur de recherche
Philippe Tessier, codirecteur de recherche
Résumé
Les leucémies myéloïdes aiguës (LMA) sont des hémopathies rares, mais très agressives. Elles résultent d’un dérèglement du processus d’hématopoïèse qui se caractérise par une prolifération incontrôlée de cellules sanguines immatures engagées dans la lignée myéloïde. En dépit des traitements actuels qui reposent sur l’utilisation d’agents chimiothérapeutiques ciblant les cellules en prolifération, le pronostic des patients souffrants de LMA est très sombre. En effet, seuls 30% des patients souffrants de LMA survivent au-delà de 5 ans suivant la prise en charge thérapeutique. L’identification des acteurs participant au développement et au maintien des LMA est donc cruciale pour l’élaboration d’une stratégie thérapeutique efficace et ciblée.
S100A8 et S100A9 sont des protéines fixatrices de calcium exprimées par les neutrophiles et les monocytes. Ce sont des alarmines jouant des rôles clés dans l’inflammation et dans des pathologies causées par une inflammation excessive. Les protéines S100A8 et S100A9 exercent également de multiples fonctions dans divers tumeurs solides. Elles favorisent la formation de niche pré-métastasique et inhibe la réponse immunitaire antitumorale. Une analyse du génome par séquençage a mis en évidence que S100A8 et S100A9 sont fortement exprimées chez les patients atteints de LMA. De plus, l’expression de la protéine S100A8 chez les patients souffrants de LMA serait corrélée avec un faible taux de survie. Principalement étudiées dans les tumeurs solides, les fonctions des protéines S100A8 et S100A9 dans les néoplasies hématologiques telles que les leucémies sont très peu documentées. Dans ces travaux de thèse, nous nous sommes donc intéressés aux rôles exercés par les protéines S100A8 et S100A9 dans les leucémies myéloïdes aiguës.
À l’aide d’un modèle murin de LMA induit par la surexpression des facteurs HOXA9 et
MEIS1 dans des cellules souches/progénitrices hématopoïétiques, nous avons démontré
l’existence d’une fraction de cellules exprimant les protéines S100A8 et S100A9. Celle-ci est également retrouvée chez les patients atteints de leucémies aiguës myélomonocytaires et monocytaires (M4-M5 d’après la classification FAB).
Les études menées in vivo et in vitro révèlent que la protéine S100A9 induit la différenciation des cellules leucémiques, tandis que la protéine S100A8, préviens l’effet de S100A9 permettant de maintenir ainsi le phénotype immature des cellules LMA. Le traitement par la protéine recombinante S100A9 permet d’accroitre la maturation des cellules LMA, diminue leur prolifération et prolonge la survie des souris LMA. De la même façon le traitement par les
anticorps anti-S100A8 provoque un effet similaire au traitement par la protéine S100A9. Nos résultats suggèrent que de forts ratios de S100A9 sur S100A8 sont requis pour induire la différenciation des cellules LMA.
Le mécanisme intracellulaire par lequel S100A9 induit la différenciation des cellules leucémiques a également été étudié dans le cadre de cette thèse. Nous avons identifié que S100A9 via la liaison au récepteur TLR (Toll-like receptor) active les voies de signalisations Mitogen Activated Protein Kinase p38, Jun N-terminal Kinase et extracellular signal-regulated kinases 1 et 2 et provoque la différenciation des cellules leucémiques. Les essais menés sur des cellules primaires de patients malades ont permis de confirmer la capacité de S100A9 et de S100A8 à réguler la différenciation des cellules leucémiques.
En somme, les données présentées dans cette thèse contribuent à une meilleure compréhension des rôles des protéines S100A8 et S100A9 dans la différenciation des cellules myéloïdes. Par ailleurs, nos données permettent également d’entrevoir les bénéfices thérapeutiques liés au blocage de S100A8 ou à l’augmentation de S100A9 dans les LMA.
Abstract
Acute myeloid leukemias (AMLs) are rare but still aggressive hematological diseases. They are the result of a perturbed hematopoietic process characterized by an uncontrolled proliferation of hematopoietic cells committed to the myeloid lineage. Despite current therapy based on chemotherapeutic agents, aimed at killing proliferating cells, prognosis of AML patients is dismal and only 30 % of patients survived beyond 5 years. Identification of actors involved in the initiation and sustaining LMA is crucial to the development of efficient and targeted therapy strategy.
S100A8 and S100A9 are calcium-binding proteins predominantly expressed by neutrophils and monocytes, and play key roles in both normal and pathological inflammation. Recently, both proteins were found to promote tumor progression through the establishment of pre-metastatic niches and to inhibit antitumor immune responses. Although S100A8 and S100A9 have been studied in solid cancers, their functions in hematological malignancies remain poorly understood. However, S100A8 and S100A9 are highly expressed in acute myeloid leukemia (AML), and S100A8 expression has been linked to a poor prognosis in AML. Although the roles of these proteins were studies in solid tumor, little is known in their functions in hematological malignancies. We studied in this thesis the role of S100A8 and S100A9 in acute myeloid leukemia.
Using AML mouse model of AML surexpressing HOXA9 and MEIS1 in hematopoietic stem and progenitor cells, we identified a small subpopulation of cells expressing S100A8 and S100A9. This subpopulation was consistently found in AML samples from patients with myelomonocytic and monocytic leukemias (M4 and M5 according FAB classification).
In vitro and in vivo analyses revealed that S100A9 induces AML cell differentiation,
whereas S100A8 prevents differentiation induced by S100A9 activity and maintains AML immature phenotype. Treatment with recombinant S100A9 proteins increased AML cell maturation, induced growth arrest, and prolonged survival in an AML mouse model. Interestingly, anti-S100A8 antibody treatment had effects similar to S100A9 therapy in vivo, suggesting that high ratios of S100A9 over S100A8 are required to induce differentiation. In this thesis, the mechanism of S100A9 leading to differentiation of leukemic cells was also study. Our in vitro
studies on the mechanisms/pathways involved in leukemic cell differentiation revealed that binding of S100A9 to toll-like receptor 4 (TLR4) promotes activation of p38 mitogen-activated protein kinase, extracellular signal-regulated kinases 1 and 2, and Jun N-terminal kinase signaling pathways, leading to myelomonocytic and monocytic AML cell differentiation. Overall, our findings indicate that S100A8 and S100A9 are regulators of myeloid differentiation in leukemia and have therapeutic potential in myelomonocytic and monocytic AMLs.
Table des matières
Résumé ... iii
Abstract ... v
Table des matières ... vii
Listes des tableaux ... x
Liste des figures ... xi
Liste des abréviations ... xii
Remerciements ... xv
Avant-propos ... xvi
Chapitre 1 : Introduction ... 1
1.1. Leucémies myéloïdes aiguës ... 1
1.1.1. Généralités sur les leucémies myéloïdes aiguës ... 1
1.1.1.1. Généralités et définitions ... 1
1.1.1.2. Épidémiologie ... 2
1.1.1.3. Étiologie ... 3
1.1.2. Leucémogenèse ... 4
1.1.2.1. Hématopoïèse physiologique... 4
1.1.2.2. Pathogenèse des LMA ... 24
1.1.3. Diagnostic et classification ... 34 1.1.3.1. Diagnostic ... 34 1.1.3.2. Classification... 35 1.1.4. Traitements et pronostic ... 37 1.1.4.1. Traitements ... 37 1.1.4.2. Facteurs pronostiques ... 40 1.2. Les protéines S100 ... 51
1.2.1. Généralités sur les protéines de la famille S100 ... 51
1.2.1.2. Fonctions des protéines S100 ... 53
1.2.2. Les protéines S100A8 et S100A9 ... 54
1.2.2.1. Généralités sur les protéines S100A8 et S100A9 ... 54
1.2.2.2. Profil d’expression des protéines S100A8 et S100A9 ... 55
1.2.2.3. Structure et conformation des protéines S100A8 et S100A9 ... 58
1.2.2.4. Fonctions intracellulaires des protéines S100A8 et S100A9 ... 60
1.2.2.5. Sécrétion des protéines S100A8 et S100A9 ... 62
1.2.2.6. Récepteurs liés par les protéines S100A8 et S100A9... 64
1.2.2.7. Fonctions extracellulaires des protéines S100A8 et S100A9 ... 66
1.2.2.8. Fonctions de la protéine S100A12 ... 70
1.2.2.9. Les protéines S100A8 et S100A9 comme cible thérapeutique ... 75
Chapitre 2 : Hypothèses et objectifs du projet de recherche ... 77
2.1. Mise en contexte ... 77
2.2. Hypothèse du projet de recherche ... 77
2.3. Objectifs et méthodologie ... 78
Chapitre 3 : La protéine S100A9 induit la différenciation des cellules leucémiques myéloïdes via TLR4 ... 80
3.1. Résumé ... 80
3.2. Abstract ... 82
3.3. Introduction ... 83
3.4. Material and methods ... 84
3.5. Results ... 87
3.11. Figures ... 95
3.12. Références... 104
3.13. Supplémental informations... 107
Chapitre 4 : Discussion et conclusions ... 117
4.1. Discussion et perspectives... 117
4.1.1. Les cellules exprimant les protéines S100A8et S100A9 (S100A+) dans les LMA 117 4.1.2. Rôle de la protéine S100A9 dans la différenciation des cellules myéloïde ... 119
4.1.2.2. Les cascades de signalisation intracellulaire activées par la protéine S100A9 menant à la différenciation des cellules LMA ... 128 4.1.3. Rôles de la protéine S100A8 dans les LMA... 129 4.1.4. L’hétérocomplexe S100A8/A9 (calprotectine) dans les LMA ... 133 4.1.5. Effets anti-tumoraux de S100A9 et de l’anti-S100A8 et possibles perspectives thérapeutiques ... 134 4.2. Conclusions ... 136 Bibliographie ... 139
Listes des tableaux
Tableau 1 : Phénotypes des différentes populations de cellules hématopoïétiques selon l’expression des marqueurs de surface. ... 11 Tableau 2: Expression des marqueurs de surface et cytoplasmique pour le diagnostic des leucémies myéloïdes aiguës. Adapté de Döhner et al., 2010 [5]. ... 35 Tableau 3 : Classification FAB des LMA [288] ... 36 Tableau 4 : Classification des leucémies myéloïdes aiguës et hémopathies apparentées par l’OMS révisée en 2016. Adapté de la revue de Arber et al., parue en 2016 [341]. ... 37 Tableau 5 : Stratification cytogénétique des LMA selon les recommandations de l’ELN. Adapté de la revue de Döhner et al., parue en 2010 [5]. ... 41 Tableau 6 : Principales caractéristiques des LMA des sujets jeunes et âgés. ... 43 Tableau 7 : Description de certaines des caractéristiques phénotypiques des souris déficientes en S100A9 [482, 483]. ... 61 Tableau 8 : Les protéines S100A8 et S100A9 dans les pathologies inflammatoires. Adapté de [520]. ... 72
Liste des figures
Figure 1 : Évolution de l’incidence des leucémies myéloïdes aiguës en fonction de la classe d’âge. ... 2 Figure 2 : Schéma simplifié de l’hématopoïèse. ... 6 Figure 3 : Les différents types de cellules et facteurs présents au sein de la niche hématopoïétique. ... 7 Figure 4: Modèle de division des cellules souches hématopoïétiques (Division asymétrique vs symétrique). ... 9 Figure 5 : Les étapes de maturation du neutrophile. ... 13 Figure 6 : Régulation transcriptionnelle de la myélopoïèse. ... 21 Figure 7 : Schéma simplifié représentant les modèles proposés pour expliquer l’origine des cellules souches leucémiques (CSL). ... 26 Figure 8 : Modèle de leucémogenèse à deux classes de mutations ... 27 Figure 9 : Changement de la survie globale des patients atteints de LMA avec l’âge. ... 42 Figure 10: Représentation schématique du locus chromosomique 1q21 de la famille S100. .. 52 Figure 11 : Structure secondaire des protéines S100. Schéma adapté de Donato et al. 2013 [439]... 53 Figure 12 : Structure primaire et alignement de séquence des protéines humaines S100A8 et S100A9. ... 55 Figure 13 : Schéma de la régulation de l’expression de S100A8 dans les macrophages murins. Schéma adapté de la revue de Donato et al., 2013 [444]. ... 57 Figure 14: Représentation en ruban de la structure tertiaire et quaternaire des protéines humaines S100A8 et S100A9. ... 59 Figure 15 : Activation des cellules endothéliales par les calgranulines et effet chimioattracteur sur les leucocytes. Schéma adapté de la revue de Kessel, Holzinger et Foell datant de 2013 [577]... 68 Figure 16 : Les protéines S100A8 et S100A9 dans l’inflammation. ... 70 Figure 17 : Rôles de la protéine S100A12 dans l’inflammation... 71
Liste des abréviations
1,25D 1, 25(OH)2 Vitamin D32-DG 2-deoxy-D-glucose
AA Acide Arachidonique
Aa Acide aminé
ADCC Antibody-Dependant Cell Cytotoxicity
AML Acute Myeloid Leukemia
ATRA Acide tout trans rétinoïque APL Acute Promyelocytic Leukemia
Akt/PKB Serine Threonine tyrosine kinase/Protein Kinase B”
ASXL1 Additional SeX Combs-Like Transcriptional Regulator 1
CAR CXCL12 Abondant Reticular cell
CBF Core Binding Factor
CCI Charlson Comorbidity Index
CD Cluster of Differentiation
CEBP CCAAT-enhancer-binding proteins
CMP Common Myeloid Progenitor
CN-LMA Caryotype normal-leucémie myéloïde aiguë
CLP Common Lymphoid Progenitor
CML Chronic Myeloid Leukemia
CP-10 Chemotactic protein-10 kDa
CREB c-AMP Response Element-Binding
CSH Cellules souche hématopoïétique CSF1R Colony Stimulating factor 1 Receptor
CSL Cellules souche leucémique CXCL12 C-X-C Motif Chemokine Ligand 12
CXCR4 C-X-C chemokine receptor type 4
DAMP Danger Associated Molecular Pattern
DNMT3 DNA MethylTransferase 3A
ECOG Eastern Cooperative Oncology Group
EGFP Enhanced Green Fluorescent Protein
EGR1 Early Growth Response gene-1
ELN European Leukemia Net
ERK Extracellular signal–Regulated kinases
FAB French-American-British
FACS Fluorescence-Activated Cell Sorting
FDA Food and Drug Administration
GMP Granulocyte/Macrophage Progenitor
GM-CSF Granulocyte Macrophage Colony Stimulating Factor
GO Gemtuzumab ozogamicin
Gro-α Growth-Regulated Oncogene-alpha
GVHD Graft Versus Host Disease
H9M1 HOXA9-MEIS1
HCT-CI Hematopoietic Cell Transplantation- Comorbidity Index
HDAC Histone déactylase
HOXA9 Homeobox A9
ICAM Intercellular Adhesion Molecule
ID Inhibitor of Differentiation
IDH1 et IDH2 Isocitrate Deshydrogenase 1 et 2
IFN Interféron
ITD Internal Tandem Duplication
IT-HSC Intermediate-Term Hematopoietic Stem Cell
JNK c-Jun N-terminal Kinases
KLF-4 Küppler like factor-4
KO Knock-out
LDA Limit Dilution Assay
LDAC Low Dose ArabinosylCytosine
LLA Leucémie Lymphoblastique Aiguë LMA Leucémie Myéloïde Aiguë
LMC Leucémie Myéloïde Chronique
LMMC Leucémie Myelo-Monocytaire Chronique LMPP Lymphoid-primed multipotent progenitor
LPS Lipopolysaccharide
LSD1 Lysine Specific Demethylase-1
LSK Lineage- Sca+ c-Kit+
Ly6C Lymphocyte antigen 6 complex locus C1
Ly6G Lymphocyte antigen 6 complex locus G6D
LT-HSC Long-Term Hematopoietic Stem Cell
MAPK Mitogen Activated Protein Kinases
M-CSF Macrophage- Colony Stimulating Factor
M-CSFR Macrophage- Colony Stimulating Factor Receptor
MDSC Myeloid Derived Suppressive Cells
MEC Matrice ExtraCellulaire
MEIS1 Meis homeobox 1
MEP Megakayocyte/Erythrocyte Progenitor
MIP Macrophage Inflammatory Protein
MLL Mixed Lineage Leukemia
MPP MultiPotent Progenitor
MPO Myélopéroxydase
MRP Myeloid Related Protein
MSCV Murine Stem Cell Virus
Mu-14 Murine 14kDa
MyD88 Myeloid Differentiation primary response gene 88
NADPH Nicotinamide adénine dinucléotide phosphate
NET Neutrophils Extracellular Trap
NF-ĸB Nuclear Factor Kappa B
NPM1 Nucleophosmine 1
NSG NOD scid Gamma
NUP98 Nucleoporine 98 kDa
OMS Organisation mondiale de la santé PAMP Pathogen Associated Molecular Pattern
PI3K PhosphoInositide Kinase 3
PML-RAR Promyelocytic Leukemia- Retinoid Acid Receptor
PRR Pathogen Recognition Receptor
PU.1 Purine box binding protein.1
RAGE Receptor for Advanced Glycation End Products
RC Rémission complete
ROS Reactive Oxygen Species
RUNX Runt-related transcription factor 1
SEER Surveillance, Epidemiology, and End Results
SLAM Signaling Lymphocyte Activation Molecule
SMD Syndrome myélodysplasique SMP Syndrome myéloprolifératif
SNO S-Nitrosylation
SPI-1 SSFV provirus integration-1
SPPB Short Physical Performance Battery
SR1 StemRegenin 1
SSG S-glutathionylation
ST-HSC Short-Term Hematopoietic Stem Cell
STAT Signal Transducer and Activator of Transcription
TCSH Transplantation de Cellules Souches Hématopoïétiques TET2 Ten Eleven Translocation 2
TLR Toll-Like-Receptor
TNF Tumor Necrosis Factor
TRIF TIR-domain-containing adapter-inducing interferon-β
Remerciements
Je tiens tout d’abord à remercier mes directeurs de recherche les Dr. Frédéric Barabé et Dr. Philippe Tessier de m’avoir accueillie au sein de leur équipe et m’avoir permis de mener à terme ce projet avec confiance et autonomie. J’ai apprécié votre disponibilité et vos conseils pas à pas au long de mon doctorat.
Je tiens également remercier les membres du jury, Dr. Paul Naccache, Dr.Trang Hoang et Dr Maria Fernandes d’avoir accepté de lire et d’évaluer ma thèse.
Je suis également reconnaissante à Mélanie avec qui j’ai commencé ce projet. Son encadrement, sa patiente et ses conseils m’ont permis de gagner en autonomie et de mener à bien ce projet.
Je suis également reconnaissant à tous les membres des deux équipes auxquelles j’ai appartenues, ceux passés et présents à savoir Laurine, Joey, Renata, Julie et Marie-Pier sans oublier notre chère Alma ! Joan je te souhaite bon courage et bonne chance pour la suite de ta thèse. Merci à Anne Bergeron et Nathalie Pagé pour leur professionnalisme et pour la patience dont elles ont fait preuve pour répondre à toutes mes questions. Je souhaiterais également remercier le Dr Martin Pelletier ainsi que son étudiante Asmaa d’avoir partagé leur expertise en métabolisme.
Enfin j’aimerais adresser mes remerciements aux personnes que j’ai eu la chance de côtoyer durant mes années au CRI et qui se reconnaitront. Vous avez fait de ce (long) séjour loin de ma famille un moment agréable, sympathique et enrichissant. Bonne chance à tous pour vos projets et pour la suite !
Avant-propos
Les travaux réalisés dans le cadre de ma thèse visent à étudier le rôle des protéines S100A8 et S100A9 dans les leucémies myéloïdes aiguës (LMA). Cette thèse se décline en quatre chapitres. Au cours du premier chapitre, nous allons aborder les aspects épidémiologiques et étiologiques des leucémies myéloïdes aiguës, faire un état des connaissances actuelles concernant les différents mécanismes aboutissant au développement de cette pathologie et les caractéristiques influençant le pronostic des patients et les traitements actuels. Dans un second temps, la famille des protéines S100 sera rapidement présentée suivie d’une description plus poussée de la famille des calgranulines et de leurs implications en situation physiologique et pathologique.
Dans le deuxième chapitre, l’état actuel des connaissances sur les rôles des protéines S100A8 et S100A9 dans les leucémies myéloïdes aigües ainsi que les enjeux de cette étude seront présentés. Par la suite, l’hypothèse de recherche, les objectifs ainsi que l’approche envisagée pour répondre à ces questions seront présentés.
Le troisième chapitre sera consacré aux travaux réalisés au cours de cette thèse sous forme d’article tel qu’il a été publié dans le journal Blood. Cet article est intitulé: “S100A9 induces differentiation of acute myeloid leukemia cells through TLR4”. Le projet de recherche qui a conduit à cet article a été élaboré et dirigé par les Dr Fréderic Barabé et Dr Philippe Tessier. Le Dr Mélanie Tardif a également contribué à la démarche scientifique lors du démarrage du projet. Les expériences visant à caractériser le profil bioénergétique des cellules ont été réalisées par Asmaa Lachaab de l’équipe du Dr Martin Pelletier. Le dosage des calgranulines dans le plasma des patients LMA a été réalisé par Marie-Astrid Raquil. Une partie des lignées humaines utilisée a été fournie par Laurine Gil. J’ai conçu les protocoles et réalisé l’ensemble des expériences in vivo et in vitro, excepté celles citées plus haut. J’ai rédigé le manuscrit, réalisé les analyses statistiques et les figures (principales et supplémentaires) sous la direction des Dr Philippe Tessier et Dr Frédéric Barabé.
Enfin, le dernier chapitre de ce manuscrit de thèse sera consacré à la discussion des résultats obtenus et de leurs portées dans le contexte des LMA et se terminera par une
Chapitre 1 : Introduction
1.1. Leucémies myéloïdes aiguës
1.1.1. Généralités sur les leucémies myéloïdes aiguës
1.1.1.1. Généralités et définitions
Les leucémies myéloïdes aiguës (LMA) constituent un groupe hétérogène d’hémopathies se caractérisant par une prolifération clonale de cellules hématopoïétiques immatures engagées vers la lignée myéloïde [1]. Les cellules myéloïdes (les neutrophiles, les monocytes, les plaquettes ou encore les érythrocytes), générées au cours de l’hématopoïèse ayant lieu dans la moelle osseuse, assurent des fonctions vitales pour l’organisme telles que la réponse immunitaire, la coagulation et le transport d’oxygène. Au cours de la leucémogenèse, les cellules hématopoïétiques acquièrent des anomalies génétiques et épigénétiques qui altèrent leurs propriétés fondamentales de survie, de différenciation et de prolifération [2]. Durant cette transformation, les cellules subissent une évolution génétique et épigénétique ; de ce phénomène naît une diversité clonale à l’origine de l’hétérogénéité des cellules leucémiques et des profils cliniques variés caractérisant les LMA [3, 4]. Ces cellules hématopoïétiques malignes (blastes) arrêtées à un stade précoce au cours du processus normal de différenciation et ayant acquis une capacité proliférative élevée, vont affecter la production des cellules sanguines spécialisées fonctionnelles. Ces dérèglements qui sont reconnaissables sur le plan clinique se manifestent par des symptômes tels que de la fatigue, des saignements ou encore de fréquentes infections. Cet appauvrissement en cellules sanguines fonctionnelles provoque chez les patients souffrants de LMA une susceptibilité accrue aux infections et des complications hémorragiques qui peuvent s’avérer fatales. Parallèlement, ces blastes incapables d’exercer leurs fonctions s’accumulent dans la moelle osseuse et gagnent par la suite la circulation sanguine. Plus rarement, ils peuvent infiltrer d’autres organes (système nerveux central et poumon entre autres). Selon l’Organisation mondiale de la santé (OMS), le diagnostic de LMA est établi lorsque l’infiltration médullaire ou sanguine des blastes est supérieure à 20 % [5]. En dépit de l’existence des traitements chimiothérapeutiques actuels, le pronostic des patients souffrants de LMA reste réservé dans la plupart des cas et est déterminé principalement par les anomalies génétiques constituant la leucémie. L'incidence des LMA est plus prononcée chez les personnes
âgées de plus de 65 ans. La survie au-delà de 5 ans suivant la prise en charge thérapeutique des sujets âgés atteint difficilement 10-20 % faisant de la LMA l’un des cancers les plus agressifs [6, 7]. Il est par conséquent indispensable d’améliorer les traitements actuels qui ont peu évolué ces 30 ans dernières années. Le progrès des technologies de séquençage a permis d’identifier de nouvelles anomalies impliquées dans la leucémogenèse et a stimulé le développement de nouvelles approches thérapeutiques plus ciblées. Ces traitements plus spécifiques commencent à voir le jour, mais les bénéfices sur la survie des patients leucémiques restent encore à être déterminés.
1.1.1.2. Épidémiologie
Les LMA sont des maladies relativement rares, environ 1200 nouveaux cas sont recensés annuellement au Canada (www.cancer.ca) [8]. Elles demeurent néanmoins les leucémies aiguës les plus communes chez l’adulte et représentent 80 % des leucémies aiguës diagnostiquées. Cette pathologie peut survenir à tout âge, on note toutefois un léger pic d’incidence chez l’enfant âgé de moins d’un an et un autre, beaucoup plus prononcé chez les adultes au-delà de 60 ans. L’âge médian au moment du diagnostic est estimé entre 65 et 69 ans. Les personnes âgées sont donc principalement concernées par les LMA avec une incidence cinq à dix fois plus élevée comparativement aux personnes en dessous de 60 ans telle qu’illustrée sur la figure 1 [1, 7].
Figure 1 : Évolution de l’incidence des leucémies myéloïdes aiguës en fonction de la classe d’âge.
Cas répertoriés durant la période allant de 2007 à 2011. Adapté de la revue de Walter & Estey publié en
In ci d en ce ( p o u r 10 0 00 0) Age (Année)
Total
Total Femme Homme1.1.1.3. Étiologie
L’étiologie des LMA n’est pas totalement claire, cependant certains facteurs de risque environnementaux tels que l’exposition à des agents toxiques ou encore des prédispositions propres aux patients ont été identifiés.
1.1.1.3.1. Facteurs étiologiques extrinsèques aux patients
L’un des premiers facteurs reconnus pour accroître la fréquence des leucémies aiguës est l’exposition excessive et chronique au benzène. Les effets leucémogènes du benzène seraient attribués aux produits génotoxiques issus lors de sa métabolisation et qui provoqueraient des anomalies génétiques dans les cellules hématopoïétiques perturbant ainsi l’hématopoïèse [10]. Le tabagisme étant une source d’exposition au benzène, il augmenterait de 1.2 à 2.3 fois le risque de développer une LMA [11].
Le rôle des radiations ionisantes a également été démontré chez les survivants des bombes atomiques. Certaines anomalies chromosomiques (sur les chromosomes 5 et 7) sont retrouvées plus fréquemment chez les personnes ayant été exposées à de fortes radiations et ayant survécu aux bombes atomiques (Hiroshima et Nagasaki) que chez celles nées avant 1945 et n’ayant pas été exposées aux radiations [12].
Environ 10 à 20 % des LMA surviennent à la suite d’une exposition à des agents cytotoxiques ou des radiations utilisées afin de traiter des patients initialement atteints de tumeurs solides [13]. Ces leucémies sont alors dites secondaires par opposition aux LMA diagnostiquées sans antécédents médicaux (LMA de novo ou primaire), une catégorie leur est consacrée dans la classification des LMA par l’OMS. Généralement, les LMA secondaires ont un mauvais pronostic avec une survie plus courte que pour les patients atteints de LMA de novo [14]. Selon le type et la dose de l’agent chimiothérapeutique administré, deux types de LMA secondaires peuvent se manifester. Les différences résident dans le temps de latence et des caractéristiques cytogénétiques des LMA. Le premier type qui se trouve être le plus répandu est le résultat de l’exposition des sujets à des agents alkykants tels que le melphalan, le cyclophosphamide et/ou de la radiothérapie [15]. Le second type de LMA secondaire, quant à lui, survient à la suite de l’administration d’agents inhibant la topoisomérase II (étoposide, teniposide et anthracyclines).
1.1.1.3.2. Facteurs étiologiques intrinsèques aux patients
Certains désordres congénitaux tels que le syndrome de Down (trisomie 21), le syndrome de Bloom, l’anémie de Fanconi ou encore la neurofibromatose prédisposent au développement des LMA, retenons néanmoins que cela ne représente qu’une faible proportion des leucémies [16]. Les enfants atteints de la trisomie 21 ont 10 à 20 fois plus de risques de développer une leucémie lymphoblastique aiguë (LLA) ou une LMA (principalement de type mégakaryocytaire) [17].
Certaines hémopathies peuvent également évoluer en LMA. On peut citer les syndromes myeloprolifératifs (SMP), plus souvent retrouvés chez les patients âgés (> 70 ans) et qui sont caractérisés par une prolifération anormale d’une ou plusieurs lignées cellulaires hématopoïétiques [18, 19]. D’autres formes d’hémopathies peuvent également évoluer en une LMA comme le syndrome myélodysplasique (SMD) qui est le résultat d’anomalies moléculaires affectant la capacité des cellules souches et/ou des progéniteurs hématopoïétiques à se différencier et créant un appauvrissement de cellules hématopoïétiques matures et fonctionnelles [18, 20].
Néanmoins, dans la large majorité des cas, les LMA surviennent sans aucun facteur étiologique identifiable.
1.1.2. Leucémogenèse
1.1.2.1. Hématopoïèse physiologique
Les cellules sanguines exercent des fonctions vitales pour l’organisme, car elles permettent le transport de l’oxygène (érythrocytes), la coagulation (plaquettes) et assurent les défenses contre les infections auxquelles il est confronté (leucocytes). L’hématopoïèse est un processus hautement régulé et hiérarchisé assurant la production continue des cellules sanguines (cellules de la lignée lymphoïde, la lignée myéloïde et la lignée érythroïde). Ce processus prend place dès l’embryogenèse dans le foie fœtal, et se poursuit durant toute la vie adulte dans la moelle osseuse (MO). Compte tenu de la demi-vie très courte de certaines cellules sanguines matures, elles doivent constamment être renouvelées afin de répondre au besoin de l’organisme [21]. Ainsi, environ 100 milliards de cellules hématopoïétiques peuvent être produites quotidiennement dans la moelle osseuse d’un adulte. L’ensemble des cellules hématopoïétiques
cellules possèdent des propriétés remarquables, elles ont la capacité de s’auto-renouveler afin de maintenir constant leur nombre et, grâce à leur propriété de multipotence, elles sont capables de se différencier en tous les types cellulaires hématopoïétiques (Figure 2). Par conséquent, une perturbation des propriétés et/ou de l’activité des CSH peut avoir des répercussions considérables sur l’ensemble de l’hématopoïèse. Outre les CSH qui restent relativement rares dans la moelle osseuse (0.01% à 0.001% chez la souris et 0.01 à 0.2% des cellules hématopoïétiques chez l’humain) [22, 23], le microenvironnement médullaire est également un acteur important pour le bon déroulement de l’hématopoïèse [24]. Celui-ci, également nommé « niche hématopoïétique », fournit un soutien structural et régule la localisation, la fonction et le devenir des CSH [25].
Les cellules hématopoïétiques sont organisées de manière hiérarchisée avec au sommet de la pyramide les cellules souches et à la base les cellules sanguines matures comme l’illustre la figure 2. En fonction de leurs capacités d’autorenouvellement et de différenciation, on peut distinguer trois compartiments. Le premier compartiment est celui des cellules les plus immatures, il est composé des cellules souches multipotentes avec une haute capacité à s’auto-renouveler et peu prolifératives. Le second compartiment est hétérogène et comprend les progéniteurs multipotents MPP (Multipotent Progenitors), les progéniteurs communs à la lignée lymphocytaire, monocytaire et granulocytaire LMPP (Lymphoid primed Mulitpotent Progenitor) et ceux propres à chaque lignée. Pour la lignée myéloïde, il s’agit des CMP (Common Myeloid Progenitors), des GMP (Granulocyte Macrophage Progenitors) et des MEP (Megacaryocyte Erythrocyte Progenitors) et enfin ceux de la lignée lymphoïde CLP (Common Lymphoid Progenitors) et CDP (Common Dendritic cells Progenitors) avec une capacité de prolifération importante et un potentiel d’autorenouvellement et de mutipotence réduit. Enfin, le dernier est composé des précurseurs hématopoïétiques engagés dans une lignée ainsi que les cellules sanguines matures et spécialisées sans potentiel d’autorenouvellement et de multipotence et avec un faible potentiel prolifératif. Pour la lignée myéloïde, cela correspond aux érythrocytes, monocytes, plaquettes, granulocytes, macrophages, et cellules dendritiques.
Figure 2 : Schéma simplifié de l’hématopoïèse.
Les CSH se différencient en progéniteurs myéloïdes/ lymphoïdes communs (CMP et GMP). Les progéniteurs vont s’engager vers la lignée érythrocytaire et plaquettaire ; MEP (Megakaryocytes Erythrocyte Progenitors), granulocytaire ; GMP (Granulocyte-Macrophage Progenitors) et donner des précurseurs (non illustré). Ces précurseurs vont à leur tour être enrôlés dans les stades finaux de différenciation pour donner des cellules matures des grandes lignées (lymphoïde et myéloïde). Schéma tiré et adapté de la revue Manz & Boettcher datant de 2014 [26].
1.1.2.1.1. La niche hématopoïétique
Le devenir des cellules hématopoïétiques, à savoir la survie, la prolifération ainsi que la différenciation, est dépendant du microenvironnement médullaire où elles sont logées. En 1978, Schofield fut le premier à introduire le concept de « niche hématopoïétique » comme étant une région dans la moelle osseuse régulant l’hématopoïèse [27]. La niche hématopoïétique est un microenvironnement complexe dans la moelle osseuse composé de divers types cellulaires (cellules endothéliales, cellules stromales mésenchymateuses, cellules hématopoïétiques et cellules nerveuses) [25] (Figure 3). Les cellules stromales présentes dans la niche sécrètent des facteurs qui vont composer la matrice extracellulaire (MEC) et réguler l’adhérence, le trafic,
constituée de glycoprotéines (collagène de type I et II, lamine et fibronectine), des glycosaminoglycanes (acide hyaluronique et de sulfate d’héparine et de chondroïtine) et métalloprotéase (MMP). Parmi les facteurs sécrétés influençant directement ou indirectement la fonction des CSH on peut citer : l’angiopoïétine-1, l’ostéopontine, SDF-1 (Stromal-Derived Factor-1) et le SCF.
Figure 3 : Les différents types de cellules et facteurs présents au sein de la niche hématopoïétique.
Le système vasculaire au sein de la niche est un acteur clé du maintien des CSH dans la moelle osseuse. Les CSH dormantes sont retrouvées autour des artérioles où des facteurs tels le CXCL12 et le SDF1, qui sont sécrétés par les cellules périvasculaires, les cellules endothéliales, les cellules réticulaires dites CAR (CXCL12-Abundant Reticular) de Schwann et les cellules nerveuses du système sympathique, assurent leur maintien dans la moelle osseuse. Les CSH sont maintenues dans un état quiescent par le transforming growth factor beta (TGF-β) et la thrombopoïétine (TPO) produits par les cellules nerveuses et les ostéoblastes respectivement. Les CSH activées sont localisées près des niches sinusoïdes où leurs propriétés d’autorenouvellement, de prolifération et de différenciation sont régulées. Les cellules hématopoïétiques telles que les macrophages ou les mégacaryocytes dérivés des CSH peuvent influencer à leur tour leur migration et leur prolifération. GFAP : Protéine acide fibrillaire gliale ; LepR : récepteur leptine. Adapté de la revue de Boulais et Frenette parue en 2015 [28]. Les cytokines et les facteurs de croissance du système hématopoïétique sont également des médiateurs ou des déclencheurs de signaux influençant positivement ou négativement la survie, la prolifération et la différenciation des cellules hématopoïétiques. Ils comprennent entre autres
les interleukines 3, 6,11 et 7, l’érythropoïétine (EPO), la thrombopoïetine (TPO) et le SCF (Stem Cell Factor). De plus, des voies de signalisations telles que Wnt, NOTCH, Hedgehog, BMP et TGF-β influencent également le devenir des CSH (Figure 3). Grâce à l’avancée des techniques d’analyse de cytométrie en flux, de tri cellulaire et d’imagerie in vivo, les connaissances concernant la niche hématopoïétique et la distribution des CSH ont considérablement évolué. Chez l’humain, les sites d’hématopoïèse se trouvent dans les os plats tels que les os de la hanche ou l’os du sternum et dans les épiphyses des os longs (fémur). L’étude de la localisation des CSH a révélé qu’elles se trouvaient préférentiellement au niveau de l’endostéum (interface entre les longs os et la MO) et proches des cellules périvasculaires (cellules entourant les vaisseaux sanguins) dans une région plus centrale de la moelle osseuse. Ces deux niches coopèrent et peuvent héberger deux types de CSH fonctionnellement distincts [29, 30]. La niche endostéale composée de cellules endothéliales, de fibroblastes et d’ostéoclastes régulerait le maintien de la quiescence des CSH avec un potentiel de reconstitution à long terme. La niche vasculaire quant à elle, composée de cellules périvasculaires, d’adipocytes, de cellules endothéliales et de progéniteurs mésenchymateux, supporterait les CSH mobilisables en quelques minutes en cas de stress et lors du phénomène de mobilisation (passage des CSH dans la circulation sanguine).
1.1.2.1.2. Les cellules hématopoïétiques
• Les cellules souches hématopoïétiques (CSH)
Les CSH furent initialement identifiées au cours d’une étude menée par Till et McCulloch comme une population clonale de cellules de la moelle osseuse capable de générer des colonies multi-lignages dans la rate de souris irradiées [31]. Bien qu’elles ne représentent qu’une infime partie des cellules de la moelle osseuse, elles sont en mesure de produire l’ensemble des cellules du système hématopoïétique, grâce à leurs propriétés fondamentales d’auto-renouvellement et leur multipotence. Au sein de la niche, la majorité d’entre elles existent dans un état quiescent (~75%) ce qui leur permet de se protéger des stress micro-environnementaux et de minimiser l’accumulation de mutations dans le but de préserver leur intégrité et d’assurer le processus d’hématopoïèse, et ce, tout au long de la vie de l’individu [32-34]. Deux populations de CSH se distinguant par des cinétiques de divisions différentes ont été identifiées ; l’une des
période de renouvellement de 21 semaines [35]. Par ailleurs, on note qu’avec le vieillissement, les propriétés de multipotence et de prolifération sont altérées [36]. Avec l’âge, il a été observé chez la souris que les CSH présentent un potentiel prolifératif supérieur accompagné d’une tendance à générer des cellules de la lignée myéloïde comparée aux plus jeunes cellules. Cela pourrait en partie expliquer cette susceptibilité aux développements de pathologies telles que les leucémies [37]. Des mécanismes extrinsèques et intrinsèques œuvrent conjointement afin de réguler l’état des CSH à savoir leur entrée en cycle cellulaire (division) ou non (quiescence) qui sont des propriétés essentielles à la prospérité du système hématopoïétique. En fonction des stimuli reçus, les CSH vont entrer en division. Lorsque les cellules entrent en cycle cellulaire, deux modes de divisions peuvent alors être envisagés (Figure 4) [38].
Figure 4: Modèle de division des cellules souches hématopoïétiques (Division asymétrique vs symétrique).
S : Cellule souche ; D : cellule différenciée.
Dans le cas d’une division symétrique, la CSH donne naissance à deux cellules filles aux propriétés identiques à la CSH mère, il en découle alors une expansion du nombre de CSH tel qu’illustré sur la figure 4A. Généralement, ce type de division se produit au cours du développement embryonnaire ou lors de greffes afin de reconstituer le système hématopoïétique, de rétablir l’homéostasie et la réserve de CSH. Dans le second cas, une CSH se divise de façon asymétrique et donne naissance à une cellule fille avec des propriétés identiques à la cellule mère et une autre cellule fille destinée à se différencier (Figure 4 B). Ce type de division favorise ainsi un maintien constant de la réserve de CSH et en condition normale, il est présumé que ce mode de division homéostatique est utilisé. Toutefois, dans la situation où les deux cellules filles ont un destin différent de celui de la cellule-mère et se
différencient (Figure 4 C), le réservoir de CSH peut rapidement s’épuiser. L’orientation vers l’un ou l’autre de ces modes de division peut être influencée par des facteurs intrinsèques ou environnementaux présents au sein de la niche [39].
En dépit de la rareté de ces cellules, l’utilisation de nouveaux outils d’imagerie a permis d’identifier des hétérogénéités phénotypiques et fonctionnelles au sein de cette population. Seuls les LT-HSC (Long Term - Hematopoietic Stem Cell) sont capables d’assurer l’hématopoïèse durant la vie d’un individu (au-delà de 6 mois). À l’inverse, les ST-HSC (Short Term Hematopoietic Stem Cell) ne peuvent reconstituer l’hématopoïèse que pendant une durée limitée de 4-8 semaines [40-42]. Récemment, une autre population avec une capacité de reconstitution intermédiaire pouvant aller jusqu’à 8 mois a été identifiée (Intermediate Term-Hematopoietic Stem Cell : IT-HSC) [43, 44]. En raison de leur faible fréquence et de l’absence de marqueurs spécifiques, l’étude des cellules CSH a été ralentie. Leurs identification et purification reposent principalement sur leurs propriétés phénotypiques et cytochimiques. La caractérisation phénotypique des CSH a contribué à la mise en œuvre de techniques basées sur le tri de cellules par cytométrie en flux et l’utilisation de combinaisons de marqueurs permettant d’obtenir une population enrichie en CSH [45]. Chez l’humain, les marqueurs de surface les plus utilisés sont les glycoprotéines CD34 et CD38. Cependant compte tenu de l’hétérogénéité des populations exprimant CD34 et CD38, il est nécessaire d’utiliser des marqueurs supplémentaires qui sont présentés dans le tableau 1 afin de distinguer les cellules souches et les progéniteurs hématopoïétiques [46]. Chez la souris, les CSH sont caractérisées par l’absence de marqueur de lignée (lin-) présent sur les cellules matures tels que le B220, CD11b, Gr-1, CD3 et TER-119 et l’expression des marqueurs de surface tels que la glycoprotéine Sca-1 et le récepteur tyrosine kinase c-Kit (LSK). L’utilisation des molécules de signalisation d’activation lymphocytaires SLAM (Signalling Lymphocyte Activation Molecule) à savoir les marqueurs CD150, CD244 et CD48 associées aux marqueurs LSK a permis d’améliorer les taux d’enrichissement des CSH (Tableau 1) [47]. La définition des CSH repose sur les caractéristiques phénotypiques, mais également fonctionnelles (multipotence et auto-renouvellement) qui sont évaluées à l’aide des méthodes de transplantation in vivo et des tests
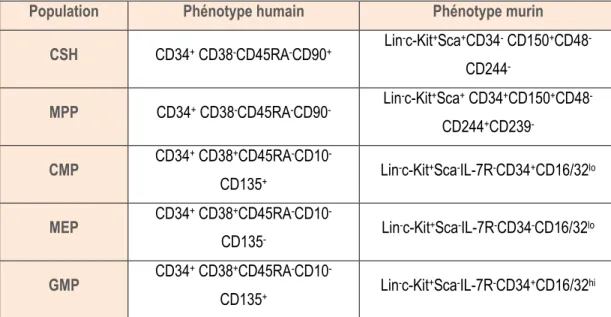
Tableau 1 : Phénotypes des différentes populations de cellules hématopoïétiques selon l’expression des marqueurs de surface.
Population Phénotype humain Phénotype murin
CSH CD34+ CD38-CD45RA-CD90+ Lin -c-Kit+Sca+CD34- CD150+CD48 -CD244 -MPP CD34+ CD38-CD45RA-CD90- Lin -c-Kit+Sca+ CD34+CD150+CD48 -CD244+CD239 -CMP CD34 + CD38+CD45RA-CD10 -CD135+ Lin
-c-Kit+Sca-IL-7R-CD34+CD16/32lo
MEP CD34
+ CD38+CD45RA-CD10
-CD135- Lin
-c-Kit+Sca-IL-7R-CD34-CD16/32lo
GMP CD34
+ CD38+CD45RA-CD10
-CD135+ Lin
-c-Kit+Sca-IL-7R-CD34+CD16/32hi CSH : Cellule Souche Hématopoïétique, MPP : MultiPotent Progenitor, CMP : Common Myeloid Progenitor, MEP : Megakaryocyte Erythroid Progenitor, GMP : Granulocyte-Macrophage Progenitor.
• Les progéniteurs hématopoïétiques
Le compartiment des progéniteurs consiste en une population très hétérogène (Figure 2). Lorsqu’une CSH se différencie, elle va générer des ST-HSC (Short Term- Hematopoietic Stem Cells), qui se différencieront progressivement en progéniteurs multipotents (MPP). Selon les besoins, les progéniteurs multipotents détenant à ce stade un pouvoir de prolifération important vont s’engager vers la voie lymphoïde ou myéloïde en générant respectivement des progéniteurs communs lymphoïdes ou myéloïdes (CLP : Common Lymphoid Progenitor ; CMP : Common Myeloid Progenitor) (Figure 2). À ce stade, ces progéniteurs ont un potentiel de différenciation plus restreint et une capacité d’auto-renouvellement quasi nulle (Figure 2). Alors que le développement des cellules composant la lignée myéloïde se poursuit dans la moelle osseuse, la maturation des cellules lymphoïdes, elle, a lieu dans les organes lymphoïdes à savoir le thymus et la rate pour les lymphocytes T et B respectivement. Les CLP vont alors donner naissance aux progéniteurs des cellules T et B, aux cellules Natural killer (NK) et à certaines sous-populations de cellules dendritiques, alors que les CMP eux, se différencient en progéniteurs des granulocytes et macrophages (Granulocyte-Macrophage Progenitor : GMP) et en progéniteurs des mégacaryocytes et érythrocytes (MEP : Megakaryocyte/Erythroid Progenitor). Le modèle hiérarchique conventionnel des CMP et CLP a été quelque peu bousculé
par la découverte des progéniteurs LMPP in vitro (Lymphoid-Primed Multipotent Progenitor) [48]. Ces derniers peuvent générer des progéniteurs de cellules lymphoïdes, monocytaires et granulocytaires, néanmoins ils ne peuvent pas donner naissance aux progéniteurs mégacaryocytaires et érythroïdes. Le modèle selon lequel les progéniteurs constituent une population uniforme capable de générer les différentes cellules de la lignée myéloïde à savoir les granulocytes, les monocytes, les érythrocytes et les mégacaryocytes à des degrés équivalents se complexifie. De récentes études décrivent une hétérogénéité au sein des cellules progénitrices, celles-ci seraient « primed » et déjà prédéterminées à générer certaines lignées [49, 50]. Les progéniteurs ne peuvent être discriminés morphologiquement, mais l’utilisation de marqueurs de surface permet leur l’identification par cytométrie en flux (Tableau 1).
• Les précurseurs hématopoïétiques
Les précurseurs hématopoïétiques sont des cellules engagées dans le processus de maturation terminale et sont identifiables morphologiquement et phénotypiquement. Ces cellules ont commencé à acquérir progressivement les fonctions propres à la lignée à laquelle elles appartiennent. Une fois le processus de maturation achevé, les cellules vont gagner la circulation sanguine afin d’assurer leurs fonctions.
Les leucémies myéloïdes aiguës résultent d’un dérèglement du processus normal de la myélopoïèse. Pour ces raisons, dans ce manuscrit de thèse, je me concentrerai sur les acteurs physiologiques de la différenciation myélomonocytaire.
1.1.2.1.3. La granulopoïèse neutrophile
Les neutrophiles, cellules de l’immunité innée représentent les premières lignes de défense de l’organisme. Elles exercent des fonctions essentielles durant l’inflammation et lors d’infections [51]. Ce sont les premières cellules à être recrutées sur le site infectieux (quelques heures) où elles assurent la destruction des agents pathogènes. Pour ce faire, les neutrophiles utilisent de multiples mécanismes incluant la phagocytose, la sécrétion de cytokines, le relargage d’enzymes antimicrobiennes, la production d’espèces réactives de l’oxygène (ROS ; Reactive Oxygen Species) ou encore des pièges extracellulaires composés de matériel nucléaire (NET ; Neutrophil Extacellular Trap) [52]. Lorsque le danger a été neutralisé, il s’en suit une phase de
résolution de l’inflammation durant laquelle les neutrophiles sont éliminés par les macrophages tissulaires présents sur le site inflammatoire.
Les neutrophiles humains sont les cellules très abondantes dans le sang. Elles constituent entre 50 % et 70% des leucocytes circulants (de 7 % à 8 % chez la souris). La demi-vie d'un neutrophile non activé dans la circulation sanguine n’est que de quelques heures (~ 7 heures voir un peu plus de 5 jours) [53]. Par conséquent, une production de neutrophile à grande échelle est nécessaire, elle représente une part considérable de l’activité de la moelle osseuse en condition homéostatique. Ainsi, de 1010 à 1011 neutrophiles doivent être générés dans la moelle osseuse quotidiennement chez l’adulte [54, 55].
Figure 5 : Les étapes de maturation du neutrophile.
Au stade myéloblastique, promyélocytaire et myélocytaire, les précurseurs ont une forte capacité proliférative. Par la suite, au cours de la maturation en métamyélocytes et en précurseur de neutrophile non segmenté, l’activité proliférative diminue pour devenir nulle une fois mature. Lorsque la maturation est complète, le neutrophile peut quitter la moelle osseuse et gagner la circulation sanguine. Schéma adapté de la revue de Coffelt et al. parue en 2016 [56].
La différenciation terminale des neutrophiles comprend différentes étapes successives reconnaissables morphologiquement (taille, morphologie du noyau, présence de granules, etc.) (Figure 5) [57, 58]. Durant la maturation, le volume des cellules diminue et les granules
commencent à apparaître. Les granules primaires et secondaires composées d’entre autres de la myélopéroxydase (MPO), de la cathépsine, et de l’élastase et de la lactoferrine se forment aux stades promyélocytes et myélocytes respectivement. Aux stades suivants de maturation, les précurseurs myélocytes-métamyélocytes acquièrent les granules tertiaires riches en gélatinase et les vésicules sécrétoires. À ces stades, les précurseurs de neutrophiles arrêtent de proliférer. Les neutrophiles se distinguent phénotypiquement grâce aux marqueurs exprimés à leur surface tels que le CD11b, le CD16 et le CD15 chez les neutrophiles humains, et CD11b et Gr-1 sur les neutrophiles murins (Figure 5). Une fois la maturation achevée, ces cellules peuvent rester quelques jours (4 à 6 jours) au sein de la moelle osseuse assurant la réserve de neutrophiles rapidement mobilisable lors d’épisodes inflammatoires ou d’infections par exemple [59]. La libération des neutrophiles de la moelle osseuse vers la circulation est en partie régulée par les signaux extérieurs comme les cytokines, les chimiokines ou la présence d’agents pathogènes. La mobilisation des neutrophiles est assurée par l’axe de signalisation CXCL12/CXCR4 (favorisant la rétention) et CXCL2/CXCR2 (favorisant la mobilisation) [60-62]. Par exemple, le facteur G-CSF (Granulocyte Colony Stimulating Factor) stimule la sortie des neutrophiles de la moelle osseuse en agissant directement sur les neutrophiles ou indirectement en modulant l’expression de CXCL12 (diminution) ou CXCL2 (augmentation). Les neutrophiles non activés seront éliminés après quelques heures passées dans la circulation par phagocytose par les macrophages dans la rate, le foie ou suite à leur retour dans la moelle osseuse causé par l’augmentation de CXCR4 [63].
Un dérèglement de la production de neutrophile ou de leurs fonctions peut avoir des conséquences assez importantes. En raison de leur rôle central dans la réponse immunitaire innée, une forte susceptibilité aux infections est observée dans des conditions de neutropénie. À l’inverse, une production ou une activité excessive des neutrophiles peut être néfaste pour l’organisme et engendrer des dégâts tissulaires. Pour toutes ces raisons, la granulopoïèse est un processus qui est hautement régulé par différents acteurs incluant une panoplie de cytokines et de facteurs de transcription.
• Les facteurs de transcription régulant la différenciation granulocytaire Les bouleversements qui découlent des anomalies affectant certains facteurs de transcription
neutrophiles est influencée par un panel de facteurs de transcription dont les principaux seront présentés ci-dessous [64] (Figure 5 et 6).
- -La famille des facteurs CCAAT/Enhancer Binding Protein (C/EBP)
Les facteurs de transcription de la famille C/EBP comprenant C/EBP-α, C/EBP-β, C/EBP-δ et C/EBP-ε sont déterminants dans la régulation du développement de la lignée myéloïde [65-67]. Ils assurent le contrôle de la différenciation, de la prolifération et de la survie cellulaire des cellules myéloïdes.
Le facteur C/EBP-α est exprimé par différents types cellulaires. Toutefois, dans le compartiment hématopoïétique, il est préférentiellement retrouvé dans les progéniteurs myéloïdes (CMP et GMP) et les granulocytes, monocytes et éosinophiles, mais pas dans les macrophages [68, 69]. C/EBP-α agit dès les premières étapes de différenciation myélomonocytaire et facilite la transition de CMP à GMP et, la formation des GMP est fortement diminuée en son absence [70, 71]. L’inactivation conditionnelle du gène cepba dans les GMP n’affecte pas la production de granulocytes. Cela suggère que C/EBP-α ne serait pas indispensable dans les stades finaux de différenciation granulocytaire (au-delà du stade GMP) [72]. La production de granulocytes semble cependant être dépendante du facteur C/EBP-α, puisqu'un niveau de C/EBP-α plus élevé est requis en comparaison à la monocytopoïèse. De plus, une diminution du niveau d’expression de C/EBP-α affecte la granulopoïèse, mais non la monocytopoïèse ce qui implique que ce facteur jouerait un rôle instructif et favoriserait l’engagement des GMP vers la lignée granulocytaire [73-75]. Son action réside principalement dans l’activation de l’expression d’autres régulateurs de la maturation des granulocytes tels que C/EBP-ε, Gfi-1 (Growth factor independence 1) et KLF5 (Küppler like factor-5) [73, 76, 77]. Conjointement avec PU.1,c-myb et RUNX1 (Runt-related transcription factor 1), il active l’expression de gènes codants pour des enzymes impliquées dans la fonction des neutrophiles, parmi elles, la myéloperoxidase, l’élastase, le lysozyme, la lactoferrine ou encore des récepteurs de facteurs de croissance (G-CSF et GM-(G-CSF) [78, 79]. Durant la transition du stade promyélocyte à celui de myélocyte, une diminution de la capacité proliférative a lieu. Cet effet est en partie régulé par le facteur C/EBP-α, lequel lie la protéine E2F et prévient le passage du stade G1 à S en réprimant l’activation des gènes impliqués dans la progression du cycle cellulaire [80].
Le facteur C/EBP-ε quant à lui voit son expression augmenter dans les stades plus tardifs de différenciation myéloïde et son absence entraîne une différenciation incomplète des granulocytes (Figure 5) [81-83]. Ce facteur étant un régulateur clé des gènes codants pour les protéines constituant les granules secondaires, la diminution de son expression affecte la fonction des granulocytes [83-85]. De manière similaire à C/EBP-α, C/EBP-ε favorise l’arrêt de la prolifération cellulaire lors du passage au stade myélocytaire en favorisant l’expression de la CKI (Cyclin dependant Kinase Inhibitor) p27kip1 et en inhibant l’expression des CDK4 (Cyclin Dependant Kinase), CDK6, cyclent A et cyclent D2 [86, 87].
Au vu de leurs importances dans la différenciation myéloïde, les facteurs de transcription de la famille C/EBP peuvent être impliqués dans la leucémogenèse. En effet, les anomalies impliquant CEBPA sont présentes dans 5 à 10% des leucémies et occasionnent un défaut dans la maturation des cellules hématopoïétiques. Nous en rediscuterons plus en détail dans la section pathogenèse des LMA.
-Growth factor independence 1 (Gfi1)
Ce facteur de transcription en doigt de zinc est exprimé par les CSH, les neutrophiles et les lymphocytes aux stades précoces de la maturation, mais pas dans les lignées mégacaryocytaires et érythrocytaires [88-90]. Le facteur Gfi-1 a un rôle pléiotropique dans l’hématopoïèse et est requis lors des stades finaux de différenciation granulocytaire (au-delà de la phase promyélocytaire). Les souris déficientes en Gfi-1 ont un nombre normal de GMP. Cependant, on constate qu’elles possèdent des promyélocytes anormaux alors que les neutrophiles sont eux absents [91, 92]. Par ailleurs, comme de nombreux facteurs de transcription impliqués dans le choix de la lignée, Gfi-1 agit d’une part comme activateur de la maturation granulocytaire en induisant entre autres l’expression du récepteur au G-CSF (Granulocyte-Colony Stimulating Factor) et des acteurs de la signalisation en aval (ex.RasGRP1) [93], et d’autre part, Gfi-1 inhibe la différenciation monocytaire par la répression du facteur de transcription PU.1 [94, 95].
• Cytokines et facteurs de croissance régulant la différenciation granulocytaire Le microenvironnement (niche) dans lequel les cellules hématopoïétiques sont logées est un acteur majeur de la régulation de l’hématopoïèse du fait des signaux induits par les interactions
la maturation, les cellules acquièrent progressivement à leur surface certains récepteurs leur permettant d’être plus sensibles aux signaux extérieurs comme les cytokines ou encore les facteurs de croissance.
-G-CSF et GM-CSF (Granulocyte/ Grarnulocyte-Macrophage Colony Stimulating Factor)
Le GM-CSF et le G-CSF ont un rôle central dans la régulation de la prolifération et de la différenciation des cellules hématopoïétiques. Ce sont des glycoprotéines sécrétées par les cellules hématopoïétiques elles-mêmes incluant les macrophages et les granulocytes ainsi que par les cellules présentes dans la moelle osseuse (cellules stromales, fibroblastes, ostéoblastes, cellules endothéliales, etc.). L’expression du récepteur du G-CSF (G-CSFR) augmente graduellement au cours de la différenciation jusqu’à atteindre des niveaux importants dans les précurseurs des neutrophiles et les neutrophiles matures amplifiant leur sensibilité au facteur de croissance G-CSF (Figure 5) [96-98]. Le G-CSF induit la survie et la prolifération des précurseurs granulocytaires et joue un rôle décisif dans le choix de la lignée [99-102]. À la suite de la liaison du G-CSF sur son récepteur, diverses voies de signalisations intracellulaires sont activées. Les voies JAK/STAT-1-3-5 (Signal Transducer and Activator of Transcription 1-3-5), Ras/ ERK (Extracellular Signal–Regulated Kinases) et Akt/PKB (Serine Threonine tyrosine kinase/Protein Kinase B) sont induites, néanmoins le signal prépondérant est relayé par STAT5 et STAT3 [103-107]. Contrairement au G-CSF, le GM-CSF est peu exprimé à l’état basal et est synthétisé en situation de stress. D’ailleurs, il n’est pas indispensable à la myélopoïèse en condition homéostatique [108]. Le GM-CSF active principalement ces trois voies, JAK (Janus Kinase)-STAT, MAPK (Mitogen-activated protein kinases) et PI3K (PhosphoInositide Kinase 3) [109]. En raison de leur capacité à induire la production des cellules myéloïdes, ces facteurs de croissance ont une utilité thérapeutique très intéressante. Leur utilisation permet d’éviter les risques d’infection lors de neutropénies qui peuvent être causées par l’utilisation d’agents cytotoxiques (traitement chimiothérapeutique par exemple) [110, 111].
1.1.2.1.4. La monocytopoïèse
Les monocytes sont des cellules participant aux fonctions essentielles de surveillance immunitaire aussi bien en situation homéostatique que durant la réponse inflammatoire [112-114]. Les monocytes représentent respectivement ~ 10 % et 2 % des leucocytes circulants chez
l’humain et la souris [115, 116]. La production de monocytes démarre lors de l’engagement des progéniteurs GMP vers les MDP (Monocyte Dendritic cells Progenitor) puis en progéniteur cMoP (Common Monocyte Progenitor). La monocytopoïèse englobe les divers processus menant à la production de monocytes à partir de monoblastes puis successivement en promonocyte et monocyte immature. Elle reste moins bien documentée que la granulopoïèse. Lorsque les cellules sont matures, grâce en partie à la chimiokine CCL2 et son récepteur CCR2, elles vont passer dans la circulation sanguine pour accomplir leurs fonctions [117]. Les monocytes contribuent à la réponse inflammatoire et sont capables de produire rapidement une quantité abondante de cytokines pro-inflammatoires (TNF, IL-1β, IL-6, IL-8) et de l’IL-18 pour recruter les cellules Natural Killer et ainsi amplifier la réponse immunitaire [118]. Par la suite, les monocytes vont intervenir dans la résolution de l’inflammation en sécrétant des cytokines anti-inflammatoires que représentent l’IL-10 et le TGF-β. Le rôle primaire des monocytes a longtemps été considéré comme le réapprovisionnement des macrophages et des cellules dendritiques dans les tissus [119-121]. Il est aujourd’hui largement admis que certains types de macrophages tissulaires dérivent d’une réserve de précurseurs embryonnaires (rate, foie, cerveau, etc.) et non uniquement des monocytes circulants [122-124].
Les monocytes expriment des antigènes de surface permettant leur identification comme le CD11b, le CD14 ou encore le M-CSFR (Macrophage- Colony Stimulating Factor Receptor ; CD115) sur les cellules humaines et CD11b et Ly6C sur les monocytes murins [125]. Les monocytes représentent une population hétérogène, en effet les MDP (Monocytes Dendritic cells Progenitors) peuvent donner naissance à différentes sous-populations de monocytes se distinguant à la fois par l’expression de marqueur de surface, mais également par les propriétés fonctionnelles propres à chacune sous-population [126]. Ainsi, d’après la nomenclature officielle de l’Union International des Sociétés d’Immunologie, trois sous populations monocytaire sont distinguées en fonction de l’expression du marqueur CD14 et CD16 [127]. La première population dite « classique » est majoritaire (~80 %). Ces monocytes ont un fort potentiel phagocytaire et ont le phénotype suivant : CD14hiCD16lo (Ly6Chi CX3CR1lo CCR2hi chez la souris). Les monocytes « intermédiaires » sont discriminables phénotypiquement par l’expression du CD14int et du CD16int, on leur attribue une fonction inflammatoire et activatrice des lymphocytes T. Enfin, les monocytes « non classiques » de phénotype suivant
CD14loCD16hi (Ly6Clo CX3CR1hi CCR2lo chez la souris) eux, jouent un rôle dans la surveillance des tissus [128, 129].
-Du monocyte aux macrophages :
La différenciation ultime des monocytes en macrophages fait suite à leur infiltration dans les tissus, celle-ci est notamment contrôlée par des signaux micro-environnementaux comme des cytokines et chimiokines (p. ex. CCL2). Les macrophages consistent en une population hétérogène tant morphologiquement que métaboliquement et fonctionnellement. Les macrophages acquièrent un phénotype spécifique au tissu dans lequel ils résident. Ces cellules de l’immunité innée représentent une première barrière de défense contre les pathogènes en les phagocytant et en présentant leurs antigènes aux lymphocytes T. D’autre part, les macrophages sont classés en deux catégories : de type M1 avec un profil inflammatoire et de type M2 anti-inflammatoire [130]. Les macrophages M1, caractérisés par une forte activité phagocytaire et une production accrue de cytokines pro-inflammatoires, seraient produits en réponse à la stimulation des monocytes par de l’IFN-γ, TNF et/ ou des produits microbiens comme le lipopolysaccaride (LPS). Les macrophages M2, eux, impliqués dans la réparation des tissus, sont formés en réponse à l’IL-4. Néanmoins, de nombreuses études ont reporté l’existence d’une plasticité des macrophages. Ainsi en fonction des conditions environnementales, la transition d’un type à l’autre est possible dans le but de répondre aux besoins de l’organisme [131-133].
Le rôle des macrophages ne se restreint pas à la protection de l’organisme contre des agents pathogènes, ils interviennent également dans l’homéostasie cellulaire par exemple en phagocytant les granulocytes et érythrocytes en sénescence [134, 135].
-Du monocyte aux cellules dendritiques :
Les cellules dendritiques, cellules présentatrices d’antigène spécialisées, jouent un rôle essentiel d’intermédiaires entre les réponses immunitaires innées et adaptatives. Ces cellules représentent une population hétérogène en termes d’origine, de morphologie et de fonction [136]. Les cellules dendritiques peuvent être générées in vitro à partir de monocyte par la stimulation au GM-CSF et à l’IL-4. La différenciation des monocytes en cellules dendritiques a également été reportée in vivo [137].
• Facteurs de transcription régulant la différenciation monocytaire et
macrophagique
De manière semblable au processus de granulopoïèse, la production de monocyte implique une régulation transcriptionnelle complexe [138, 139] .
-PU.1/SPI-1 (purine box binding protein.1/ SSFV provirus integration-1)
Ce membre des facteurs de transcription de la famille Ets exprimé par différentes lignées hématopoïétiques (principalement granulocytes, monocytes et lymphocytes B) est un régulateur clé de l’hématopoïèse et agit à de multiples étapes de différenciation des cellules hématopoïétiques [140, 141]. Ainsi, lorsque PU.1 est absent plusieurs lignées hématopoïétiques sont perturbées comme les lymphocytes (B et T), les monocytes, les neutrophiles ainsi que les macrophages démontrant toute son importance dans la régulation de l’hématopoïèse [142, 143]. Le facteur de transcription PU.1 agit dès les premières étapes de différenciation hématopoïétique en orientant les progéniteurs CMP vers des GMP au détriment des MEP (Figure 6) [144, 145]. Dans un second temps, PU.1 va intervenir dans l’équilibre entre les granulocytes et les monocytes. L’effet de ce facteur de transcription est dépendant de son niveau d’expression et dès les premières étapes de maturation monocytaire, son expression est augmentée [146]. À l’inverse, des niveaux en PU.1 plus faibles entraînent la formation des précurseurs granulocytaires [147, 148]. PU.1 favoriserait la monocytopoïèse au détriment des lignées granulocytaires et erythro-mégacaryocytaires en réprimant l’activité des facteurs de transcription C/EBP-α et GATA-1 respectivement [148-151]. D’autre part, PU.1 contrôle également l’expression des gènes critiques qui régulent la différenciation et la fonction des monocytes/macrophages en induisant l’expression des récepteurs de facteurs de croissance (GM-CSF et M-CSF) et des facteurs de transcription tels qu’Egr-1 (Early Growth Response-1), IRF-8 (Interferon Regulatory Factor-8) et KLF-4 (Küppler like factor-4) (Figure 6) [152-154].
-Interferon Regulatory Factor-8 (IRF-8)
Membre de la famille des facteurs de transcription IRF, IRF-8 est faiblement exprimé dans les cellules souches et les progéniteurs hématopoïétiques. Son niveau d’expression augmente graduellement dans les progéniteurs communs et granulocyte-macrophage [155]. En collaboration avec PU.1, il induit le développement des monocytes et macrophages et réprime






![Tableau 5 : Stratification cytogénétique des LMA selon les recommandations de l’ELN. Adapté de la revue de Döhner et al., parue en 2010 [5]](https://thumb-eu.123doks.com/thumbv2/123doknet/5557544.133090/57.918.178.793.144.676/tableau-stratification-cytogénétique-recommandations-adapté-revue-döhner-parue.webp)


