Migration des cellules du lignage mélanocytaire
Texte intégral
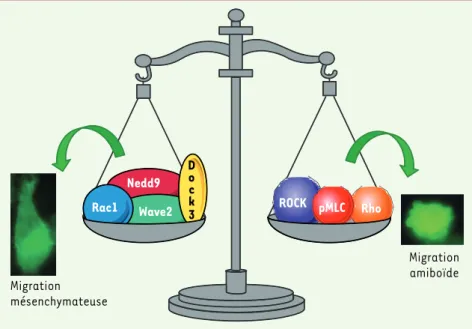
Figure


Documents relatifs
Il s'agit de la détermination de la place et du rôle que prend le sujet dans la définition des contours de l'orientation du devenir de son soi (Kaddouri M., 2002).Le
B — Plus loin page 21 : « De même il est inexact que d'une « manière générale, le Verrucaria maura, du moins si on « ne limite pas ses observations à une seule
In this thesis we will focus on the migration of bladder cancer cells and we will try to understand how the processes presented above can be described in terms of traction stresses
Figure 4 shows the bistatic RCS of a surface made of 1 cosine component, calculated by both the cosine approaches and the “numerical neglected slopes PO” approach, versus
Since the Lapcs problem is NP-complete for arc-annotated sequences of Nested types, so does the last open case for the Edit problem, Edit(Nested, Nested)?. In
De fait, des observations morpholo- giques ultrastructurales révèlent la présence d’annexine II dans une zone de pontage entre les granules et la membrane plasmique lorsque des
Différentes topographies des protéines
En conclusion, nous avons démontré qu'une exposition aux agonistes des PPAR modifie I'expression de protéines impliquées dans I'adhérence et la migration des cellules HT29,
