L’efficacité du CD154 monomérique dans le traitement des complications thrombotiques
Texte intégral
Figure
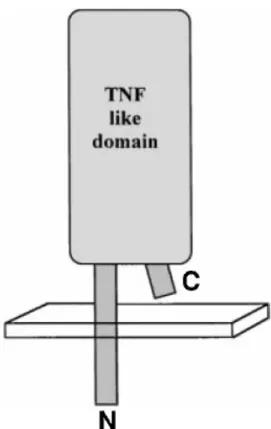
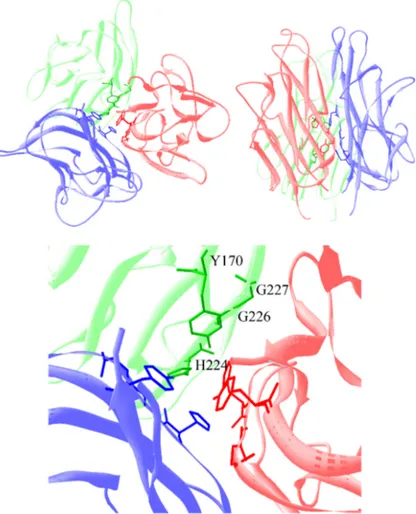
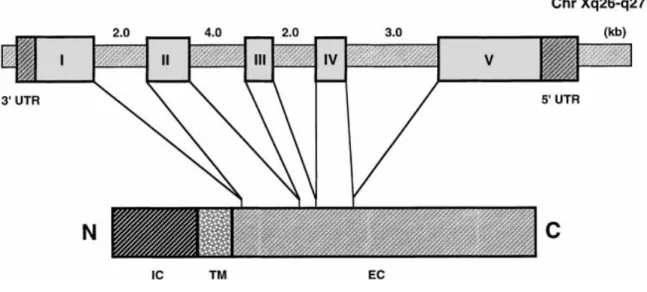
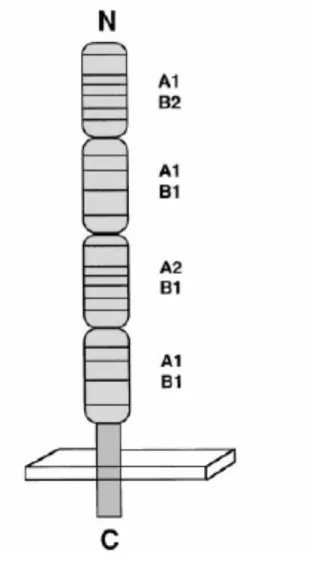

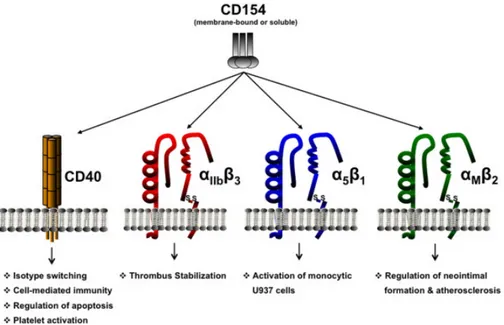
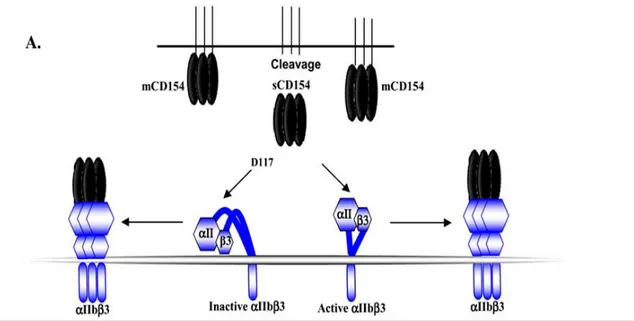
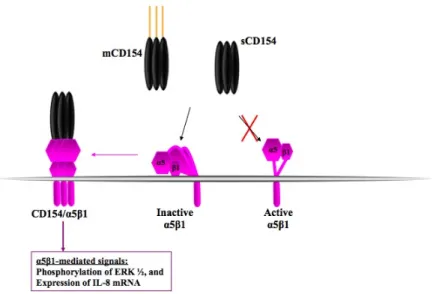
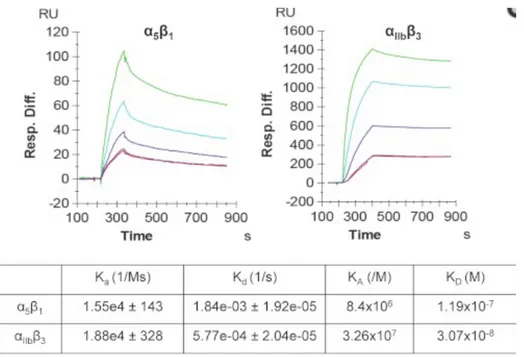

Documents relatifs
• Malgré le fait que 11% de la population est directement concerné par le syndrome du côlon irritable, l’efficacité des thérapies manuelles sur ce syndrome n’a été
L’objectif de cette revue de littérature était de déterminer au travers de la littérature scientifique, si le massage avec des huiles essentielles avait une efficacité sur la
[34,35] Une cohorte de 105 patients atteints de mucoviscidose chroniquement infectés par PA (âge 6 ans, et VEMS 40 %, patients non traités par an- tibiothérapie inhalée depuis
Severity and specifi city of neglect-like symptoms in patients with complex regional pain syndrome (CRPS) compared to chronic limb pain of other origins. Etiologic factors in the
La présente étude avait pour but d’évaluer l’efficacité et la tolérance de cette combinaison comme traitement ARV de première ligne des patients de Goma.. Matériel
Les patients bénéficiaient d’un traitement de CPAP versus CPAP placebo (CPAP sans pression positive) durant trois mois, suivi d’une période sans traitement d’un mois,
Pour aider les patients à mieux gérer leur pathologie, nous nous sommes interrogés sur le rôle du pharmacien dans l’optimisation de l’observance des
Extended adjuvant therapy with anastrozole among postmenopausal breast cancer patients: results from the randomized Austrian Breast and Colorectal Cancer Study Group Trial 6a..
