Implication des lipoprotéines dans le métabolisme normal et pathologique du tissu osseux
223
0
0
Texte intégral
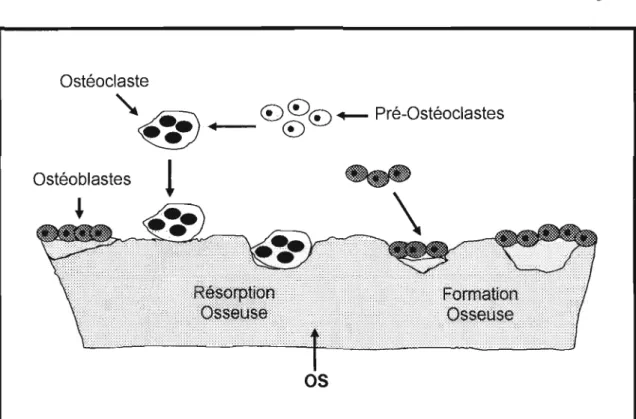
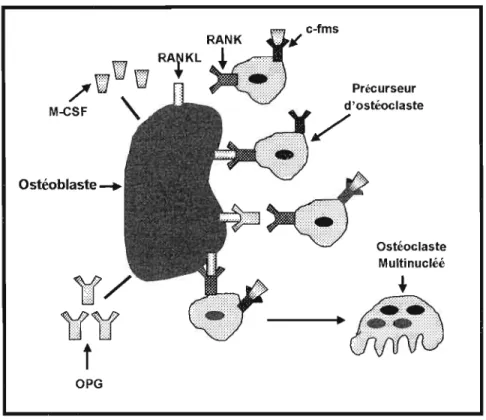
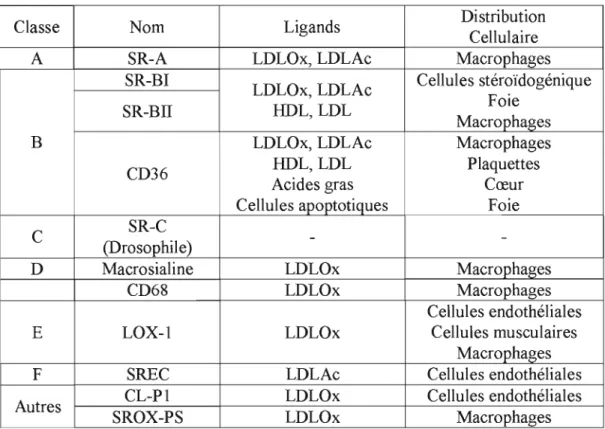
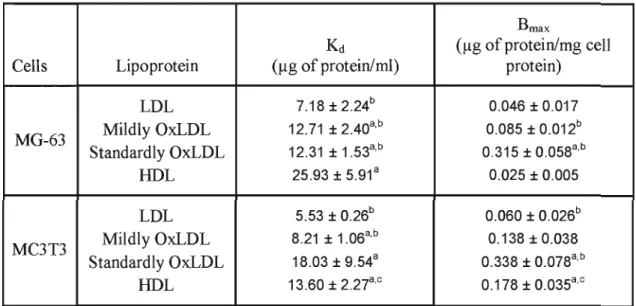
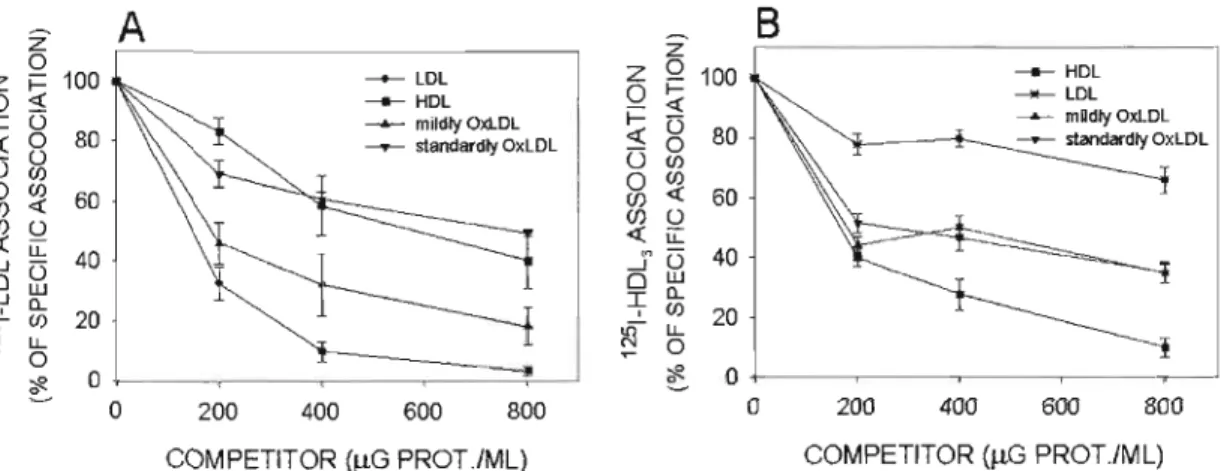
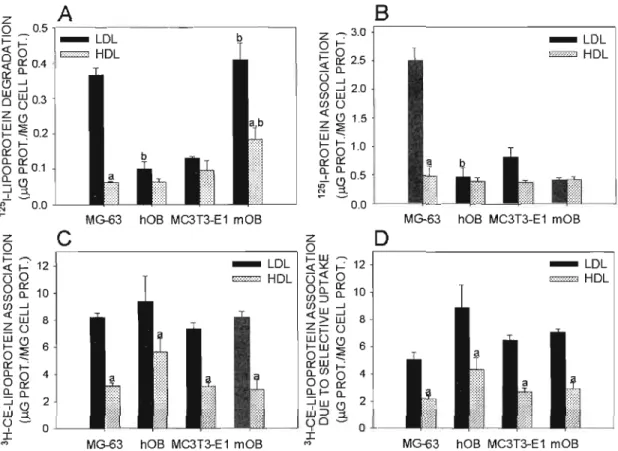
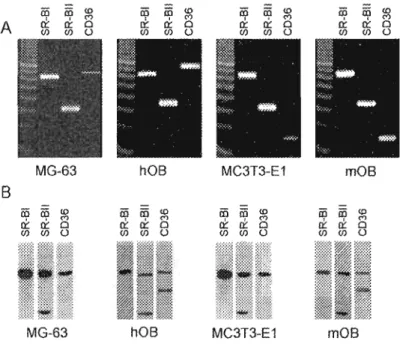
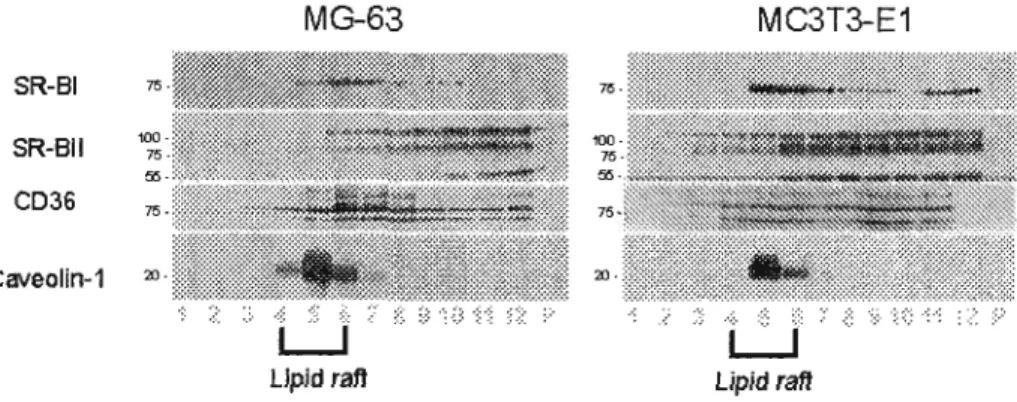
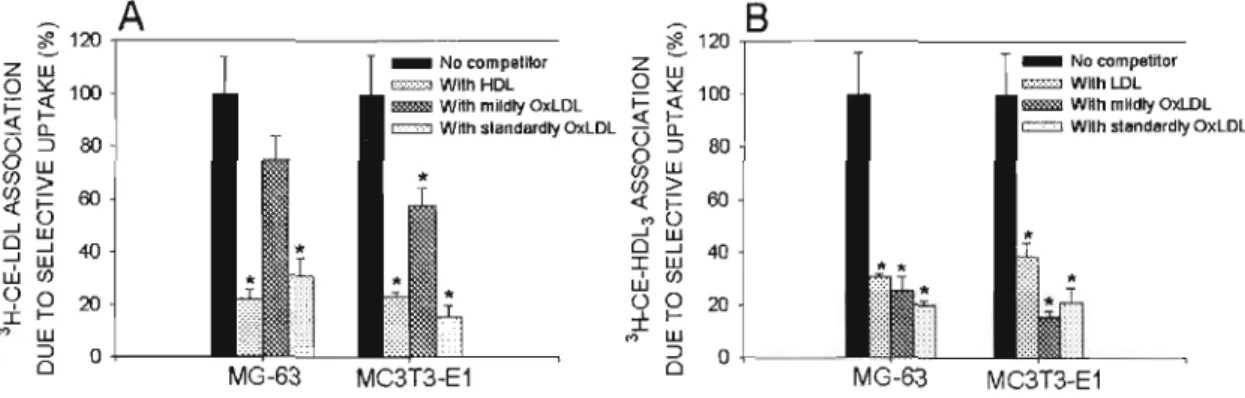
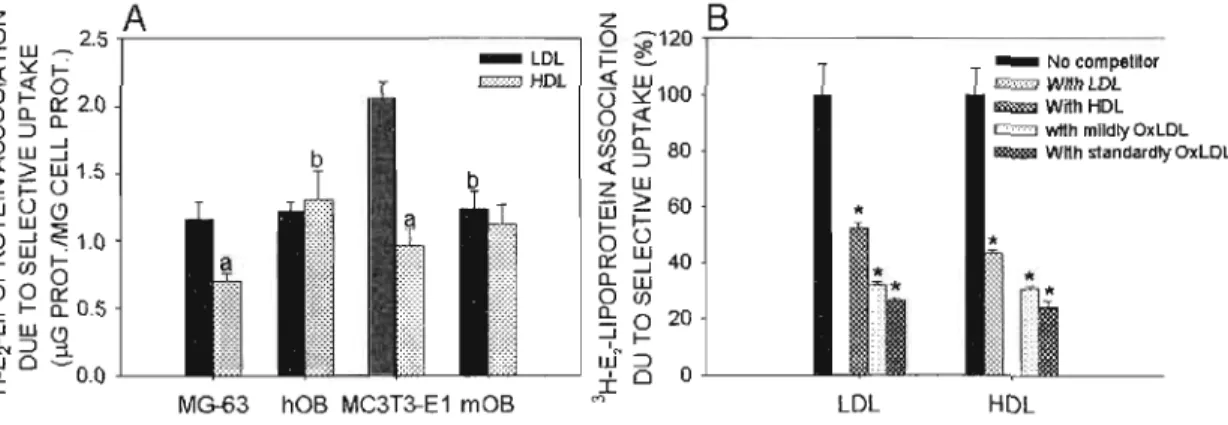
Figure
+7
Documents relatifs