Virulence Plasmids of Enterotoxigenic Escherichia Coli Isolates from Piglets
Texte intégral
Figure

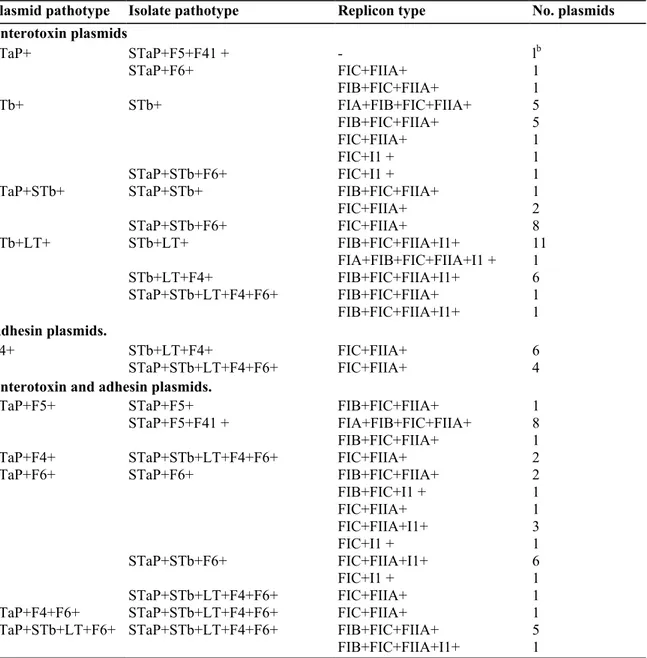
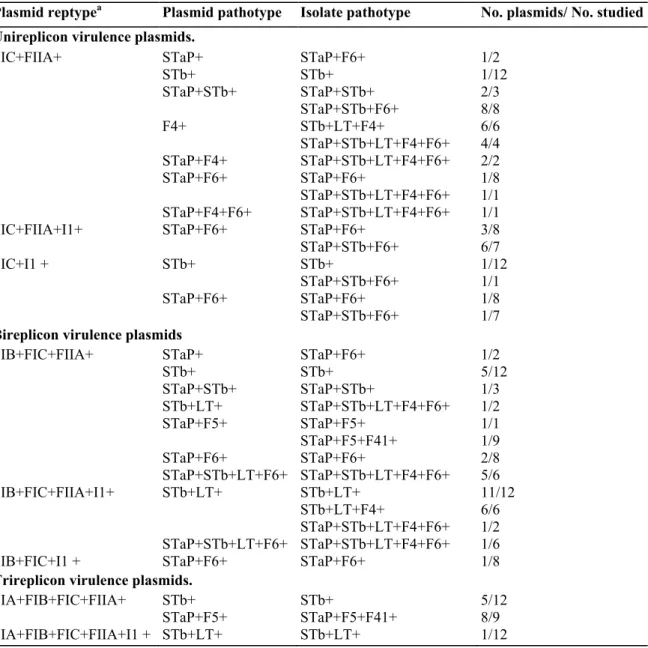
Documents relatifs
Inutile de chercher dans quel ensemble elle prend ses valeurs car la fonction x " x 2 est.. dérivable
Ainsi, la présence des indicateurs de contamination fécale (Coliformes totaux, Escherichia coli, Entérocoques et Anaérobie Sulfito-réducteurs) et des pathogènes (Staphylococcus
More than 80% of the farm A isolates belonged to the dominant MLVA cluster, except on two sampling events (three and seven days after farrowing) where < 40% of the animals carried
Strain typing of Mycoplasma cynos isolates from dogs with respiratory disease..
The sensitivity and specificity of the in vitro villous adhesion test for F4ab and F4ac 138. bacteria with the genotyping test for the mucin 4 as golden standard is represented in
Please cite this article as: Monecke, S., Kuhnert, P., Hotzel, H., Slickers, P., Ehricht, R., Microarray based study on virulence-associated genes and resistance determinants
Title: Dietary specific antibodies in spray-dried immune plasma prevent enterotoxigenic Escherichia coli F4 (ETEC) post weaning diarrhoea in piglets..
Please cite this article as: Gerjets, I., Traulsen, I., Reiners, K., Kemper, N., Comparison of virulence gene profiles of Escherichia coli isolates from sows with Coliform mastitis
