Impacts des dépositions atmosphériques de fer sur
les assemblages phytoplanctoniques et la
production de diméthylsulfure dans le Pacifique
Nord-Est contemporain et dans le contexte de
l’acidification des océans
Thèse
Josiane Mélançon
Doctorat en océanographie
Philosophiae doctor (Ph. D.)
Québec, Canada
© Josiane Mélançon, 2016
Impacts des dépositions atmosphériques de fer sur
les assemblages phytoplanctoniques et la
production de diméthylsulfure dans le Pacifique
Nord-Est contemporain et dans le contexte de
l’acidification des océans
Thèse
Josiane Mélançon
Sous la direction de :
Maurice Levasseur, directeur de recherche
Résumé
La croissance du phytoplancton est limitée par les faibles concentrations de fer (Fe) dans près de 40% de l’océan mondial. Le Pacifique subarctique Nord-Est représente une de ces zones limitées en fer et désignées High Nutrient - Low Chlorophyll (HNLC). Cet écosystème, dominé par des cellules de petite taille telles les prymnésiophytes, est caractérisé par de très faibles concentrations estivales de chlorophylle a et de fortes concentrations de macronutriments. Il a été maintes fois démontré que les ajouts de fer, sous différentes formes chimiques (habituellement FeSO4), dans les zones HNLC, stimulent la croissance et modifient la structure des communautés planctoniques en favorisant la croissance des cellules de grande taille, notamment les diatomées. Ces effets sur la communauté planctonique ont le potentiel d’influencer les grands mécanismes régulateurs du climat, tels la pompe biologique de carbone et la production de diméthylsulfure (DMS). Les poussières provenant des déserts du nord de la Chine sont reconnues depuis longtemps comme une source sporadique importante de fer pour le Pacifique Nord-Est. Malgré leur importance potentielle, l’influence directe exercée par ces poussières sur l’écosystème planctonique de cette zone HNLC n’a jamais été étudiée. Il s’agit d’une lacune importante puisque le fer associé aux poussières est peu soluble dans l’eau de mer, que la proportion biodisponible n’est pas connue et que les poussières peuvent avoir un effet inhibiteur chez le phytoplancton. Cette thèse propose donc, dans un premier temps, de mesurer pour la première fois l’effet de la fertilisation de la communauté planctonique du Pacifique Nord-Est par un gradient de concentrations de poussières désertiques naturelles. Cette première expérimentation a démontré que le fer contenu dans les poussières asiatiques est biodisponible et qu’une déposition équivalente à celles prenant place au printemps dans le Pacifique Nord-Est peut résulter en une stimulation significative de la prise de nutriments et de la croissance du phytoplancton. Mes travaux ont également montré que l’ajout de 0,5 mg L-1 de poussières peut résulter en la production d’autant de biomasse algale que l’ajout de FeSO4, l’espèce chimique utilisée lors des expériences d’enrichissement en fer à grande échelle. Cependant, les ajouts de FeSO4 favorisent davantage les cellules de petite taille que les ajouts de poussières, observation démontrant que le FeSO4 n’est pas un proxy parfait des poussières asiatiques. Dans un deuxième temps, je me suis intéressée à une source alternative de fer atmosphérique, les cendres volcaniques. Mon intérêt pour cette source de fer a été attisé par les observations d’une floraison spectaculaire dans le
Pacifique Nord-Est, ma région d’étude, associée à l’éruption de 2008 du volcan Kasatochi dans les îles Aléoutiennes. Forte de mon expérience sur les poussières, j’ai quantifié l’effet direct de ces cendres volcaniques sur la communauté planctonique du Pacifique Nord-Est. Mes résultats ont montré que le fer contenu dans les cendres volcaniques est également biodisponible pour le phytoplancton. Ils ont également montré que cette source de fer peut être aussi importante que les poussières désertiques dans la régulation de la croissance du phytoplancton dans cette partie de l’océan global à l’échelle millénaire. Dans un troisième temps, j’ai estimé comment l’acidification des océans modulera les réponses des communautés planctoniques aux dépositions naturelles de fer mises en évidence lors de mes expériences précédentes. Pour ce faire, j’ai effectué des enrichissements de poussière dans de l’eau de mer au pH actuel de 8.0 et dans l’eau de mer acidifiée à un pH de 7.8. Mes résultats ont montré une diminution du taux de croissance du phytoplancton dans le milieu acidifié mais pas de changement notable dans la structure de la communauté. Les ajouts de poussières et de cendres, de même que les variations de pH, n’ont pas eu d’effet significatif sur la production de DMS et de son précurseur le diméthylsulfoniopropionate (DMSP), probablement en raison de la courte durée (4 jours) des expériences. L’ensemble des résultats de cette thèse montre que le fer contenu dans diverses sources atmosphériques naturelles est biodisponible pour le phytoplancton du Pacifique Nord-Est et que des taux de déposition réalistes peuvent stimuler la croissance de manière notable dans les premiers jours suivant une tempête désertique ou une éruption volcanique. Finalement, les résultats de mes expériences à stresseurs multiples Fer/acidification suggèrent une certaine résistance des communautés phytoplanctoniques à la diminution du pH prédite d’ici la fin du siècle pour les eaux de surface des océans.
Abstract
Phytoplankton growth is limited by low concentrations of iron (Fe) in about 40% of the world’s oceans. The northeast subarctic Pacific Ocean embodies one of these Fe-limited regions that are termed High Nutrient - Low Chlorophyll (HNLC). Its ecosystem is dominated by small phytoplankton cells, such as Prymnesiophyceae, and is characterized by weak summer concentrations of chlorophyll a and high concentrations of macronutrients. It has repeatedly been shown that Fe additions in HNLC zones, using various artificial forms of Fe (commonly FeSO4), stimulate the growth and modify the structure of planktonic communities by favoring the blooming of large phytoplankton cells such as diatoms. The impacts brought about on the planktonic communities by these Fe additions have the potential of influencing large-scale climate regulating mechanisms, namely the biological carbon pump and the oceanic production of dimethylsulfide (DMS). Dust arising from northern Chinese deserts is well recognized as an important albeit sporadic source of Fe for the northeast Pacific Ocean. Despite their potential importance, the direct influence of these dust deposition events on the planktonic ecosystem of the northeast Pacific has never been studied before. This represents a serious shortcoming considering that, Fe associated with dust is scantily soluble in seawater, the bioavailable proportion of Fe in dust remains unknown, and dust may exert an inhibiting effect on phytoplankton. My thesis thus proposes, firstly, to conduct avant-garde measurements of the fertilization effects of a natural Asian dust gradient on plankton communities of the northeast Pacific. My first experiment shows that the Fe contained in Asian dust is bioavailable and that a deposition event, equivalent to naturally occurring spring depositions in the northeast Pacific, may result in a significant stimulation of nutrient uptake and growth by phytoplankton. My results also demonstrate that the addition of 0,5 mg L-1 of dust may induce as much biomass accumulation as what is observed during large scale fertilizing experiments utilizing FeSO4. However, small-celled phytoplankton assemblages are stimulated by Fe fertilizations, more so through FeSO4 than through dust additions demonstrating the flaws of this proxy in accurately representing Asian dust. In a second step, I focused my attention on an alternative source of atmospheric Fe, specifically volcanic ash. My interest for this subject was instigated by observations of a spectacular bloom occurring in my study region, the northeast Pacific, and associated to the 2008 eruption of the Kasatochi volcano located on one of the Aleutian Islands. I
quantified the direct effects of volcanic ash on the plankton community of the northeast Pacific. My results show that the Fe contained in these ashes is also bioavailable for phytoplankton and that this source of Fe may be as important as desert dust in regulating the growth of phytoplankton in this part of the global ocean on a millennial scale. Thirdly, I assessed the manner in which ocean acidification could modulate, on a short timescale, the responses of the plankton communities to natural Fe depositions highlighted in this thesis. In order to do so, I performed dust enrichments in seawater at the contemporary seawater pH of 8.0 and acidified to a pH of 7.8. My results show a reduction in phytoplankton growth rate in the acidified environment with no conspicuous changes to community structure. The additions of dust and ash, as well as the variations in pH, had little significant impacts on the production of DMS or its precursor dimethylsulfoniopropionate (DMSP). This lack of response could likely be attributable to the short timescale (4 days) of investigation carried out. As a whole, the results of this thesis show that Fe contained in various natural atmospheric sources is bioavailable to phytoplankton of the northeast Pacific Ocean. Furthermore, realistic deposition rates may distinctly stimulate phytoplankton growth in the first days following dust storms or volcanic eruptions. Finally, results from the multi-stressor experiments (Fe/acidification) I carried out suggest a demonstrable robustness of surface plankton communities to the pH reduction predicted before the turn of this century.
Table des matières
Résumé ... iii
Abstract ... v
Table des matières... vii
Liste des tableaux ... ix
Liste des figures ... x
Liste des principaux acronymes ... xii
Remerciements ... xiii
Avant-propos... xiv
Chapitre 1 – Introduction générale ... 1
1.1 Les zones High Nutrient - Low Chlorophyll et la limitation en fer ... 2
1.2 Les expériences de fertilisation à grande échelle ... 3
1.3 Sources de fer pour les océans ... 5
1.4 DMS et climat ... 6
1.5 L'acidification des océans ... 8
1.6 Le Pacifique nord-est : une zone limitée par le fer... 10
1.7 Problématique et objectifs ... 13
Chapitre 2 – Experimental assessment of the impact of Asian dust on phytoplankton dynamics and dimethylsulfide production in the northeast subarctic Pacific ... 15
2.1 Résumé ... 15
2.2 Abstract ... 15
2.3 Introduction ... 16
2.4 Material and Methods ... 18
2.4.1 Experimental settings ... 18
2.4.2 Treatments ... 19
2.4.3 Chemical and biological variables ... 20
2.4.4 Statistical Analysis ... 23
2.5 Results ... 23
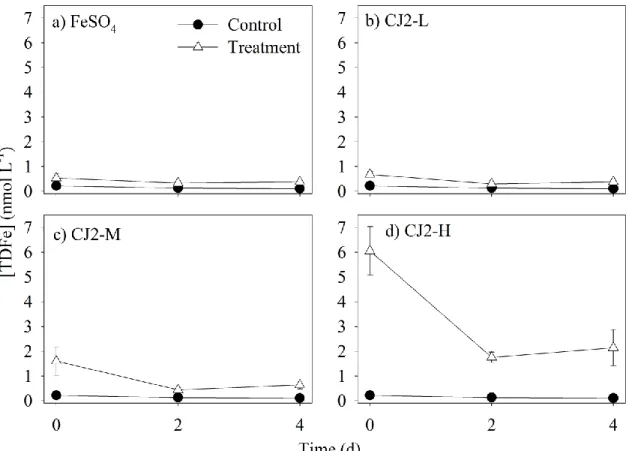
2.5.1 Iron concentrations ... 23
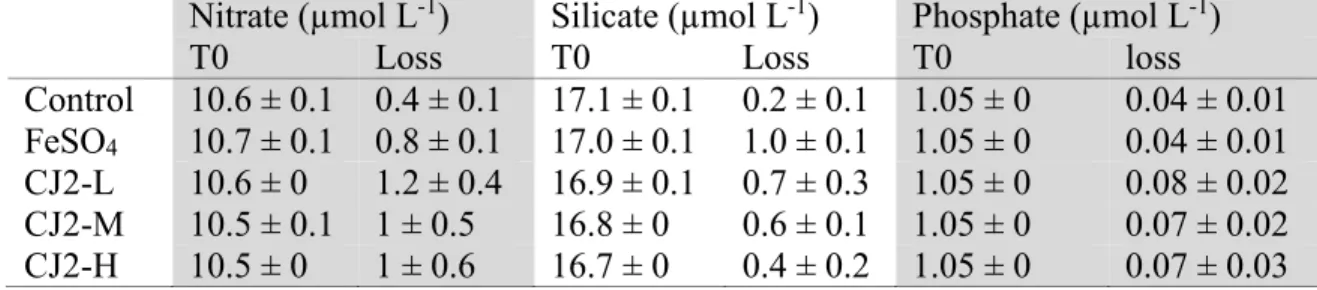
2.5.2 Nutrient concentrations and drawdown ... 24
2.5.3 Phytoplankton biomass ... 25
2.5.4 Phytoplankton assemblage ... 28
2.5.5 DMSP and DMS ... 30
2.6 Discussion ... 32
2.6.1 Initial conditions and experimental biases ... 32
2.6.2 Phytoplankton response to FeSO4 fertilization ... 32
2.6.3 Phytoplankton response to CJ-2 dust addition ... 35
2.6.4 Impact of dust deposition on DMSPt and DMS ... 38
2.7 Conclusions ... 39
2.8 Acknowledgements ... 41
Chapitre 3 - Early response of the northeast subarctic Pacific plankton assemblage to volcanic ash fertilization... 42
3.1 Résumé ... 42
3.2 Abstract ... 42
3.3 Introduction ... 42
3.4.1 Study area and experimental design ... 44
3.4.2 Ash origins, composition, and amendments ... 46
3.4.3 Biological variables ... 48
3.4.4 Statistical analyses ... 51
3.5 Results ... 51
3.5.1 Ash properties and Fe concentrations during the incubations ... 51
3.5.2 Phytoplankton ... 53
3.5.3 Primary productivity ... 57
3.6 Discussion ... 58
3.6.1 Considerations on Fe measurements ... 58
3.6.2 Response of phytoplankton to the addition of Kasatochi ash ... 60
3.6.3 Kasatochi ash vs. Chaitén ash ... 62
3.6.4 Response of phytoplankton to a concentration gradient of Chaitén ash ... 63
3.7 Acknowledgements ... 66
Chapitre 4 - Impact of ocean acidification on phytoplankton assemblage, growth and DMS production following Fe-Dust additions in the northeast Pacific high-nutrient, low-chlorophyll waters ... 67
4.1 Résumé ... 67
4.2 Abstract ... 67
4.3 Introduction ... 68
4.4 Material and Methods ... 71
4.4.1 Experimental setting and location ... 71
4.4.2 Treatments and acidification protocol ... 71
4.4.3 Fe and dust addition ... 72
4.4.4 Chemical and biological variables ... 73
4.4.5 Statistical Analysis ... 77
4.5 Results ... 77
4.5.1 State of the carbonate system, macronutrients and Fe concentrations ... 77
4.5.2 Plankton biomass ... 79
4.5.3 Taxonomy ... 82
4.5.4 Carbon and Fe uptake rates at T4 ... 83
4.5.5 DMSPt and DMS ... 84
4.6 Discussion ... 86
4.6.1 Considerations regarding the experimental protocol ... 86
4.6.2 Initial in situ conditions and impact of acidification ... 87
4.6.3 Dust fertilization in a high-CO2 northeast subarctic Pacific ... 89
4.6.4 Impact of acidification and dust deposition on DMSPt and DMS ... 92
4.7 Conclusion ... 94
4.8 Acknowledgments ... 94
Chapitre 5 - Conclusion ... 96
5.1 Contribution de la thèse aux connaissances ... 99
Liste des tableaux
Tableau 1.1 : Sources de fer dans l'océan global (adapté de Jickells et al. 2005)... 6 Table 2.1: Treatments applied and their natural representativeness. ... 20 Table 2.2 : Initial concentrations and loss after 4 days of nitrate, silicate and phosphate. ... 25 Table 2.3: Abundance of each taxonomic class at T0 and in one bag of each selected treatment and T4 (x102 cells L-1), and ratios of the abundances measured in the two treatments at T4. ... 28 Table 3.1: Physical and chemical properties of Chaitén and Kasatochi volcanic ash used in the fertilization experiments. ... 47 Table 4.1: Description of experimental conditions for each treatment. All treatments were conducted in triplicate. ... 72 Table 4.2: Values of DIC, alkalinity, pH and pCO2 in each treatment at T0 and T4. ... 78 Table 4.3: Biomarker pigment initial ratio matrix: Chl a ratios for eight algal groups. ... 82
Liste des figures
Figure 1.1: Concentrations moyennes annuelles de nitrate (µmol L-1) dans les eaux de surface de l’océan global (cellules de un degré). Le Pacifique Nord, le Pacifique Équatorial et l'Océan Austral présentent des concentrations élevées de nitrates. ... 3 Figure 1.2 : Prédictions de la diminution du pH des océans causée par l'augmentation de la concentration atmosphérique de dioxide de carbone. Tirée de Meehl et al. (2007) ... 8 Figure 1.3 : Variations temporelles des concentrations mensuelles moyennes de chlorophylle à la station PAPA dans le Pacifique Nord-Est (e, panneau de gauche) et répartition spatiale de la chlorophylle en août 2008 suite au passage du nuage de cendres du volcan Kasatochi (f, panneau de droite). Tiré de Hamme et al (2010). ... 12 Figure 2.1: Location of the sampling station (P26) along the Line P transect in the northeast subarctic Pacific Ocean. Graphic generated in Ocean Data View. ... 19 Figure 2.2 : Average concentrations of TDFe during the incubation in a) FeSO4 treatment, b) CJ2-L treatment, c) CJ2-M treatment and d) CJ2-H treatment. TDFe in the control incubation is repeated on each panel (black dots). Bars represent standard deviation. ... 24 Figure 2.3:Average concentrations of Chl a during the incubation and POC at T4 in a) FeSO4 treatment, b) CJ2-L treatment, c) CJ2-M treatment and d) CJ2-H treatment. Chl a and POC in the control incubation are repeated on each panel (black dots). Bars represent standard deviation. . 26 Figure 2.4: Average a) POC:Chl a and b) PON:Chl a ratios at T4 in all treatments and in the control incubation. Bars represent standard deviation. ... 27 Figure 2.5: Average concentrations of DMSPt during the incubation in a) FeSO4 treatment, b) CJ2-L treatment, c) CJ2-M treatment and d) CJ2-H treatment. DMSPt in the control incubation is repeated on each panel (black dots). Bars represent standard deviation. ... 29 Figure 2.6: Average DMSPt:Chl a ratio during the incubations. Bars represent standard deviation. ... 30 Figure 2.7: Average concentrations of DMS during the incubation in a) FeSO4 treatment, b) CJ2-L treatment, c) CJ2-M treatment and d) CJ2-H treatment. TDFe in the control incubation is repeated on each panel (black dots). Bars represent standard deviation. ... 31 Figure 2.8: Net rate of increase of Chl a during the exponential phase (days 2 to 4) plotted against (a) initial TDFe and (b) dust. ... 36 Figure 3.1: The northeast subarctic Pacific. Canadian monitoring Line P stretches from Sta. P1 to Ocean Sta. Papa (OSP). Experimentation was conducted at P20, a station within the HNLC region. The Kasatochi volcano location is identified by a triangle... 45 Figure 3.2: Average concentration of TDFe, DFe and Chl a in each treatment from day 0 to day 6. (A) Control, (B) Kasatochi Medium concentration (KasaM), (C) Chaitén Low concentration (ChaL), (D) Chaitén Medium concentration (ChaM), (E) Chaitén High concentration (ChaH), and (F) FeSO4. Error bars represent standard deviation. TDFe measurements for ChaH at T6 were over the analytical detection range (˃˃ 35 nmol L-1), their minimum value is represented by an open symbol. Dotted lines and open symbols in Chl a concentration panels illustrate the
replicate in which no growth has been observed in ChaM and ChaH treatments. Note the different scales on the y axis. ... 55 Figure 3.3: Average Chl a concentration (µg L-1) attributable to each of the measured groups of phytoplankton initially (T0; white bar) and for each of the treatments after 6 days of incubation (Control, KasaM, ChaL, ChaM, ChaH, Fe; grey bars). (A) Diatoms, (B) Prasinophytes, (C) Pelagophytes, (D) Haptophytes, (E) Cryptophytes, and (F) Cyanobacteria. Error bars indicate standard deviation. Letters above the bars indicate significant differences between Chl a concentrations of each group for each treatment. If two bars of a same panel share a letter, they are not statistically different. ... 56 Figure 3.4: 13C-uptake measurements. 5 to 7 hours parallel incubations enriched with 13C were run 24 h after the ash addition (T1) and after 6 days of incubation (T6). (A) Absolute uptake, (B) Pb: Absolute uptake normalized to Chl a. Filled symbols T1, open symbols T6... 58 Figure 3.5: (A, C) Absolute carbon uptake and (B, D) Chl a specific productivity (Pb) measured after 6 days of incubation as a function of (A, B) initial TDFe and (C, D) initial DFe. A hyperbolic Monod curve µ = µmax • [Fe] (KFe + [Fe])-1 was fitted through the data, where KFe corresponds to the half-saturation constant of TDFe or DFe. ... 64 Figure 3.6: Chl a increase rate (µg Chl a L-1d-1) during the exponential phase (between days 4 and 6 after enrichment) plotted against the concentration of Chaitén ash added in the replicates where growth has been observed. A Monod equation µ* = µmax • [ash] (Kash + [ash])-1 was fitted through the data. Correlation is significant (p = 0.0003). ... 65 Figure 4.1: Average concentration of DFe in each treatment during the incubations measured at T0, T2, and T4. (a) Control and Control+Acid. (b) Fe and Fe+Acid. (c) Dust and Dust+Acid. Error bars indicate standard deviations. ... 79 Figure 4.2: Average concentration of Chl a (left axis) during the incubations and POC at T4 (right axis) in each treatment. (a) Control and Control+Acid. (b) Fe and Fe+Acid. (c) Dust and Dust+Acid. Error bars indicate standard deviations. Dashed line indicates POC concentration at T0. ... 81 Figure 4.3: Average Chl a concentration (µg L-1) attributable to each of the measured groups of phytoplankton initially (T0: white bar) and for each of the treatments after 4 d of incubation (Control, Control+Acid, Fe, Fe+Acid, Dust, Dust+Acid; gray bars). (a) Diatoms. (b) Dinoflagellates. (c) Haptophytes. (d) Pelagophytes. (e) Chlorophytes. (f) Cyanobacteria. Error bars indicate standard deviations. ... 84 Figure 4.4: Average (a) Absolute C assimilation rates. (b) C assimilation rates normalized to Chl a concentration at T4 and (c) Fe uptake rates normalized to Chl a concentration at T4. Error bars indicate standard deviations. ... 85 Figure 4.5: DMSPt (a, b and c) and DMS (d, e and f) concentrations (nmol L-1) in the Control and Control+Acid treatments (a and d), the Fe and Fe+Acid treatments (b and e), and the Dust and Dust+Acid treatments (c and f). Error bars indicate standard deviations. ... 86 Figure 5.1: Courbe de type Monod mettant en relation les taux de d’augmentation de la chlorophylle a en phase exponentielle et la concentration initiale de fer labile (TDFe) dans les enrichissements en cendres du Chaitén et en poussières CJ2 pour les 3 expérimentations présentées dans chacun des chapitres. ... 100
Liste des principaux acronymes
ANOVA Analysis of variance
Chl a Chlorophyll a
DFe Dissolved iron
DIC Dissolved inorganic carbon
DMS Diméthylsulfide
DMSP Diméthylsulfoniopropionate
FeAXs Expérience d'enrichissement en fer à grande échelle
HNLC High nutrient - Low chlorophyll
HPLC High Performance Liquid Chromatography
NGCC/CCGS Navire de la Garde Côtière Canadienne/Canadian Coast Guard Ship
OA Ocean acidification
POC Particulate organic carbon
PON Particulate organic nitrogen
SERIES Subarctic Ecosystem Response to Iron Enrichment
TDFe Total dissolvable iron
Remerciements
Je tiens premièrement à remercier mon directeur de thèse Maurice Levasseur, qui m’a fait confiance dès le début. Il a su m’épauler aux moments où j’en avais le plus besoin. Mes conversations avec lui étaient toujours source d’inspiration et de motivation. Maurice, tu es un excellent directeur de recherche, et ton humanité n’a rien à envier à ta rigueur scientifique et perspicacité.
Je tiens aussi à remercier toute l’équipe qui m’a aidée dans cette aventure. D’abord Martine Lizotte, qui a été une pierre angulaire à toutes les étapes de mon projet de recherche, qui m’a initiée à pas mal tous les aspects pratiques de la recherche, de l’utilisation de l’équipement en laboratoire à la rédaction en passant par les missions en mer, et qui est d’un dévouement exemplaire. Mes co-directeurs Jean-Éric Tremblay et Michael Scarratt, ainsi que les membres de mon comité de suivi Connie Lovejoy et Philippe Tortell pour leurs conseils judicieux. Mes compagnes de missions Marjolaine Blais et Jessie Motard-Côté, et tous les membres du labo, présents et anciens, qui ont égayé mon cheminement de leur compagnie et ont su offrir aide, conseils et idées à des moments opportuns. Le groupe Takuvik et les biologistes qui dynamisent le département et en font une famille. Et bien sûr, les équipages du NGGC John P. Tully et les partenaires scientifiques de Pêche et Océans Canada qui ont été d’une aide inestimable.
Je tiens finalement à remercier ma famille qui m’a toujours encouragée dans mes études et mes nombreux projets, qui m’a poussée à aller toujours plus loin et à donner le meilleur de moi-même, qui m’a soutenue à tous les moments. Je remercie aussi Éric, qui m’a accompagnée au quotidien dans ce projet et a su me réconforter, m’encourager et me préparer des bons petits soupers…
Ce projet de recherche doctoral n’aurait pas été possible sans le soutien financier du FQRNT, du CRSNG, de Québec-Océan, de SOLAS et du département de biologie de l’Université Laval, je les en remercie.
Avant-propos
Cette thèse présente les résultats de mes travaux de recherche doctorale effectués sous la supervision du professeur Maurice Levasseur. J’ai effectué les expérimentations et collectes de données lors de trois campagnes océanographiques à bord du navire de la Garde Côtière Canadienne (NGCC) John P. Tully aux stations P20 et P26 du programme de monitorage de l’Institut des sciences océaniques, Pêches et Océans Canada, en juin 2009, août 2010 et juin 2011. J’ai été responsable de la planification, de la réalisation, de la collecte et de l’analyse des données de ces trois missions océanographiques réalisées avec l’appui de mon directeur de recherche, de collègues de l’Université Laval et de partenaires d’autres institutions.
Il s’agit d’une thèse par insertion d’articles, c’est-à-dire que les trois chapitres formant le corps de la thèse sont des articles publiés ou sur le point de l’être. En plus de ces articles, la thèse comprend une introduction générale et une conclusion générale rédigées en français. J’ai rédigé toutes les parties de cette thèse comme première auteure. Les co-auteurs de mes articles ont contribué à la rédaction en participant à la planification des expérimentations, en révisant et commentant les versions préliminaires de mes manuscrits, et en participant à l’interprétation des résultats.
Le premier article, intitulé « Experimental determination of the impact of Asian dust deposition on phytoplankton dynamics and dimethylsulfide production in the northeast subarctic Pacific », a pour co-auteurs Maurice Levasseur, Martine Lizotte, Michael Scarratt, Jean-Éric Tremblay, Marjolaine Blais, Gui-Peng Yang, Hui-Wang Gao, Shigenobu Takeda, Marie Robert, Michael Arychuk et Keith Johnson. Il sera soumis prochainement au Journal of Geophysical Research. Le deuxième article, intitulé « Early response of the northeast subarctic Pacific plankton assemblage to volcanic ash fertilization » a pour co-auteurs Maurice Levasseur, Martine Lizotte, Pierre Delmelle, Jay Cullen, Roberta C. Hamme, Angelica Peña, Kyle G. Simpson, Michael Scarratt, Jean-Éric Tremblay, Jie Zhou, Keith Johnson, Nes Sutherland, Michael Arychuk, Nina Nemcek et Marie Robert. Il a été publié dans Limnology and Oceanography en janvier 2014. Le troisième article, intitulé « Effects of dust additions on phytoplankton growth and DMS production in high CO2 northeast Pacific HNLC waters » a pour co-auteurs Maurice Levasseur,
Martine Lizotte, Michael Scarratt, Jean-Éric Tremblay, Philippe Tortell, Gui-Peng Yang, Hui-Wang Gao, David M. Semeniuk, Marie Robert, Michael Arychuk, Keith Johnson, Nes Sutherland, Marty Davelaar, Nina Nemcek et Angelica Peña. Il a été soumis le 10 juillet 2015 à la revue Biogeosciences. Il a été accepté le 16 juillet 2015 et a été publié en ligne le 6 août 2015. Il n’y a pas de difference entre les articles insérés et ceux publiés. Au terme de leur processus de révision, les versions publiées des chapitres 1 et 3 pourront différer légèrement de celles présentées dans la thèse, mais ces modifications ne devraient pas affecter la nature et l’interprétation générale des résultats.
Chapitre 1 – Introduction générale
Le phytoplancton est constitué d’algues unicellulaires vivant dans tous les plans d’eau douce ou salée. Il exerce une influence majeure pour plusieurs systèmes et cycles planétaires. De par son activité photosynthétique, le phytoplancton participe au cycle du carbone et joue le rôle essentiel de base de la chaîne trophique océanique. On estime à 43.5 Pg C an-1 la quantité de carbone fixée par le phytoplancton marin, soit environ la moitié de la production primaire mondiale (Behrenfeld and Falkowski, 1997; Field et al., 1998). C’est de cette production que dépendent tous les organismes marins pour se nourrir, du zooplancton aux mammifères en passant par les poissons et les oiseaux marins. La photosynthèse produit aussi de l’oxygène (O2), une molécule essentielle à la vie de la grande majorité des organismes vivants. On estime que le phytoplancton produit ainsi 50% de l’oxygène disponible sur terre. De plus, une partie du carbone fixé par le phytoplancton lors de la photosynthèse est exportée vers les eaux profondes et ultimement les sédiments marins où elle demeurera confinée pour des durées pouvant atteindre plusieurs milliers d’années. Le phytoplancton participe ainsi à la régulation du climat planétaire par la réduction de la concentration de gaz carbonique (CO2), un gaz à effet de serre; c’est un mécanisme appelé la pompe biologique du carbone (Falkowski et al., 1998; Sabine et al., 2004). De plus, le phytoplancton produit le diméthylsulfoniopropionate (DMSP), un osmolyte dont une part est transformée en diméthylsulfure (DMS), un composé soufré contribuant à l’équilibre climatique de la Terre. Lorsqu’émis dans l’atmosphère, le DMS est photooxydé en acides sulfurique et en acides méthane sulfonique, composés contribuant à la formation de noyaux de condensation de nuages, augmentant ainsi l’albedo de la planète et réduisant sa température (Charlson et al., 1987; Mahajan et al., 2015).
Au vu des nombreux rôles et services écosystémiques du phytoplancton et de son influence dans les cycles majeurs assurant une stabilité au système planétaire, il apparaît important de bien connaître son écologie et les facteurs modulant sa croissance. Cette thèse propose d’étudier la façon dont les dépositions atmosphériques de fer peuvent influencer la croissance et la spéciation du phytoplancton dans une zone océanique où les faibles concentrations de fer limitent sa croissance. Pour ce faire, des expérimentations de
fertilisation de communautés phytoplanctoniques ont été effectuées pour la première fois à l’aide de deux sources naturelles de fer, soit les poussières de désert et les cendres volcaniques. De plus, y sont présentés les résultats d’une étude multifactorielle montrant comment l’acidification des océans, une conséquence de l’augmentation du gaz carbonique dans l’atmosphère, pourra moduler l’impact des dépositions atmosphériques de fer sur les écosystèmes planctoniques de ces larges zones océaniques limitées en fer.
1.1 Les zones High Nutrient - Low Chlorophyll et la limitation en fer
La communauté scientifique a longtemps été intriguée par l’existence de grandes zones océaniques où la croissance phytoplanctonique demeure faible à l’année malgré une surabondance de nutriments tels les phosphates, les nitrates et les silicates (Figure 1.1). Ces zones, désignées High Nutrient – Low Chlorophyll (HNLC), couvrent près de 40% de l’océan global. On les retrouve dans le Pacifique nord-est subarctique, le Pacifique équatorial et l'Océan Austral. D'abord émise en 1931 par Gran (de Baar et al., 2005), l'hypothèse de la limitation en fer de ces zones ne fut démontrée qu'en 1988 par Martin et Fitzwater (1988). Ces auteurs effectuèrent des expériences d’enrichissement en fer dissous avec de l’eau prélevée à la station PAPA, au cœur du Pacifique nord-est subarctique. Leurs résultats furent probants: après 3 jours d'incubation, la quantité de chlorophylle a dans les bouteilles enrichies en fer augmentait, tandis qu'elle restait stable dans les contrôles. L’assimilation de nitrate, autre indicateur de croissance phytoplanctonique, supportait aussi leur hypothèse: les nitrates étaient presqu'entièrement consommés dans les enrichissements en fer tandis que seulement 25% étaient consommés dans les bouteilles contrôles. Forts de ces premiers résultats, ils posèrent l'hypothèse qu'une carence en fer pouvait expliquer la faible croissance phytoplanctonique et l’utilisation des éléments nutritifs caractérisant les autres zones HNLC. De plus, par extrapolation des variations synchrones mais de direction opposée observées au cours des périodes glaciales/interglaciales entre les concentrations de poussière et les teneurs en CO2 des bulles d’air trappées dans les carottes prélevées en Antarctique, les auteurs avancèrent que la déposition de fer dans les océans pourrait jouer un rôle majeur dans le contrôle du climat en favorisant la photosynthèse et l’absorption du CO2 atmosphérique. Cette découverte, une des plus importantes en océanographie, attira
l'attention de nombreux scientifiques et entraîna une multiplication rapide des travaux de recherche sur la biogéochimie du fer dans les océans.
Figure 1.1: Concentrations moyennes annuelles de nitrate (µmol L-1) dans les eaux de surface de l’océan global (cellules de un degré). Le Pacifique Nord, le Pacifique Équatorial et l'Océan Austral présentent des concentrations élevées de nitrates.
1.2 Les expériences de fertilisation à grande échelle
Afin de mieux comprendre l’influence des dépositions de fer sur les zones HNLC, il a rapidement été convenu par la communauté scientifique qu’il était nécessaire de mener des expériences à plus grande échelle. De 1993 à aujourd'hui, 13 expérimentations de fertilisation en fer à moyenne échelle (FeAXs) ont eu lieu dans les zones HNLC (Boyd et al., 2007; de Baar et al., 2005; Strong et al., 2009). Ces expérimentations sont résumées par Boyd et collaborateurs (2007). Malgré quelques différences au niveau de la saison, de la superficie fertilisée ou du nombre d'additions de fer, elles ont toutes suivi le même modèle général: une grande quantité de sulfate de fer (concentration finale de 2-3 nmol L-1) préalablement dissous dans une eau acidifiée, est répandue sur une superficie de plusieurs kilomètres carrés. L'aire ainsi fertilisée est ensuite échantillonnée selon le principe
Lagrangien pendant des périodes de quelques jours à plusieurs semaines selon les contraintes logistiques.
Dans toutes les FeAXs, une stimulation de la croissance phytoplanctonique a été observée suite à l'ajout de fer. Cette stimulation s’est traduite par des augmentations de l'assimilation de nitrates et de phosphates, de la performance photosynthétique des microalgues et des concentrations de chlorophylle a (voir Boyd et al., 2007; de Baar et al., 2005 pour des revues des FeAXs). La croissance phytoplanctonique induite par le fer est souvent suivie d'une croissance des populations zooplanctoniques. Cependant, ce ne sont pas tous les groupes phytoplanctoniques qui bénéficient de l'apport de fer: dans la plupart des cas, une croissance immédiate de tous les groupes est mesurée, mais les diatomées dominent rapidement les floraisons après quelques jours (Boyd et al., 2007; de Baar et al., 2005; Marchetti et al., 2006b). La prédominance des diatomées résulterait en partie du taux de croissance plus lent de leur prédateur (macrozooplankton) alors que le taux de croissance rapide du microzooplankton lui permettrait d’exercer un contrôle plus efficace sur la biomasse des petites cellules (Boyd et al., 1996).
Si ces études ont permis de confirmer l'importance du fer dans la régulation de la croissance planctonique dans les zones HNLC, elles mirent également en évidence une faible efficacité de séquestration à long terme du carbone nouvellement fixé par unité de fer ajouté. En effet, très peu d'exportation de carbone organique particulaire (POC) a été mesurée dans l'ensemble de ces expériences (de Baar et al., 2005), vraisemblablement à cause du broutage, de la respiration et de la reminéralisation du carbone organique avant sa séquestration sous la thermocline. De Baar et ses collaborateurs notent aussi que même en utilisant de grandes quantités de fer, donc en soulageant cette carence à moyen terme, d'autres facteurs viennent rapidement limiter la croissance: la lumière (reliée au cycle saisonnier et à la profondeur de couche de mélange) et l'épuisement des macronutriments, tels les silicates. Les FeAXs ont été malgré tout très importantes pour l'avancement des connaissances sur l'écologie du plancton des zones NHLC.
1.3 Sources de fer pour les océans
Les flux globaux de fer dans les océans ont été résumés par Jickells et collaborateurs (2005) et sont présentés dans le tableau 1.1. Les fleuves et les sédiments glaciaires représentent des sources importantes de fer pour l’océan, cependant, le fer provenant de ces sources reste principalement confiné aux zones côtières et n’est donc pas disponible pour les organismes pélagiques, particulièrement dans les zones HNLC, éloignées des côtes (Poulton and Raiswell, 2002). Compte-tenu de la profondeur et de la stratification de l'Océan, le fer d'origine hydrothermique (sources thermales sous-marines) et authigénique (provenant des sédiments marins) influence peu les couches de surface où la croissance phytoplanctonique prend place (Jickells et al., 2005). Par conséquent, les dépositions atmosphériques, sèches et humides, représentent la principale source de fer en haute mer.
Le fer atmosphérique provient principalement des poussières des déserts, des activités anthropiques et, dans une moindre mesure, du cosmos (Jickells et al., 2005). Encore peu étudiées, les dépositions de cendres d'origine volcanique pourraient également représenter une source non négligeable de fer pour les océans (Olgun et al., 2011). De récentes études ont mis en évidence un relargage rapide (de l’ordre des minutes) de fer suivant le contact de la cendre volcanique avec l’eau de mer (Duggen et al., 2007; Jones and Gislason, 2008; Olgun et al., 2011). Des calculs effectués par Olgun et al. 2011 suggèrent que depuis le dernier maximum glaciaire et sur une échelle temporelle approchant le millénaire, le flux de cendres volcaniques dans l’Océan Pacifique se compare au flux atmosphérique de poussières des continents. Ainsi, les dépositions de cendres volcaniques et de poussières pourraient avoir un effet comparable sur le cycle du fer et le phytoplancton dans les régions HNLC. Cependant, la réponse du phytoplancton à l’ajout de cendres volcaniques a été très peu étudiée. Les deux études réalisées jusqu’à maintenant ont montré des réponses variées, allant de la stimulation à l’inhibition de la croissance du phytoplancton (Duggen et al., 2007; Hoffmann et al., 2012).
Tableau 1.1 : Sources de fer dans l'océan global (adapté de Jickells et al. 2005).
Source Flux (Tg de fer an-1)
Fluvial (total particulaire) 625 à 962
Fluvial (dissous) 1.5 Sédiments glaciaires 34 à 211 Atmosphérique 16 Érosion côtière 8 Hydrothermique 14 Authigénique 5 1.4 DMS et climat
Le DMS est un gaz d'origine naturelle abondant et ubiquitaire dans les océans. Il représente environ 50% des émissions naturelles de soufre dans l'atmosphère (Andreae, 1990), soit 17.6-34.4 Tg de soufre par année (Lana et al., 2011) et est présent dans les eaux de surface de l'ensemble des océans à une concentration moyenne de 3 nmol L-1 (Cooper and Matrai, 1989). La ventilation du DMS vers l'atmosphère participe à la régulation du climat de la planète. Dans l’atmosphère, le DMS est photooxidé en acide sulfurique et méthane sulfonique, des précurseurs d’aérosols qui contribuent à la formation de noyaux de condensation de nuages. Les aérosols et les nuages augmentent l'albédo de la planète en reflétant et dispersant les rayons solaires, contribuant ainsi à diminuer la quantité de rayons atteignant sa surface et de ce fait, diminuant sa température (Charlson et al., 1987; Mahajan et al., 2015).
Le DMS provient du clivage de son précurseur, le diméthylsulfoniopropionate (DMSP), par des DMSP-lyases algales et bactériennes. La production de DMS est un processus complexe mettant en scène les algues, le zooplancton, les bactéries et les virus planctoniques (voir Kiene et al., 2000 pour une schématisation du cycle du DMS et DMSP). La production nette de DMS est le résultat de l’équilibre dynamique entre les processus de production (clivage du DMSP par les lyases algales et bactériennes) et les
puits, principalement la consommation bactérienne (Zubkov et al., 2004; Zubkov et al., 2002). Le DMSP est produit par de nombreuses espèces algales à des concentrations intracellulaires variables selon l'espèce, l'état de santé des cellules et les conditions environnementales (Keller et al., 1989; Yoch, 2002). Les groupes phytoplanctoniques présentant les concentrations cytosoliques de DMSP les plus élevées sont les petits dinoflagellés, les prymnésiophytes et les chrysophytes. À l’autre bout du spectre se situent les diatomées, faibles producteurs de DMSP. D'abord identifié comme osmolyte ou cryoprotecteur (Kirst et al., 1991), le DMSP est maintenant aussi connu comme un antioxydant important pour le phytoplancton (Sunda et al., 2002). En effet, la production de DMSP et son clivage sont stimulés par des facteurs de stress tels les rayons UV, les limitations en fer ou en CO2 et une forte concentration en Cu2+ ou en H2O2.
Plusieurs études se sont intéressées à l’impact des limitations en fer sur la production océanique de DMS. Ces expériences de fertilisation en fer ont généré des réponses très variables (Boyd et al., 2007). En augmentant par addition de fer la productivité et la biomasse phytoplanctonique, on peut s'attendre à ce que la quantité absolue de DMSP augmente, amplifiant ainsi le potentiel de formation du DMS. Des augmentations de la concentration de DMS suite à l'addition de fer ont d’ailleurs souvent été observées, par exemple lors des expériences SOIREE et EisenEx dans l'Océan Austral (Turner et al., 2004) et lors des études IronEx I et II dans le Pacifique équatorial (Coale et al., 1996; Martin et al., 1994). Cependant, lors de l’expérience Subarctic Ecosystem Response to Iron
Enrichment (SERIES) réalisée dans le Pacifique subarctique nord-est, la concentration de
DMS a diminué suite à l’ajout de fer (Boyd et al., 2005; Levasseur et al., 2006) et ce malgré une augmentation des concentrations de DMSP. Cette diminution des concentrations de DMS a été attribuée à une stimulation de la consommation bactérienne de DMS qui a éventuellement dépassé le taux de production (Merzouk et al. 2006). Une fertilisation en fer pourrait également diminuer la production de DMS pour des raisons physiologiques. Sunda et collaborateurs (2002) ont montré qu'une limitation en fer, agissant comme un stress oxydant, induisait une plus grande production de DMSP et un plus fort taux de clivage en DMS. En soulageant cette limitation en fer, on pourrait donc s'attendre à ce que la production de DMSP et le clivage algal en DMS diminue. Il est donc difficile à ce jour de
prédire avec confiance quel sera l’impact des fertilisations en fer sur la production du DMS dans les zones HNLC. De plus, les quelques études mentionnées précédemment ont toutes utilisé du fer sous forme dissoute, ce qui rend l’extrapolation des résultats aux dépositions naturelles encore plus risquée.
1.5 L'acidification des océans
Figure 1.2 : Prédictions de la diminution du pH des océans causée par l'augmentation de la concentration atmosphérique de dioxide de carbone. Tirée de Meehl et al. (2007)
Souvent qualifiée de ''l'autre problème du CO2'', l'acidification des océans préoccupe de plus en plus la communauté scientifique internationale. Cette acidification est causée par l'absorption par les océans de l'excès de gaz carbonique émis par les activités anthropiques; jusqu'à maintenant, l'océan aurait absorbé 30% de ces émissions. Cette absorption par l'océan permet de ralentir le réchauffement global causé par les émissions de gaz à effet de serre; par contre, en se dissolvant dans l'eau, le gaz carbonique contribue à la formation
d'acide carbonique et par conséquent, à la réduction du pH de l'océan. Depuis le début de l'ère industrielle, le pH aurait déjà diminué de 0.1 unité, soit une augmentation de 30% de la quantité d'ions H+. Selon le scenario IS92a du GIEC (Meehl et al., 2007), le pH de l'océan pourrait diminuer de 0.3 à 0.4 unité supplémentaire d'ici 2100 (Figure 1.2). Cette acidification des océans pourrait avoir des conséquences variées sur les organismes y vivant et possiblement sur la manière dont les océans influencent les grands cycles biogéochimiques (Reid et al., 2009). Parmi les organismes qui risquent d'être le plus grandement affectés par l'acidification, on trouve ceux formant un squelette de carbonate de calcium (Denman et al., 2007). En effet, l'acidification de l'océan a pour effet net une réduction de la concentration d'ions carbonate (CO32-), qui réagissent avec les protons libres pour former du bicarbonate (HCO3-). Dans ces conditions, on peut s'attendre à ce que la calcification soit inhibée ou ralentie, ce que montrent quelques études (Delille et al., 2005; Riebesell et al., 2000). Cependant, la croissance des organismes calcifiants, ainsi que leur efficacité photosynthétique et leur abondance ne semblent pas affectées (Kroeker et al., 2013 et les références incluses). Par contre, d'autres études montrent aussi une augmentation de la calcification avec l'augmentation de la pression partielle de CO2 (Feng et al., 2008; Iglesias-Rodriguez et al., 2008; Wood et al., 2008).
Une augmentation de la pression partielle de CO2 peut aussi avoir un effet sur la structure de la communauté phytoplanctonique. Une étude combinant des mesures in situ et des incubations en bouteilles a démontré qu'une forte concentration de CO2 pouvait induire une plus grande productivité primaire et un changement de la structure de la communauté phytoplanctonique en faveur des grandes diatomées dans l'Océan Austral (Tortell et al., 2008). Cette étude a mis en évidence que, tout comme le fer, le CO2 peut avoir un effet sur la pompe biologique de carbone et par conséquent, sur la régulation du climat. Une étude similaire dans le Pacifique équatorial a aussi montré une prédominance de diatomées à de fortes concentrations de CO2, mais cette fois, sans effet notable sur la productivité primaire (Tortell et al., 2002). L'acidification peut également modifier la physiologie de certains organismes. De tels changements de la physiologie et de la structure des communautés phytoplanctoniques pourraient amener une modification de la production nette de DMSP et de DMS et avoir un effet sur la production d’aérosol soufrés et sur le cycle global du
soufre. Les études réalisées à ce jour sur l’effet d’une réduction de pH sur la production de DMS ont révélé soit des diminutions, soit des augmentations de la production de DMS dans des eaux acidifiées (Archer et al., 2013; Arnold et al., 2013; Avgoustidi et al., 2012; Hopkins et al., 2010). Ces résultats divergents illustrent la nécessité de poursuivre les études afin de mieux cerner les mécanismes responsables de la variabilité des réponses.
L’acidification des océans pourrait modifier la réponse de la communauté planctonique aux additions de fer en rendant le fer plus ou moins biodisponible. Les quelques études sur la biodisponibilité du fer en milieu acidifié ont généré des résultats contrastés. Breitbarth et al. (2010b) ont observé une augmentation de la concentration et de la demi-vie de la forme biodisponible Fe(II) en milieu acidifié, ce qui devrait se traduire par une meilleure utilisation du fer par les organismes marins. Cependant, Shi et al. (2010) ont montré que la complexation du Fe(III) par des ligands organiques contenant des groupements fonctionnels non-protonés acides est plus forte à faible pH, ce qui résulterait en une diminution du fer biodisponible. D’autre part, le taux de prise de fer par certains ligands organiques pourrait diminuer à faibles pH, la réduction enzymatique du fer à la surface des cellules pouvant relâcher des protons (Shi et al., 2010). Il est donc difficile de prévoir quel sera l’effet de l’acidification des eaux de surface des océans et plus particulièrement des zones HNLC sur la biodisponiblité du fer. De plus, aucune étude n’a été effectuée jusqu’à maintenant sur l’effet d’une réduction de pH sur la biodisponibilité du fer lors de dépositions atmosphériques de poussière.
1.6 Le Pacifique nord-est : une zone limitée par le fer
Le gyre de l’Alaska, situé dans le Pacifique nord-est, est une des trois grandes zones HNLC de l’océan mondial. C’est d’ailleurs dans ce gyre que Martin et Fitzwater (1988) ont démontré pour la première fois la limitation par le fer des zones HNLC. Les caractéristiques physiques, chimiques et biologiques du Pacifique nord-est sont relativement bien connues. La région est l’objet d’une ligne de monitorage qui s’étend de la zone côtière du Canada jusqu’à la Station PAPA, située à 50oN, 145oW, en plein cœur de la zone HNLC. Cette ligne de monitorage est visitée plusieurs fois par année depuis la fin des
années ’50 (Tabata and Weichselbaumer, 1992). Aux stations les plus au large, les concentrations moyennes de fer dissous dans les eaux de surface sont généralement faibles (0.01–0.06 nM), les concentrations en macronutriments (nitrate, phosphate, silicate) élevées et la production primaire faible et régulière toute l’année (0.3 g C m−2 d−1 en hiver et 0.85 g C m−2 d−1 au printemps/été) (Boyd and Harrison, 1999; Nishioka et al., 2001). La communauté phytoplanctonique y est dominée par de petites cellules, principalement des prymnésiophytes dont le coccolithophore Emiliania huxleyi. Le gyre de l’Alaska présente des concentrations estivales particulièrement élevées de DMS, jusqu’à 10 fois supérieures à la moyenne océanique de 3 nmol L-1 (Lana et al., 2011). Il s’agissait donc d’une zone HNLC idéale pour mener les recherches de ma thèse.
Les poussières asiatiques ont longtemps été considérées comme la principale source de fer dans le Pacifique subarctique nord-est (Duce and Tindale, 1991; Moore and Braucher, 2008; Moore et al., 2002). De récentes études ont cependant démontré que d'autres sources importantes de fer existaient dans cette région. Cullen et collaborateurs (2009) ont montré que le transport horizontal par les courants de marées en provenance du plateau continental de la Colombie-Britannique pouvait transporter une quantité de fer dans les eaux HNLC du gyre de l'Alaska du même ordre de grandeur que les dépositions atmosphériques. D’autre part, on sait maintenant que les tourbillons d’eaux côtières riches en fer pouvaient contribuer de 5 à 50% de l’apport en fer dans la partie sud du gyre de l’Alaska (Johnson et al., 2005; Ladd et al., 2005; Miller et al., 2005). L'apport en fer des sources hydrothermales a aussi été revu et de récentes études montrent qu'une partie resterait soluble et participerait au réservoir de fer disponible dans le Pacifique (Bennett et al., 2008; Toner et al., 2009).
Les cendres volcaniques pourraient aussi constituer une source non négligeable de fer pour cette zone HNLC, comme l’ont suggéré des observations suivant une éruption volcanique récente. En août 2008, l’éruption du volcan Kasatochi dans les îles aléoutiennes a déclenché une floraison massive dans le gyre de l’Alaska (Figure 1.3) (Hamme et al., 2010; Langmann et al., 2010a). Des indicateurs de croissance ont été détectés tant par satellite que par des instruments autonomes in situ quelques jours après le passage du nuage de cendres au-dessus du gyre de l’Alaska (Langmann et al., 2010b). Dix jours après la déposition des
cendres et le début de la floraison, les mesures effectuées dans le cadre programme de monitorage Line P rapportaient des concentrations inhabituellement élevées de chlorophylle a reliées à une croissance des diatomées (Hamme et al., 2010). Un lien, bien que controversé, entre cette floraison spectaculaire et un taux de retour particulièrement élevé des saumons sockeye dans la rivière Fraser en 2010 a d’ailleurs été proposé (McKinnell, 2013; Parsons and Whitney, 2012). Plusieurs facteurs ont contribué à faire de cette éruption un évènement écologique important et bien documenté : sa coïncidence avec un système de basse pression qui a transporté les cendres vers le gyre de l’Alaska, le fait que l’éruption se soit produite pendant un mois d’été alors que la production primaire n’est pas limitée par la lumière, et finalement la présence de mouillages et d’un navire océanographique dans la zone touchée pendant et après la déposition. Malgré la quantité impressionnante de données récoltées, plusieurs questions demeurent. Parmi celles-ci, on ne connait pas exactement la quantité de cendre requise pour initier la réponse observée au niveau du phytoplancton, ni quelles espèces sont plus particulièrement affectées (positivement ou négativement), et finalement comment ces enrichissements affectent la dynamique du DMS.
Figure 1.3 : Variations temporelles des concentrations mensuelles moyennes de chlorophylle à la station PAPA dans le Pacifique Nord-Est (e, panneau de gauche) et répartition spatiale de la chlorophylle en août 2008 suite au passage du nuage de cendres du volcan Kasatochi (f, panneau de droite). Tiré de Hamme et al (2010).
1.7 Problématique et objectifs
Les expérimentations ayant permis d'étudier l'effet du fer sur les écosystèmes en régions HNLC ont utilisé des sources de fer différant des sources naturelles de par leur nature et leur forte concentration (Boyd et al., 2007). Or, le fer de sources naturelles peut présenter des propriétés différentes au niveau de sa solubilité et de sa biodisponibilité en plus d'être accompagné d'autres éléments organiques ou inorganiques pouvant aussi avoir un effet sur la croissance du phytoplancton. L’absence d’expériences menées avec des poussières et des cendres naturelles limite grandement notre compréhension de cet écosystème important.
De plus, il est important d’explorer comment l’inévitable acidification des océans, et dans le cas qui nous intéresse ici celle des zones HNLC, affectera la biodisponibilité du fer issu des poussières et les communautés phytoplanctoniques. Ces problématiques très actuelles m’ont interpellée et ont constitué la base de mon projet de doctorat. Pour arriver à mes fins, des incubations expérimentales d’eau naturelle du Pacifique nord-est ont été menées lors de trois campagnes à bord du NGCC John P. Tully, chacune portant sur un thème différent. Les résultats de ces trois expérimentations sont décrits dans trois articles, formant les trois chapitres centraux de ma thèse.
Cette thèse a donc pour objectif, dans un premier temps, de vérifier la biodisponibilité du fer dans les poussières asiatiques et de quantifier la réponse biologique, y compris la production de DMS, de l'écosystème planctonique du Pacifique subarctique nord-est à l'ajout de poussières provenant du désert de Gobi selon un gradient de concentrations représentant des évènements de dépositions atmosphériques naturelles.
Le second objectif de cette thèse est de mesurer les quantités de TDFe et DFe relarguées par un ajout de cendres volcaniques et d’en quantifier les effets sur la communauté phytoplanctonique.
Le troisième objectif de cette thèse est de mesurer l’effet de l'acidification de l’océan sur la réponse biologique aux apports naturels et artificiels de fer, plus particulièrement sur la
croissance, la composition taxonomique et la production de DMS de la communauté planctonique.
Chapitre 2 – Experimental assessment of the impact of Asian
dust on phytoplankton dynamics and dimethylsulfide
production in the northeast subarctic Pacific
2.1 Résumé
Une communauté phytoplanctonique naturelle de la zone HNLC du Pacifique Nord-Est a été enrichie de poussières de désert asiatique. Les additions de poussières à une concentration de 0.5 mg L-1 et plus ont résulté en une augmentation significative de la concentration de chorophylle a, de carbone organique particulaire et de la prise de nitrate et silicate. Le taux de croissance en fonction de la concentration initiale de TDFe suit une équation de Monod avec une constante de mi-saturation de 0.33 mg L-1 et un taux de croissance maximal de 0.27 µgchla L-1d-1. Les concentrations de DMSPt et DMS n’ont pas été affectées par l’enrichissement, mais une diminution significative du ratio DMSPt:Chl a a été observée et attribuée au passage d’une communauté dominée par les prymnesiophytes, forts producteurs de DMSP, à une communauté dominée par les diatomées pennées, faibles productrices de DMSP. Cette étude démontre le potentiel fertilisant des poussières asiatiques.
2.2 Abstract
Dust deposition following episodic storm events represents an important source of iron (Fe) for High Nutrient - Low Chlorophyll (HNLC) oceanic regions. In this study, a 4-day incubation experiment was conducted to test the effect of a dust addition on nutrient uptake, phytoplankton speciation and growth, as well as on the production of dimethylsulfide (DMS) and its algal precursor dimethylsulfoniopropionate (DMSP) in the HNLC waters of the northeast subarctic Pacific. FeSO4 (final concentration of 0.6 nmol L-1) and Certified Reference Asian mineral dust (CJ-2) were added at different concentrations (0.12, 0.5, and 2 mg L-1) in 5 L incubation bags. FeSO
4 and dust additions exceeding 0.5 mg L-1 resulted in significant and comparable increases in nitrate and silicate drawdown, and in chlorophyll a (Chl a) and particulate organic carbon concentrations. The FeSO4- and dust-induced increases in biomass were accompanied by a general shift in phytoplankton dominance from Prymnesiophyceae to pennate diatoms. However, our results revealed phytoplankton group-specific responses to the mode of Fe delivery, with less centric diatoms, Cryptophyceae, Chrysophyceae and Prymnesiophyceae in the dust treatment than in the FeSO4 treatment. Variations in Chl a net accumulation rates as a function of the amount of dust added and initial total dissolvable iron (TDFe) exhibited a Monod-like model of growth. During the 4 days following dust addition, the half-saturation constant for net Chl a increase rate was 0.33 mg L-1 and the maximum increase rate value was 0.27 µg L-1d-1. TDFe proved to be a good predictor of microalgal Chl a net increase rate, with a half-saturation constant of 1.88 nmol L-1. DMSPt and DMS concentrations remained mostly unaffected by the addition of dust, but a significant decrease of the DMSPt/Chl a ratio was observed as a result of the shift in phytoplankton dominance from Prymnesiophyceae to
diatoms. Results from this study show that an Asian dust deposition event, corresponding to a typical monthly spring dust episode, may significantly stimulate phytoplankton nutrient uptake and growth and alter the structure of the phytoplankton assemblage in the northeast subarctic Pacific. Commonly dominant nanoplankton species are however less stimulated by dust than by the addition of FeSO4. Thus, FeSO4 represents a convenient but not perfect proxy for Fe-dust fertilization in HNLC regions.
2.3 Introduction
Vast areas of the world ocean are known as High Nutrient – Low Chlorophyll (HNLC). These regions, namely the Southern Ocean, the Equatorial Pacific, and the northeast subarctic Pacific, are characterized by low chlorophyll a (Chl a) concentrations in spite of an excess of macronutrients (nitrate, silicate, phosphate). The low biomasses measured in HNLC regions remained enigmatic until Martin and Fitzwater (1988) demonstrated in their canonical paper that iron (Fe) was limiting the growth of the phytoplankton in these areas. The Fe-limited status of the autotrophic and heterotrophic assemblages has been subsequently explored and confirmed in the three known HNLC regions through several small (bottles) and large scale Ocean Iron Fertilizations (OIFs; see reviews by Boyd et al., 2007; de Baar et al., 2005). These experiments have shown that Fe fertilization stimulates primary productivity and growth of most phytoplankton groups, with larger cells (diatoms) usually benefitting the most from the added micronutrient (Boyd et al., 2007 and references therein). By stimulating primary productivity and affecting the structure of phytoplankton communities, Fe fertilization has the potential to influence climate through carbon sequestration as well as through the dynamics of non-CO2 climate biogenic active gases such as dimethylsulfide (DMS).
The northeast subarctic Pacific HNLC waters are characterized by a dominance of prymnesiophytes and exceptionally high concentrations of the climate-active gas DMS in summer (Wong et al., 2005). DMS is the product of the enzymatic cleavage of dimethylsulfoniopropionate, a compound produced by most phytoplankton groups and potentially acting as osmolyte, cryoprotectant, antioxidant, chemotactic compound, and grazing deterrent (Kirst et al., 1991; Sunda et al., 2002; Tout et al., 2015; Van Alstyne et al., 2001). DMS production is the result of complex interactions involving biosynthesis of
DMSP by phytoplankton, bacterial assimilation of DMSP and DMS (Kiene et al., 2000; Simo et al., 2002), as well as DMSP-to-DMS conversion by both phytoplankton and bacteria (Kiene et al., 2000). Phytoplankton cell lysis due to grazing or viral attack is also known to stimulate DMS production (Dacey and Wakeham, 1986; Daly and DiTullio, 1996; Malin et al., 1998; Wolfe and Steinke, 1996). Fe fertilization has thus the potential to modify DMS/P dynamics by altering total biomass and community structure (selectively favoring high or low producers of DMSP) or by affecting the physiology of bacteria and phytoplankton. While previous OIFs conducted in the Southern Ocean and in the equatorial Pacific showed a consistent increase in DMS production following fertilization with FeSO4 (Turner et al., 2004), the sole OIF conducted in the northeast Pacific resulted in a decrease in DMS concentrations (Levasseur et al., 2006). The impact of Fe fertilization on DMS dynamics is thus variable and remains difficult to predict.
Sources of iron to the northeast subarctic Pacific include vertical mixing, eddies, tidal currents and convection (Cullen et al., 2009; Royer et al., 2010), volcanic ash (Mélançon et al., 2014; Olgun et al., 2011), and desert dust (Boyd et al., 1998; Jickells et al., 2005). An averaged 480 x 1012 g of mineral dust is delivered yearly, predominantly in spring during occasional dust storms originating from the deserts of northern Asia (Duce and Tindale, 1991). So far, none of the fertilization experiments conducted in the northeast Pacific used dust as a source of Fe. FeSO4, the artificial form of Fe used in these experiments, cannot be considered as a perfect proxy for dust given that Fe speciation in dust particles may alter its bioavailability (Breitbarth et al., 2010b; Shi et al., 2010) and that other compounds present in dust (e.g. macronutrients such as nitrate, phosphate and silicate and trace-metals such as copper) may either stimulate or inhibit algal growth (Mills et al., 2004; Paytan et al., 2009). Experiments with direct addition of Asian dust are thus still needed to assess the impact of these natural Fe deposition events on the northeast Pacific and to validate the results of previous bottle and large-scale experiments.
The general objective of this study was to determine the initial (first 4 days) bioavailability of iron contained in Asian dust to phytoplankton from the northeast subarctic Pacific. Specific objectives were 1) to determine the minimum dust concentration required to
induce a measurable response in terms of nutrient drawdown, phytoplankton biomass, and community composition, and 2) to assess how these variations affect the production of DMSP and DMS.
2.4 Material and Methods 2.4.1 Experimental settings
On deck incubations were conducted during a cruise along the Line-P transect aboard the Canadian Coast Guard Ship John P. Tully (Fig. 2.1). Water was collected at Ocean Station Papa (OSP – P26) (50°N, 145°W) from 5 m depth on June 14th 2009 using a Teflon® diaphragm compressed air activated pump with Teflon® tubing. Water was filtered through a 200 µm nylon mesh to remove large zooplankton and incubated in 5 L collapsible bags (Hyclone® Labtainers ™). A flow of surface water was continuously pumped through the incubators to keep temperature at in situ levels. Measured transmittance shows that the incubation bags filtered 55 % of ultraviolet A radiation, 70 % of ultraviolet B radiation and 33 % of photosynthetically active radiation (PAR), irradiance corresponding roughly to a depth of 10 m for the 400-600 nm wavelength in this part of the northeast Pacific (Sasaki et al., 2001). Incubations lasted 4 days and the bags were subsampled to measure a suite of chemical and biological variables (described below) at T0 (0-20 min after enrichment), at T2 (after 2 days) and at T4 (after 4 days, at the end of the incubation). Water was withdrawn from the bags by connecting a C-Flex® (Cole-Parmer) tubing to the Quick Connect port. Pressure was mechanically applied on the bag to fill the sampling tubing with water, hence preventing air from flowing into the bags. About 300 mL from each bag were flushed through the tubing prior to sampling. The bags were then suspended to a hook and sampled by gravity. Proper care was taken to prevent the back flush of water present in the tubing. Subsampling for Fe measurements took place in a trace-metal clean plastic tent supplied with positive pressure high-efficiency particulate air (HEPA). All materials in contact with seawater were cleaned to prevent trace-metal contamination according to protocols established by the international GEOTRACES program and available in GEOTRACES’ Methods Manual (Cutter et al., 2010).
Figure 2.1: Location of the sampling station (P26) along the Line P transect in the northeast subarctic Pacific Ocean. Graphic generated in Ocean Data View.
2.4.2 Treatments
Two sources of iron were used for the fertilization: FeSO4 and Certified Reference Asian mineral dust CJ-2. Briefly, CJ-2 dust was collected from the Tengger desert surface soil, in northern China, roughly sieved and blown through a wind tunnel designed to collect fine particles. Average median diameter of CJ-2 dust particles is 24.1 µm and its Fe content is 3.02 ± 0.12% (Nishikawa et al., 2000). Table 2.1 summarizes the 5 treatments and their correspondence to expected natural depositions. FeSO4 was added at the final concentration of 0.6 nmol L-1. CJ-2 dust was added following a gradient of concentrations of 0.12 (CJ2-L), 0.5 (CJ2-M) and 2 (CJ2-H) mg L-1. The first concentration, 0.12 mg L-1, represents a monthly average dust deposition during spring in the northeast subarctic Pacific, if confined to the upper 1 m of the water column (Zhao et al., 2003). The second concentration, 0.5 mg L-1, represents an estimate of the deposition for a full year in the northeast subarctic Pacific (0.5 g m-2), also confined to the first meter of the water column (Mahowald et al., 2005). Finally, 2 mg L-1 represents an extreme depostion event, corresponding, for example, to a strong Saharan dust storm in North Atlantic (Mahowald et al., 2005; Mills et al., 2004;