Validation des Dicer-substrate siRNA au niveau du système nerveux central dans le contexte de la recherche sur la douleur
Texte intégral
Figure

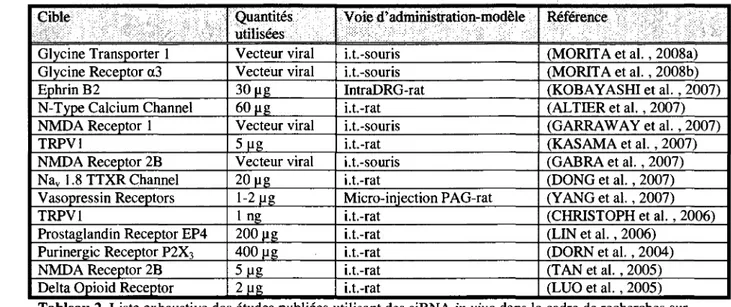

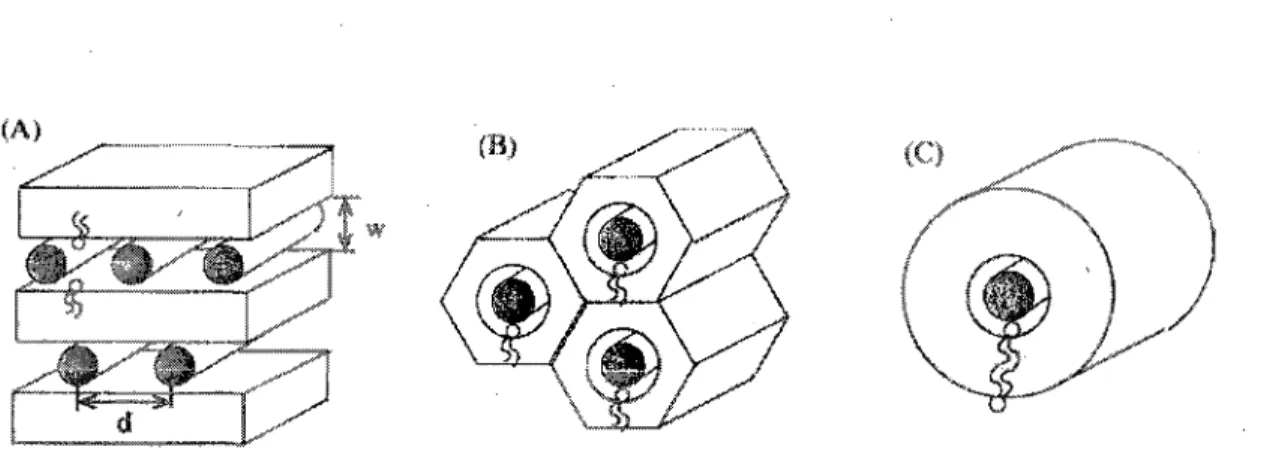
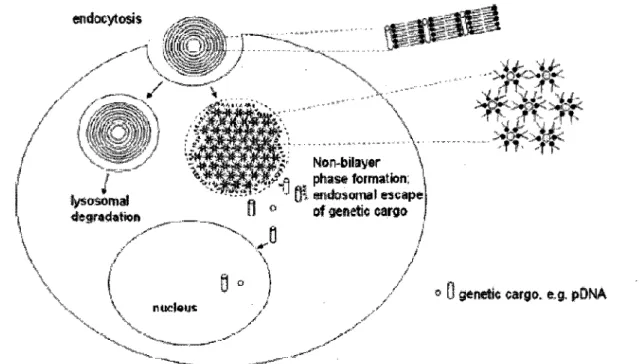




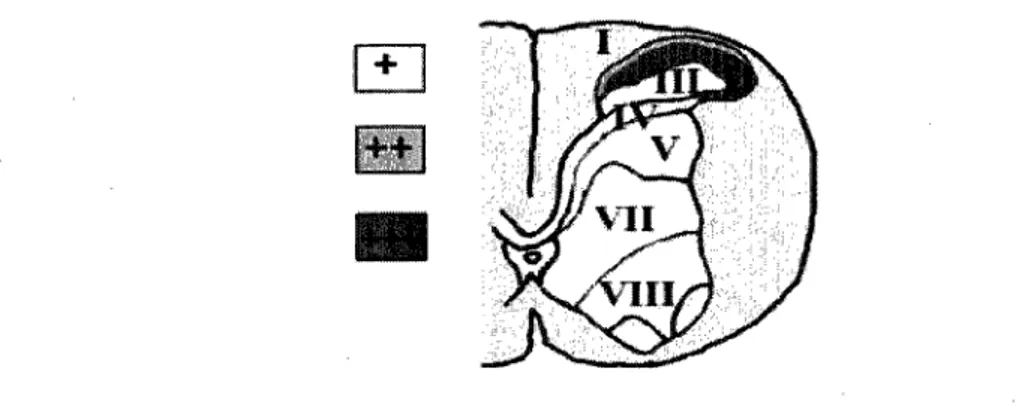
Documents relatifs
Les posologies particulières sont mentionnées en rouge dans le tableau. Pour les posologies usuelles voir p. 48-49 Références:.. o Clinical Practice Guideline for the diagnosis
Hundreds of pathogenic mtDNA variants implicated in a variety of human diseases ( Lott et al., 2013 ) have now been reported in the continuously updated Human Mitochondrial
Ceci revient à dire que des effets sanitaires pourraient être craints chez l’homme, si les mécanismes biologiques impliqués sont transposables, pour une valeur de DAS de 24
En effet, la comparaison entre le niveau général de vie de la population suisse et la précarité dans laquelle sont plongées les personnes à l’aide d’urgence accentue
La myélinisation est assurée par deux types cellulaires , les cellules d e Schwann e n périphérie e t les oligoden drocytes dans le système nerveux
On diffé rencie deux grands types de sys tèmes : les systèmes spécifiques - dans lesquels, schématiquement, un neurone transmet un message précis à un seul
lulaires de cette région projettent des fibres dans le cortex cérébral, l'hippocampe et le striatum. Enfin, un troisième groupe de péricaryons à POMC, récemment
Les puissances d’exposition ont été choisies par rapport à des puissances absorbées dans les tissus superfi ciels lors de l’utilisation d’un téléphone mobile : 6 W/kg :
