Both osteopontin-c and osteopontin-b splicing isoforms exert pro-tumorigenic roles in prostate cancer cells.
Texte intégral
Figure
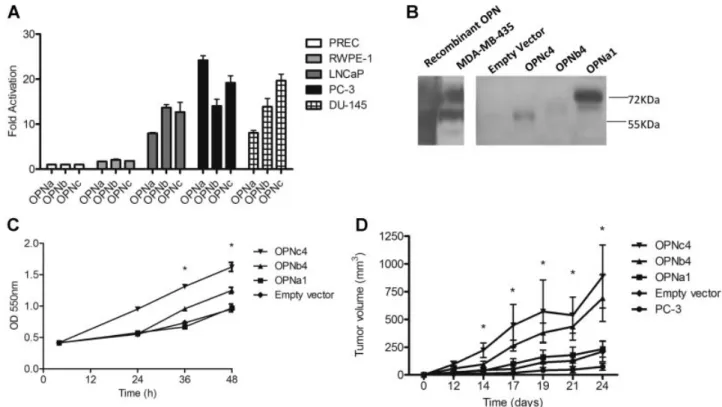
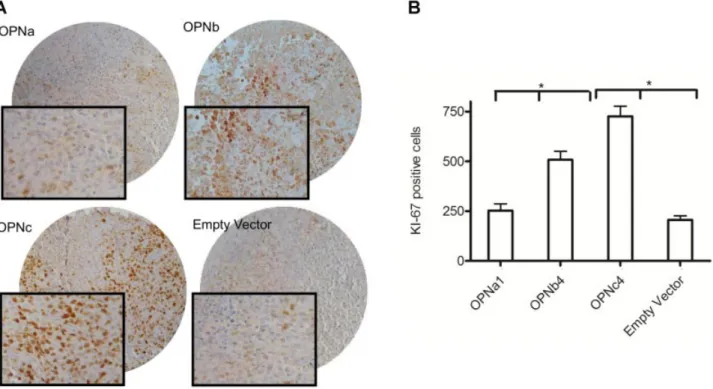
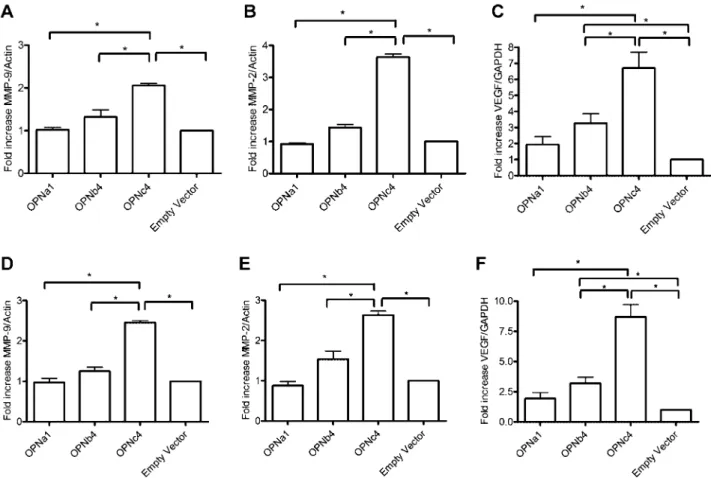
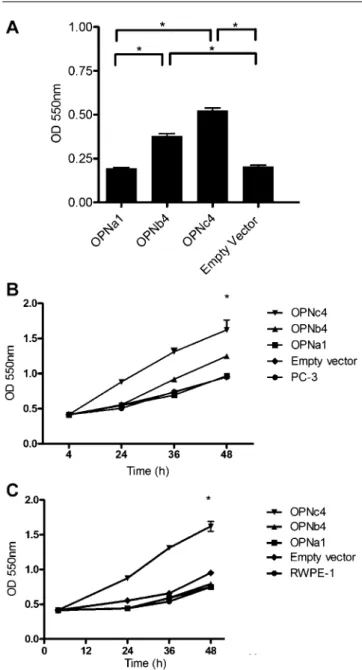
Documents relatifs
Northern blotting and quantitative RT-PCR analyses of MMP-2, TIMP-2, and MT1-MMP mRNA were performed on total RNA extracted from BZR and 16HBE14o- cells cultured with either
Nous posons que l’atelier philosophique pratiqué régulièrement : H’1 : développerait des prédispositions positives d’intégrité cognitive chez les philosophant
Copyright and moral rights for the publications made accessible in the public portal are retained by the authors and/or other copyright owners and it is a condition of
Moreover the measured and calculated softening of opti- cal phonon modes can be traced back to the existence of the hole pocket Fermi surface. Pa-
Pour lui, le gothique, « n’est pas un phénomène lié au temps mais, dans sa nature la plus profonde, à une race ; il est indépendant du temps, enraciné dans la
L’archive ouverte pluridisciplinaire HAL, est destinée au dépôt et à la diffusion de documents scientifiques de niveau recherche, publiés ou non,. ER beta inhibits proliferation
AgNOR staining also identified osteocyte canaliculi, cement and resting lines of the bone matrix and granules in the cytoplasm of the renal tubular cells.. In the present study,
