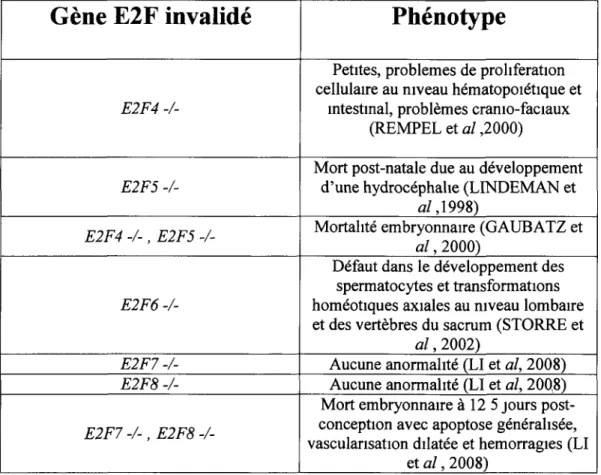
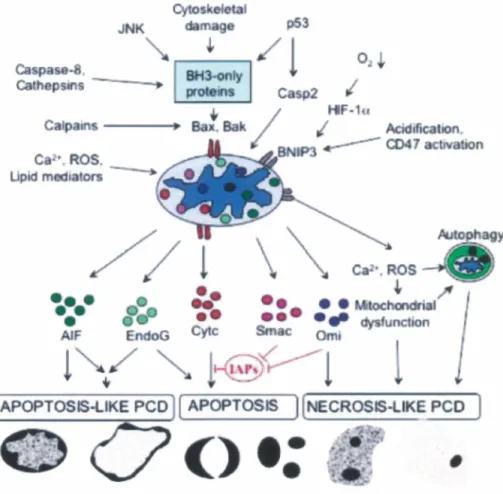
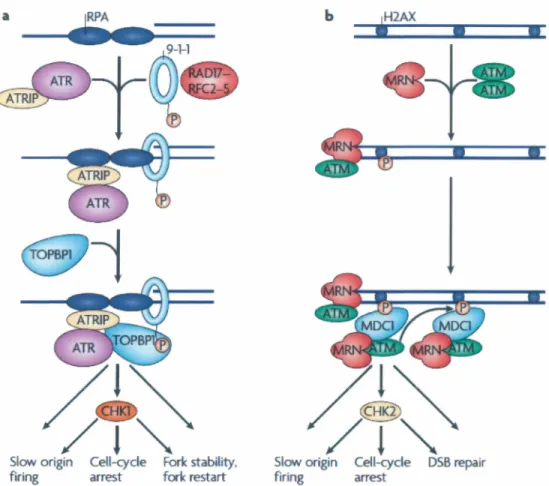
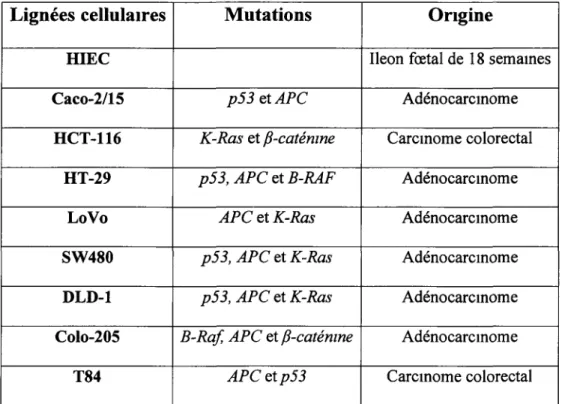
Implication du facteur de transcription E2F4 dans le contrôle du cycle cellulaire, de la survie et de la sénescence des cellules épithéliales intestinales humaines normales et cancéreuses
Texte intégral
Figure






Documents relatifs
To cite this version : Chauvet, Eric and Fustec, Eliane and Gas, Gilbert Etude expérimentale de la dégradation des lignines de Peuplier dans un sol alluvial et dans l'eau d'un
The DEM simula- tion was successfully used to model the behaviour of particles in a screw feeder and to obtain the highly inhomogeneous (although periodic in time) velocity field at
D'autres editeurs commerciaux : John Wiley & Sons, Blackwell Science Limited, Academic Press, Taylor & Francis, mettent leurs publications sur le reseau, soit en
D’une part (et c’est le point le plus décisif) le problème de l’émergence d’un schéma mental partagé concernant la configuration de l’organisation ( point 3/,
les poids à l'hectolitre élevés pour les lots récoltés avant les pluies et nettement plus faibles pour les lots récoltés après les pluies ;. les nombres de chute
In an opportunistic network where message delivery can sometimes be delayed significantly (at least for some receivers), this property of the OTR algorithm is an asset, for
osts of updating λ > 1 points will typi ally be greater than in the single point ase, unless every one out of λ omputational nodes is faster than the one applied in
