Tissue distribution of perfluorinated chemicals in harbor seals (Phoca vitulina) from the Dutch Wadden Sea
Texte intégral
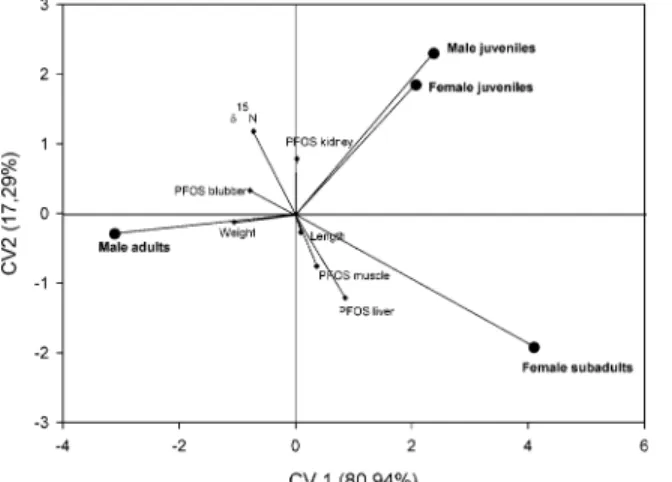
Figure

Documents relatifs
The mo;t efficient way to check the hairpin conformation con;I;t; in mea,uring the chain dimen;ion; in the direction, parallel (fl) and perpendicular (£) to the nematic director..
In a second part, we provide a feedback law yet inspired by sliding mode which not only stabilises the pure chain of integrators in prescribed time but also exhibits some robustness
The Multi Level Perspective of Geels (2002 and 2004) and the concept of Strategic Niche management (Kemp et al, 1998) provide a useful framework to describe or understand the
of dietary fibers, namely the short chain fatty acids (SCFAs) linking host nutrition to intestinal 26... SCFAs are an important fuel for intestinal epithelial cells (IEC) and
The mechanism has been written to reproduce the formation and consumption of formic, acetic, propionic and propenic acids under flame conditions; i.e.. the low- temperature
collective thinking and knowledge. The design can become an instrument that can transform the decision-making process and include the user as part of a cooperative development
Rath- er, the issue is with the definition of the activities carried out, the nature of certain activities at the harvesting stage (or rather the way it is practiced
3 restricts the study to polynomial spaces in one variable; in this case, a bound on the best approximation constant is derived and used to prove Theorem 3, which provides a rule
