Development of a highly specific sandwich ELISA for the detection of Listeria monocytogenes, an important foodborne pathogen
7
0
0
Texte intégral
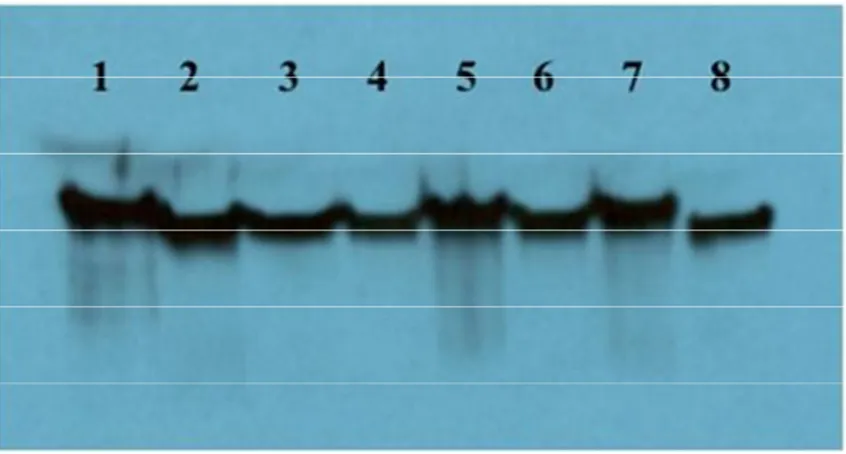
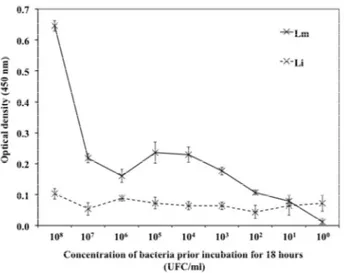
Figure
Documents relatifs
