Aqueous extraction of proteins from microalgae: Effect of different cell disruption methods
6
0
0
Texte intégral
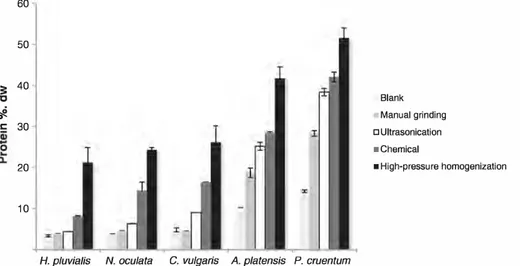
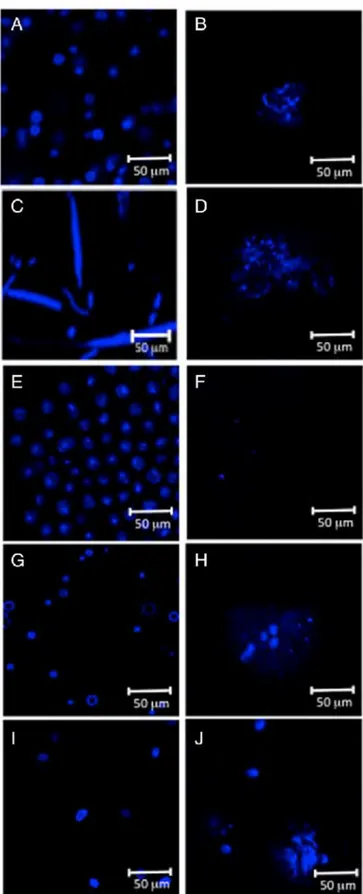
Figure
Documents relatifs