Évaluation de la reproduction des oiseaux
en forêt boréale aménagée
Thèse
Aude Catherine Corbani
Doctorat en sciences forestières
Philosophiae Doctor (Ph.D.)
Québec, Canada
iii
Résumé
L’objectif général de ce doctorat est d’évaluer le succès reproducteur d’une communauté d’oiseaux chanteurs en forêt boréale aménagée, et d’étudier certains facteurs qui lui sont associés. La méthode traditionnellement employée est le suivi de nids, mais elle reste laborieuse en forêt boréale où les nids sont généralement au sol ou cachés dans la cime fournie des arbres. Il existe d’autres approches pour documenter la reproduction sans trouver les nids. Elles s’appuient sur l’observation de certains comportements parentaux ou de jeunes à l’envol accompagnant leurs parents. Cependant, ces observations souffrent d’un biais de détection non négligeable, les parents ne présentant pas en permanence les indicateurs de succès. Le premier chapitre s’intéresse au succès de nidification (présence de jeunes au nid) et présente une méthode innovante permettant de mesurer le succès de nidification à partir d’observations de transport de nourriture en tenant compte du biais de détection du statut parental. Je montre que ce biais peut être important (la détection ne dépasse pas 38%), et que l’adaptation des modèles d’occupation (présence/absence) au succès de nidification est possible et efficace. Dans un second chapitre, j’applique cette méthode d’analyse à des observations de transport de nourriture récoltées depuis 1995 dans mon aire d’étude (Forêt Montmorency, Canada). Je montre que la tendance générale du succès de nidification au cours des 17 années est à la baisse, en lien avec une modification de l’habitat et des conditions météorologiques estivales qui se sont refroidies et asséchées. Également, le succès de nidification est meilleur dans les peuplements plus âgés et plus homogènes, de même que plus loin des chemins forestiers. Finalement, dans un troisième temps, je m’intéresse au succès d’envol (i.e. la proportion d’adultes ayant des jeunes à l’envol) et à sa détection. Je trouve que, dans la moitié des observations d’adultes sans jeunes à leurs côtés en fin de saison de reproduction, les individus sont en fait parvenus à produire des jeunes à l’envol. Je déduis de toutes les observations un succès d’envol de 59% que je n’ai pas pu relier aux caractéristiques du paysage (âge et homogénéité des peuplements). L’ensemble des résultats des trois chapitres vient appuyer l’idée que la forêt boréale est un lieu important pour les oiseaux en Amérique du Nord, et met en lumière l’importance du biais de détection lorsqu’on utilise des méthodes alternatives au suivi de nids pour mesurer le succès reproducteur.
v
Abstract
The aim of this PhD is to assess the reproductive success of a managed boreal forest songbird community, and to explore its causality. The traditional method to achieve this objective is nest monitoring, but it is laborious in boreal forest where nests are often on the ground or hidden at the top of trees. Alternative methods have already been used in boreal forest to document songbird reproduction without finding nests. They are based on observations of parental behavior (such as food provisioning) or fledglings accompanying their parents. However, these observations suffer from significant detection bias, parents not displaying success indicators all the time. In the three chapters of this thesis, I am interested in two stages of bird reproductive cycle: nesting success (i.e. the presence of young at the nest) and fledging success. The first chapter presents an innovative method to measure nesting success from food provisioning observations taking into account of parental status detection bias. I show that this bias may be significant (detection does not exceed 38%), and that the application of two-state occupancy models (presence/absence) to nesting success (young/no young at nest) is possible and effective. In a second chapter, I apply my analytic method to food provisioning observations collected since 1995 in the study area (Forêt Montmorency, Québec, Canada). I show that nesting decreases during the 17 studied years, in correlation with habitat modifications and meteorological changes over time. Additionally, nesting success is greater in older and more even-aged forests, as well as far from forestry roads and trails. In the third chapter, I show that, in half of the cases where adults are found without fledglings at the end of the reproductive season, they do in fact have fledglings. After accounting for this imperfect detection, I estimate fledging success as 59%, uncorrelated to landscape attributes (stand age and homogeneity). All results in this thesis support the claim that boreal forest represents an important breeding ground for birds in North America. I highlight the importance of detection bias when using alternative methods to nest monitoring in order to measure reproductive success and propose applications of these new tools for atlassing and similar large-scale ornithology initiatives.
vii
Table des matières
Résumé ... iii
Abstract ... v
Table des matières ...vii
Remerciements ... xv
Avant-propos ...xvii
Introduction générale ... 1
Le choix d’un habitat pour la nidification ... 2
Se reproduire dans son habitat ... 3
La difficulté d’estimer la reproduction ... 5
La phénologie et l’importance des conditions météorologiques et climatiques ... 6
La phénologie ... 6
La synchronisation ... 7
Le changement du climat ... 8
La fragmentation et la perte d’habitat ... 9
Les causes ... 9
Les conséquences ... 9
La modification des caractéristiques forestières ... 11
La gestion et l’aménagement forestiers au Québec ... 12
Objectifs et organisation de la thèse ... 14
CHAPITRE 1 ... 17
ESTIMATING NESTING SUCCESS: A NEW APPROACH INSPIRED FROM OCCUPANCY MODELS ... 17
Abstract ... 18
Résumé ... 19
INTRODUCTION ... 20
MATERIAL AND METHODS ... 21
Study area ... 21
Sampling parental provisioning ... 22
Application of two-state models to parental status ... 23
Validation of two-state models ... 25
RESULTS ... 28
viii
Proportion of adults with broods ... 31
Validation of two-state models ... 34
DISCUSSION ... 36
CONCLUSION ... 38
CHAPITRE 2 ... 41
A 17-YEAR ASSESSMENT OF SONGBIRD NESTING SUCCESS IN AN EASTERN CANADIAN BOREAL FOREST ... 41
Abstract ... 42
Résumé ... 43
INTRODUCTION ... 44
STUDY AND METHODS ... 45
Study area ... 45 Sampling protocol ... 46 Explanatory variables ... 48 Nesting success ... 51 RESULTS ... 52 DISCUSSION ... 59 CONCLUSION ... 63 CHAPITRE 3 ... 65
MEASURING SONGBIRD FLEDGING SUCCESS BASED ON IMPERFECT DETECTION OF FAMILY GROUPS ... 65
Abstract ... 66
Résumé ... 67
INTRODUCTION ... 68
MATERIAL AND METHODS ... 70
Study area ... 70
Survival time analysis ... 73
RESULTS ... 75 Overall productivity ... 75 Effect of covariates ... 76 DISCUSSION ... 77 CONCLUSION ... 79 Conclusion générale ... 81 Le succès de nidification ... 81
ix
La communauté vs. les différentes espèces ... 84
Les implications pour l’aménagement forestier ... 85
Conclusion et perspectives ... 86
Bibliographie ... 89
APPENDIX A: R code for songbird nesting success modeling with two-state occupancy models in Forêt Montmorency, Québec, Canada (chapter 1). ... 109
APPENDIX B: Total number of observations for each songbird species and number of cases where parental provisioning was observed for the period 1995-2011 in Forêt Montmorency, Québec, Canada (chapter 2). ... 112
APPENDIX C: Number of individuals per route count during Breeding Bird Surveys in Québec from 1995 to 2011 (Sauer et al. 2011) for the studied community (top; grey line represents mean number of individuals over the 17-year period.) and species analyzed in details (bottom) in chapter 2. ... 115
APPENDIX D: R code for survival modeling to assess fledging success in 2010 and 2011 of songbird community in Forêt Montmorency, Québec, Canada (chapter 3). ... 117
APPENDIX E: Number of individuals observed for each studied species at the end of the breeding season in 2010 and 2011 at Forêt Montmorency, Québec, Canada (chapter 3). . 121
APPENDIX F: Proportion of adults observed in each species (number of adults of the species/total number of adults observed) at the end of the breeding season in 2010 and 2011 (in black) and during the whole breeding season during the period 1995-2013 (in grey) at Forêt Montmorency, Québec, Canada (chapter 3). ... 123
xi
Liste des tableaux
Table 1. Assumptions of the MacKenzie et al. (2002) two-state model applied to the
estimation of site occupancy (species present or not) vs. parental status (brood present or not, given the presence of the species). ... 26
Table 2. Ranked two-state models of parental status () and its detection probability (p) at
site level. ... 29
Table 3. Estimated parameters (logit scale) and their standard error (SE) of parental status
() and its detection probability (p) for the best model ( (Full), p (Full)) at site level. ... 30
Table 4. Total number of observed adults for each species and number of cases where
parental provisioning was observed in Forêt Montmorency, Québec (Canada). ... 32
Table 5. Ranked two-state models of parental status () and its detection probability (p) at
species level. ... 33
Table 6. Estimated parameters (logit scale) and their standard error (SE) of parental status
() and its detection (p) for the best model ( (Julian day), p (Julian day)) at species level. . 33 Table 7. Relationship between probability of observing food provisioning, age of nestlings, time of day, temperature, hourly precipitation, and distance of parent to nest. Correlation between observed and fitted frequencies: r = 0.20 (n = 16 nests). ... 35
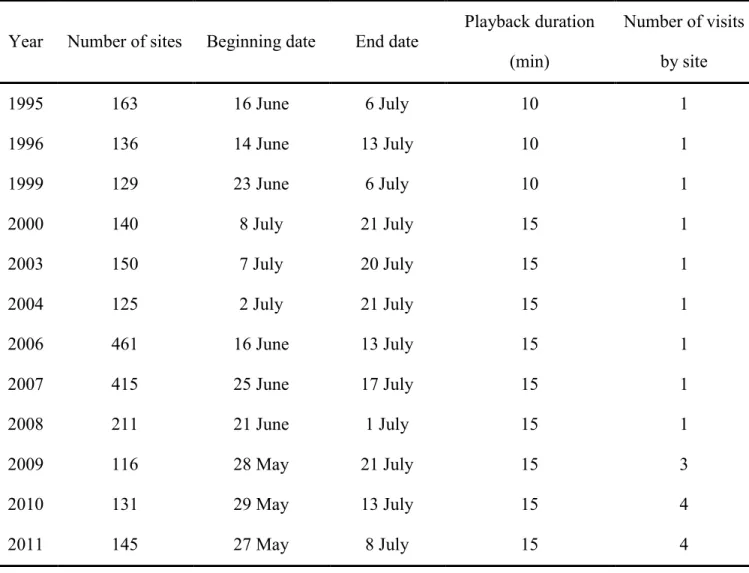
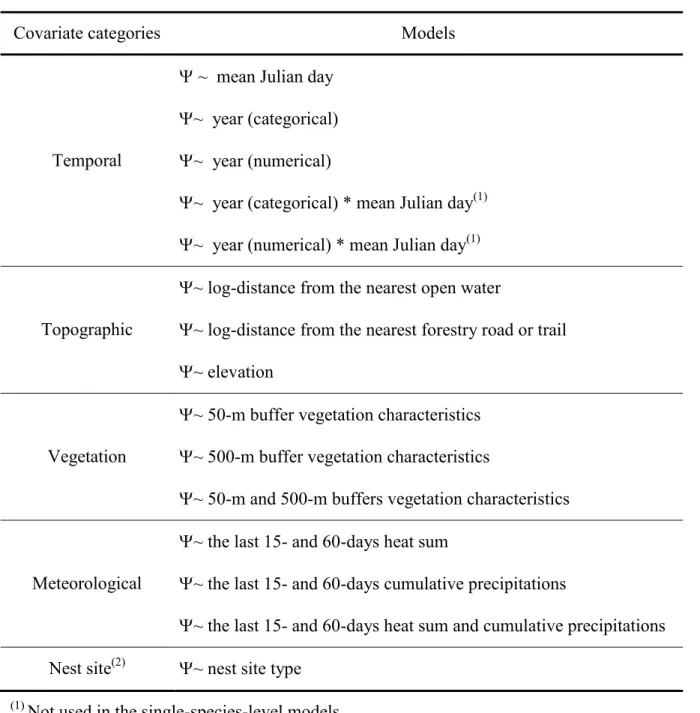
Table 8. Sampling conditions during the 12 studied years. ... 48 Table 9. Models considered for each level of analysis to analyze potential influence of
covariates on parental status (). Mean Julian day was included as a covariate of detection probability in every model (not presented here). ... 50
Table 10. Model averaged estimates (logit scale) ± standard error for each level of analysis,
i.e. species-, site- and single-species level. Blanks denote estimates that overlapped with zero. For readability, we omitted individual year effects (year as categorical variable), as well as all covariates with no estimates significantly different from zero. ... 55
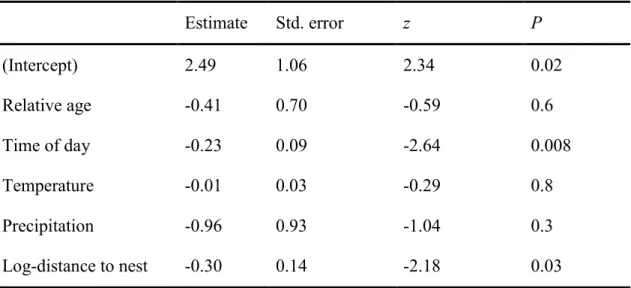
Table 11. Estimated parameters and their associated standard error (SE) for covariates of
time-to-detection and fledging success. Based on the z scores and an error rate of α = 5%, none of the estimates differed from zero. ... 77
xiii
Liste des figures
Figure 1. Domaines bioclimatiques du Québec méridional et localisation de la Forêt
Montmorency. ... 13
Figure 2. Exemple de mosaïque d’âge des peuplements à l’échelle du paysage à la Forêt
Montmorency. Source : André Desrochers. ... 14
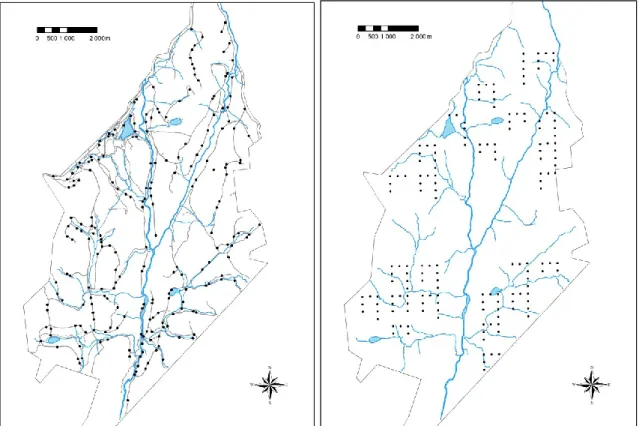
Figure 3. Sampling site locations on a) roads and trails, and b) systematic transects. Blue
lines and areas are rivers and lakes, grey lines, unpaved forestry roads and trails. ... 23
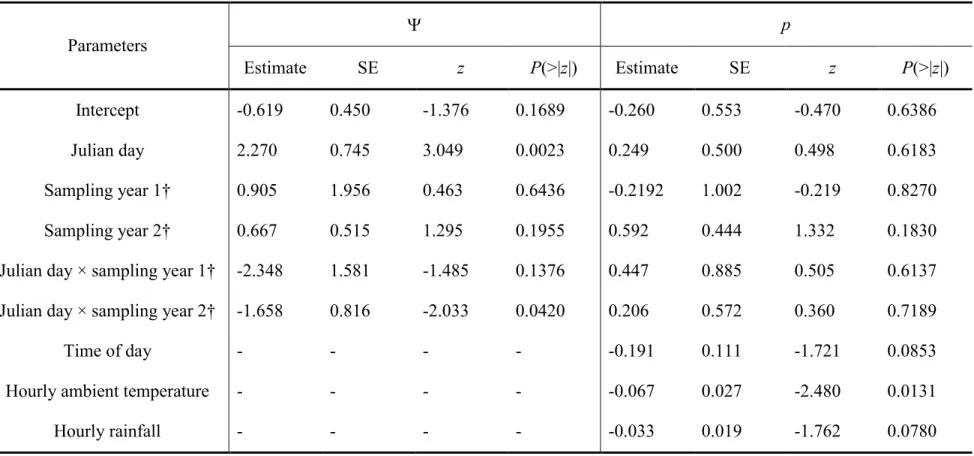
Figure 4. Estimated parental status () at species level as a function of mean Julian day (1st
January = day 1) in Forêt Montmorency, Québec (Canada). Dashed lines represent 95% confidence interval. ... 34
Figure 5. Study area with sampling sites in black, rivers and lakes in blue. The dashed line
represents the boundaries of Forêt Montmorency. ... 47
Figure 6. 17-y trend of meteorological conditions and habitat covariates in Forêt
Montmorency and neighboring areas, Quebec, Canada (1995-2011). Grey lines represent regression line. ... 53
Figure 7. Seasonal change in the proportion of adults with broods during the breeding
season at Forêt Montmorency and neighboring areas, Quebec, Canada. Based on a species-level analysis, 1995-2011. ... 54
Figure 8. Temporal trends of maximal proportion of adults with broods (species-level
analysis). Dashed lines represent gaps in sampled years. ... 56
Figure 9. Seasonal change in the proportion of adults with broods during the breeding
season for Yellow-rumped Warbler (YRWA, n=987), Golden-crowned Kinglet (GCKI,
n=610), Magnolia Warbler (MAWA, n=828), Blackpoll Warbler (BLPW, n=566) and
Black-throated Green Warbler (BTNW, n=558) at Forêt Montmorency and neighboring areas, Québec, Canada. ... 57
Figure 10. Influence of mean stand age (in years) and stand age homogeneity (in %) in a
50-m buffer around sampling sites on the proportion of adult songbirds with broods (species-level analysis). ... 58
Figure 11. Correlation between maximal parental activity each year and red squirrel
abundance at Forêt Montmorency, Québec, Canada. ... 59
Figure 12. Sampling site locations on roads and trails. Blue lines and areas are rivers and
lakes, gray lines, unpaved forestry roads and trails. ... 72
Figure 13. Individual selection procedure and data sets. Censored individuals corresponded
to adults for which we did not find any fledgling, and that were lost during the focal monitoring time or that were followed for the maximal focal monitoring duration. ... 73
Figure 14. Evolution of survival probability (Kaplan-Meier estimator) over focal bird
monitoring time (n = 206 monitored individuals). Dashed lines delimit the 95% confidence interval. ... 76
xv
Remerciements
La thèse de doctorat que j’ai le privilège de présenter ici est l’aboutissement d’un long cursus universitaire. Ce projet n’aurait pas été possible sans l’aide et la participation de plusieurs personnes que je tiens à remercier. Pour commencer, je remercie mon directeur de recherche, André Desrochers, pour sa confiance au départ d’accueillir une novice en ornithologie. Merci André pour avoir fait abstraction de ce « détail » et pour m’avoir permis de découvrir le monde merveilleux des oiseaux. Également, un grand merci pour tes précieux conseils et ton attention tout au long de ce doctorat.
Je voudrais également citer les différentes personnes qui se sont succédées sur le terrain depuis 1995 et qui ont permis d’engranger toutes les précieuses données ornithologiques que j’ai pu exploiter : Bruno Drolet (1995), Jean-François Rousseau et Jacques Ibarzabal (1996), Maude Pelletier (1999), Philippe Beaupré et Yan Cossette (2000), Bertrand LeGrand et Mélodie Lioret (2003), Sarah Overington et Jacinthe Tardif (2004), Christine Renaud (2004 et 2006), Ghislain Rompré (2006), Mathilde Jean-Saint-Laurent et Céline Macabiau (2007), Mélanie Major (2006 et 2008), Hermann Frouin et Johanne Theriault (2009), et André Desrochers (2004-2010). J’ai un remerciement particulier à faire à Fanny Senez-Gagnon et Josiane Bégin qui m’ont assisté au cours des campagnes de terrain de 2010 et 2011. Merci les filles pour votre aide, votre motivation et votre bonne humeur sur le terrain. J’ai découvert avec vous la forêt boréale avec ses superbes paysages, sa faune variée et ses insectes piqueurs !
Un clin d’œil également à mes amis étudiants du département qui, même s’ils n’étaient pas tous dans mon laboratoire, m’ont bien aidé et soutenu au cours de différentes étapes de mon doctorat, que ce soit pour des questions pointues en statistiques ou les moments de détente. Alors merci à Juliette, Ruth, Kaysandra, Arnaud, Matthieu, Sébastien, Christophe, Christian et tous les autres membres de l’Association des étudiants gradués de foresterie de l’Université Laval.
xvi
A Frédéric, mon compagnon, qui a accepté de traverser l’Atlantique avec moi pour découvrir le Québec. Merci d’avoir partagé cette expérience avec moi et de m’avoir épaulé dans cette grande étape de ma vie d’étudiante.
xvii
Avant-propos
Cette thèse de doctorat se compose de trois chapitres rédigés sous forme d’articles scientifiques qui peuvent être lus de façon indépendante les uns des autres. Chacun comprend une partie introductive exposant les objectifs, une partie méthodologique expliquant les procédures d’échantillonnage et d’analyse, une partie résultats, et enfin une partie discussion revenant sur les résultats et apportant une réponse à la question posée au départ. Une introduction et une conclusion générales permettent de présenter ce qui a motivé cette thèse ainsi que le fil conducteur reliant les différents chapitres. Je suis l’auteur principal de tous les chapitres, et mon directeur de recherche, André Desrochers, en est le co-auteur pour m’avoir encadré à toutes les étapes de mon projet doctoral. Également, Marie-Hélène Hachey (étudiante à la Maîtrise 2010-2011) est coauteur de mon premier chapitre pour avoir apporter des données et des analyses substantielles permettant une bonification de ce chapitre. Finalement, M’Hamed Lajmi Lakhal-Chaieb est co-auteur du troisième chapitre pour sa participation à la mise en place et à la réalisation des analyses de survie.
Chapitre 1
Corbani, A.C., M.-H. Hachey and A. Desrochers. Estimating nesting success: a new approach inspired from occupancy models. Soumis à la revue Avian Conservation and
Ecology - Ecologie et conservation des oiseaux.
Chapitre 2
Corbani, A.C. and A. Desrochers. A 17-year assessment of songbird nesting success in an eastern Canadian boreal forest.
xviii
Chapitre 3
Corbani, A.C., A. Desrochers and M.L. Lakhal-Chaieb. Measuring songbird fledging success based on imperfect detection of family groups. Soumis à la revue Journal of Field
Ornithology.
Coauteurs des chapitres
André Desrochers : Département des sciences du bois et de la forêt, Université Laval, Québec, Québec, Canada, G1K 7P4
Marie-Hélène Hachey : Regroupement QuébecOiseaux et Atlas des oiseaux nicheurs du Québec, Québec, Québec, Canada, G1J 0C3
M’Hamed Lajmi Lakhal-Chaieb : Département de mathématiques et statistiques, Université Laval, Québec, Québec, Canada, G1K 7P4
1
Introduction générale
La forêt boréale, ou encore taïga, représente près de la moitié des grandes forêts intactes de la planète (Potapov et al. 2008). Elle est composée de grandes forêts dominées par les conifères comme les pins, les épinettes, les mélèzes ou encore les sapins, et constitue une véritable mosaïque de paysages avec ses nombreux lacs et tourbières. La forêt boréale joue un rôle essentiel dans le cycle du carbone à l’échelle mondiale (Pan et al. 2011), et a ainsi un énorme potentiel pour affecter le changement climatique global malgré le peu d’attention qu’on a pu lui porter en terme de gestion et de conservation de la biodiversité (Bradshaw et al. 2009, Warkentin & Bradshaw 2012). En Amérique du Nord, la forêt boréale représente le plus vaste écosystème forestier et s’étend de l’Alaska à Terre-Neuve-et-Labrador, globalement entre le 45e et le 57e parallèle Nord même si elle peut dépasser le cercle polaire par endroits.
Au Québec, la zone boréale représente plus d’un million de kilomètres carrés, et se divise en plusieurs domaines bioclimatiques (Ministère des Ressources naturelles 2013b). Parmi eux, la sapinière à bouleau blanc représente 13% de la superficie de la zone boréale. Elle est dominée par des peuplements de sapins baumiers (Abies balsamea, (L.) Miller) et d’épinettes blanches (Picea glauca, (Moench) Voss), accompagnés de bouleaux blancs (Betula papyrifera, Marshall) sur les sites mésiques. Sur les sites moins favorables, l'épinette noire (Picea mariana, (Mill.) Britton), le pin gris (Pinus banksiana, Lamb.) et le mélèze (Larix laricina, (Du Roi) Koch) sont souvent en mélange avec des bouleaux blancs ou des peupliers faux tremble (Populus tremuloides, Michx.). Le bouleau jaune (Betula
alleghaniensis, Britton) et l'érable rouge (Acer rubrum, L.), eux, ne se retrouvent que dans
la partie la plus méridionale du domaine.
La forêt boréale canadienne constitue un lieu de reproduction important pour de nombreuses espèces d’oiseaux. D’après Blancher (2003), près de trois cents espèces se reproduisent en forêt boréale canadienne, représentant entre 1,5 et 3 milliards d’individus reproducteurs chaque année. D’une manière générale, les populations d’oiseaux nicheurs au Canada ont diminué de 12% depuis 1970 (North American Bird Conservation Initiative
2
Canada 2012). Les groupes les plus touchés ont été les espèces prairiales, les insectivores aériens et les oiseaux de rivage. Dans la région boréale de l’Est, qui s’étend de l’Ontario à la pointe nord-est de Terre-Neuve-et-Labrador, les populations d’oiseaux forestiers sont restées relativement stables et représentent près de la moitié des espèces caractéristiques de la région boréale de l’Est (North American Bird Conservation Initiative Canada 2012). Le déclin de certains groupes est lié à la fois aux conditions rencontrées au cours de leur période de nidification, mais également, pour les migrateurs, aux conditions rencontrées tout au long de leur cycle vital, c’est-à-dire au cours de la migration et sur les aires d’hivernage (Newton 2004). Le manque d’information sur la qualité des habitats en dehors de la période de reproduction a d’ailleurs été plusieurs fois cité comme une priorité de recherche pour la conservation, de même que la nécessité d’améliorer les suivis à long terme (Donovan et al. 2002, Faaborg et al. 2010).
Le choix d’un habitat pour la nidification
La sélection des habitats consiste pour les oiseaux adultes à choisir un site pour la période de nidification qui offrira la meilleure survie et la meilleure reproduction. Deux théories sont reconnues pour tenter de comprendre la répartition des oiseaux : la distribution libre idéale et la distribution despotique idéale (Fretwell & Lucas 1969). La première théorie suppose que les oiseaux sont libres de choisir leur territoire et prédit qu’ils s’établissent en priorité dans les habitats de meilleure qualité. Lorsque la densité en individus dépasse un certain seuil (capacité de charge du milieu), l’effet de masse se fait ressentir (Sinclair 1989, Murdoch 1994) et la qualité de l’habitat diminue jusqu’à atteindre celle des habitats de moindre qualité ; ces habitats de qualité inférieure commencent alors à être occupés (Rodenhouse et al. 1997).
La seconde théorie (distribution despotique idéale) s’appuie sur le principe de territorialité. Lorsque les individus ont sélectionné un habitat, ils doivent se définir un territoire avec les informations disponibles à ce moment là, et délimiter ce territoire qui sera défendu et utilisé pour l’accouplement, la nidification et comme site de nourrissage pour les jeunes. Le territoire est une zone dans le domaine vital de chaque individu dans laquelle il aura
3 l’exploitation exclusive ou prioritaire ; le domaine vital, lui, correspond à une zone traversée de manière répétée où un individu a une probabilité d’occurrence prédéterminée pendant une période donnée (Powell 2000, Kernohan et al. 2001). En règle générale, les individus plus âgés arrivent en premier et s’octroient les territoires de meilleure qualité (Holmes et al. 1996), laissant les plus jeunes ou les nouveaux individus avec des territoires de moins bonne qualité et des domaines vitaux généralement plus vastes.
Dans certains cas, les individus choisissent des habitats par fidélité au site ou par une mauvaise lecture des signaux mais ces habitats ne sont pas optimaux en termes de reproduction et/ou de survie ; on parle alors de pièges écologiques (Donovan & Thompson 2001, Schlaepfer et al. 2002, Battin 2004). Également, de nombreux individus peuvent être contraints de se retrouver dans des territoires de moins bonne qualité, soit par manque d’information sur l’aire de reproduction comme c’est le cas pour les plus jeunes individus reproducteurs (Fretwell & Lucas 1969), soit par un accès limité ou une compétition intra- ou interspécifique à leur désavantage (Rodenhouse et al. 1997).
Se reproduire dans son habitat
Une fois le type d’habitat sélectionné, la période de nidification peut débuter. Elle est constituée de plusieurs étapes conditionnant le succès reproducteur. Après avoir établi leur territoire, les mâles devront s’apparier avec une femelle. Un nid est ensuite construit qui permettra l’installation du couple, et la ponte ainsi que la couvaison des œufs. Après éclosion des œufs, les jeunes seront nourris par les adultes. L’étape finale du cycle reproducteur annuel sera l’envol des juvéniles. Le succès relatif de chacune de ces étapes peut varier selon l’espèce considérée, la région étudiée ou en encore les perturbations du paysage. En forêt boréale, Dalley et al. (2009) ont évalué que, dans leur zone d’étude au Nouveau-Brunswick, le succès d’éclosion était supérieur à 92%, le succès au nid pouvait atteindre 85%, et le succès d’envol était de 82 % pour trois espèces de passereaux. Dans un contexte de fragmentation de l’habitat, chaque étape peut être affectée par la modification du paysage, mais le succès d’appariement semble être l’aspect le plus limitant (Lampila et al. 2005).
4
Le succès reproducteur, défini ici comme la production par un individu d’une descendance viable, peut se mesurer de différentes façons (Etterson et al. 2011). La plus populaire est la recherche et le suivi de nids afin de mesurer différents paramètres tels que le succès d’éclosion (p.ex., Dalley et al. 2009), la taille des pontes (p.ex., Winkel & Hudde 1997), la survie au nid (p.ex., Donovan et al. 1995b), la condition physique des oisillons (p.ex., King et al. 1996) ou encore le succès d’envol (p.ex., Hoover et al. 1995). Le nombre d’études sur le succès reproducteur en forêt boréale reste relativement faible par rapport à d’autres biomes comme la forêt tempérée, et peu de choses sont connues sur les différentes étapes de la période de reproduction. Une des raisons qui explique ce manque d’informations est la difficulté à suivre les nids. Il est en effet laborieux et coûteux en temps de trouver des nids, notamment dans les futaies de conifères, car ils sont la plupart du temps localisés dans la cime fournie des arbres. Bien que le suivi de nids reste la mesure la plus directe du succès de reproduction, cette méthode demande un grand effort d’échantillonnage, elle est restreinte à des territoires de faible superficie, et elle peut nécessiter plusieurs années de terrain pour amasser des échantillons valables (Dececco et al. 2000, Fauth 2000, Willson & Gende 2000). De plus, malgré l’échantillonnage intense nécessaire, le suivi de nids peut mener à des biais d’estimation du succès de nidification (Ball & Bayne 2012).
Il existe d’autres approches déjà utilisées avec succès en forêt boréale pour documenter la reproduction sans trouver les nids. Parmi elles, il y a l’estimation du succès d’appariement, c’est-à-dire la proportion de territoires occupés par des couples (p. ex. Villard et al. 1993, Sabine et al. 1996, Lambert & Hannon 2000, Bayne & Hobson 2001a). Il y a également l’observation de la présence ou non de juvéniles avec les adultes (p. ex. Rangen et al. 2000, Griesser et al. 2007, Vitz & Rodewald 2010) et leur abondance relative dans les captures au filet en fin de saison de reproduction (p.ex. DeSante & Kaschube 2007). L’activité parentale, comme le transport de nourriture au bec indiquant la présence de jeunes à alimenter, peut également être utilisée comme indicateur du succès de nidification des oiseaux (p. ex. Vickery et al. 1992, Gunn et al. 2000, Ibarzabal & Desrochers 2005).
À ce jour, la majorité des études extensives sur la reproduction des oiseaux en forêt boréale a été sur de courtes périodes (entre un et trois ans). Pourtant, seules les études à long terme
5 peuvent permettre une compréhension de l’interaction entre productivité et facteurs environnementaux comme le changement climatique (p.ex. Sillett et al. 2000, Both et al. 2004, Torti & Dunn 2005) ou encore la modification du paysage (p.ex. Holmes & Sherry 2001, Zuckerberg & Porter 2010). Les études publiées à date suggèrent que le succès de nidification est généralement élevé en forêt boréale (Doran et al. 2005, Ibarzabal & Desrochers 2005), mais les conclusions sont généralement de portée limitée en raison de la courte durée de ces études mais également du fait qu’elles s’appuient sur des indices plutôt que d’estimations directes du phénomène d’intérêt (Royle & Dorazio 2008).
La difficulté d’estimer la reproduction
La vision classique en écologie statistique et en biologie des populations est d’estimer ce qui est observé par échantillonnage (Royle & Dorazio 2008), le plus fréquemment à l’aide d’indices. Les méthodes présentées plus haut s’inscrivent dans cette logique traditionnelle, en procurant généralement des indices de reproduction (Rangen et al. 2000, Betts et al. 2005b). Or, les indices tels que la proportion d’oiseaux vus avec des jeunes sont le résultat de deux processus confondus : l’échantillonnage imparfait et l’état réel du système étudié. Même si les indices peuvent parfois être calibrés (Bonifait et al. 2006), ils ont une utilité limitée quand vient le temps de décrire objectivement un système écologique tel qu’une population d’oiseaux en reproduction. Les indices qui confondent échantillonnage et état objectif du système s’avèrent particulièrement inefficaces lorsque la détection des phénomènes étudiés tels que la présence ou la reproduction d’une espèce est elle-même influencée par une gamme de facteurs, et c’est cette problématique qui fait l’objet de ma thèse de doctorat.
Dans le cas du transport de nourriture, les parents ne passent pas cent pour cent de leur temps à approvisionner leurs poussins. Ils répartissent leur temps entre différentes activités comme le toilettage ou encore le repos. Ainsi, un adulte qui a des jeunes au nid peut être observé sans nourriture au bec malgré son statut parental, et l’estimation du succès de nidification basée sur de telles observations est alors biaisée négativement (Rivers et al. 2003, Morgan et al. 2010). On peut constater le même biais lorsqu’on s’intéresse cette fois
6
aux jeunes à l’envol. Dans les premiers temps après l’envol, les jeunes restent avec leurs parents qui leur procurent encore des soins parentaux comme le nourrissage ou encore la protection contre les prédateurs (Lack 1968). Pendant cette période, il est possible d’observer des groupes familiaux constitués d’un ou de deux parents accompagnés par un ou plusieurs jeunes. Un problème majeur est que l’observation des jeunes est difficile, en particulier dans les peuplements forestiers denses souvent retrouvés en forêt boréale (Rangen et al. 2000), d’autant plus que ces habitats seraient préférentiellement sélectionnés par les jeunes (King et al. 2006).
La phénologie et l’importance des conditions météorologiques et
climatiques
La phénologie
Le moment où se déroule la reproduction est l’un des facteurs les plus importants qui déterminent la performance reproductrice des oiseaux. Des indicateurs proximaux (utilisés pour prédire et préparer la saison de reproduction) et ultimes (exerçant une pression évolutive sur les oiseaux pour nidifier au moment de l’année qui favorise le succès reproducteur maximal) sont utilisés par les oiseaux afin de d’optimiser la période à laquelle ils se reproduisent (Baker 1938, Visser et al. 2010). Comme dans d’autres groupes de vertébrés, de nombreuses espèces d’oiseaux utilisent la photopériode comme indicateur proximal (Dunlap et al. 2004). La photopériode a à la fois un effet physiologique en influant par exemple sur le développement des gonades (Dawson et al. 2001, Ball & Balthazart 2003), mais également un effet sur le comportement des oiseaux qui vont initier leur recherche de partenaire et leur parade nuptiale (Dawson et al. 2001). La température ambiante est également utilisée comme indicateur proximal, et va aider à la décision de l’instant auquel débuter le cycle de reproduction, c’est-à-dire la date de ponte (Perrins 1965, Dunn 2004, Visser et al. 2009). En général, le moment de la reproduction (timing of
7
La synchronisation
Les individus qui nichent plus tôt, le plus souvent les adultes les plus âgés (Desrochers 1992), produisent généralement des couvées plus grandes et avec une meilleure survie que les individus qui nichent plus tardivement (Perrins & McCleery 1989, Price & Liou 1989, Dunn et al. 2011). Il est à noter que la date optimale de ponte peut dépendre d’autres paramètres du cycle de reproduction comme le nombre de couvées par saison (Crick et al. 1993, Visser et al. 2003). La période de nidification est celle à laquelle les demandes énergétiques sont certainement les plus importantes, avec une demande en approvisionnement des poussins qui augmente avec leur âge (Pinkowski 1978, Johnson & Best 1982, Bédard & Meunier 1983, Goodbred & Holmes 1996, Reed et al. 2007, Geiser et al. 2008, Barba et al. 2009) et la taille de la couvée (Johnson & Best 1982, Wright et al. 1998, Rauter et al. 2000). La concordance entre disponibilité en nourriture et moment de la reproduction apparaît alors comme un élément essentiel. Chez de nombreuses espèces d’oiseaux, la date de ponte se synchronise de manière à établir une correspondance entre le pic de croissance des oisillons et le pic de disponibilité en nourriture (Lack 1966, Perrins 1970, Vatka et al. 2011), et la modification de la quantité de nourriture disponible peut faire varier la date de ponte ainsi que la taille des couvées (Newton & Marquiss 1981, Arnold 1992, Kallander & Karlsson 1993). Une mauvaise synchronisation entre le moment de la reproduction et une abondance élevée en nourriture peut engendrer une survie et une qualité des jeunes plus faibles et un succès reproducteur moins élevé (Thomas et al. 2001, Visser et al. 2006, Vatka et al. 2011).
Outre son rôle de facteur proximal, la température ambiante, mais également les précipitations, sont des déterminants importants pour la disponibilité en nourriture. Des conditions météorologiques printanières médiocres (surtout des températures basses) peuvent affecter la disponibilité en nourriture, comme la biomasse de chenille, et diminuer la productivité des oiseaux (Visser et al. 2006, Vatka et al. 2011). Le manque de nourriture mène à une croissance plus faible des oisillons, un envol plus tardif et une plus grande mortalité par inanition (McNamara & Houston 1987, Wright et al. 1998, Dunn 2004). La température pendant la période où les jeunes sont au nid change la proportion d’insectes volants attrapés par les parents, et la fréquence de nourrissage augmente avec la
8
température (Veistola et al. 1997). La survie au nid peut également être réduite si la disponibilité en nourriture est faible car les parents passent plus de temps à rechercher de la nourriture et moins à défendre leur nid (Pearse et al. 2004, Dunn et al. 2010), surtout si les conditions sont froides et causent de l’hypothermie chez les jeunes au nid (Stevenson & Bryant 2000).
Le changement du climat
Depuis le début du XXème siècle, la température de l’air au sol a augmenté en moyenne de 0.6°C, et plusieurs spécialistes du climat s’attendent à ce que cette augmentation se poursuive à un taux plus fort encore au cours des cent prochaines années (Intergovernmental Panel on Climate Change 2007). Le changement du climat engendre d’importantes conséquences pour les populations d’oiseaux. En Amérique du Nord, on a observé une remontée vers le Nord (1.48 km par an pour la période 1975-2004) des aires d’hivernage (La Sorte & Thompson 2007), possiblement associée aux changements climatiques récents. L’augmentation de la température modifie sur le long terme la phénologie de la reproduction avec un avancement général des dates de ponte et d’éclosion (Winkel & Hudde 1997, Brown et al. 1999, Crick & Sparks 1999, Dunn & Winkler 1999, Both & Visser 2001, Both et al. 2004, Torti & Dunn 2005). Ce changement peut augmenter la taille des couvées (Järvinen 1989, Winkel & Hudde 1997) et engendrer un meilleur succès d’envol (Winkel & Hudde 1997). Cependant, les conditions d’élevage des jeunes se détériorent avec l’augmentation de la taille des couvées. Plus il y a de jeunes à nourrir par couvée, plus la qualité de l’approvisionnement par oisillon est faible (Wright et al. 1998). Les jeunes dans des grosses couvées subissent alors des taux de croissance plus faibles et possèdent une masse corporelle à l’envol plus faible que des jeunes issus de couvées plus petite (Wright et al. 1998), ce qui engendrerait une plus forte mortalité après l’envol (Magrath 1991, Vitz & Rodewald 2011). Le principal problème pour les populations d’oiseaux est la vitesse à laquelle se produit le changement de climat (Miller-Rushing et al. 2010), vitesse qui a néanmoins diminué cette dernière décennie (Otto et al. 2013). La persistance des populations d’oiseaux dépendra alors de leur capacité d’adaptation écologique et évolutive (Cox 2010).
9
La fragmentation et la perte d’habitat
Les causes
Depuis le début de l’ère industrielle au Québec, la forêt boréale a subi de profondes modifications. L’exploitation forestière et minière, de même que les aménagements nécessaires à l’exploitation des ressources naturelles tels que le réseau routier ou encore le développement des lignes de transports d’énergie, ont façonné les paysages (Desponts 1995). En forêts mixte et coniférienne, les peuplements de bouleaux blancs et peupliers faux-trembles ou les peuplements denses de jeunes sapins (selon l’intensité de coupe) ont remplacé les peuplements d’origine (Desponts 1995). La surface forestière a été de manière générale réduite (perte d’habitat) et il en a résulté un morcellement des différents éléments du paysage (fragmentation).
Les conséquences
La conséquence directe de la fragmentation et de la perte d’habitat est la perturbation de la dynamique source-gouffre (source-sink) des populations d’oiseaux (Donovan et al. 1995a, Donovan et al. 1995b, Robinson et al. 1995, Lloyd et al. 2005). D’après Andrén (1994), le premier impact sur les populations vient de la perte d’habitat qui diminue la quantité d’habitats appropriés ; ensuite vient l’effet de la taille et de l’isolement des fragments rémanents qui influence la taille de la population. Ainsi, la fragmentation seule n’expliquerait pas la diminution de la tailles des populations (Villard 2002), et les conséquences dépendraient des préférences des espèces pour certains habitats (Bender et al. 1998). Un habitat est défini comme source si le taux de croissance de la population d’intérêt, λ, est supérieur à 1, un gouffre étant un habitat où λ < 1 (Pulliam 1988, Dias 1996). La dynamique de ce système source-gouffre consiste en des variations des paramètres démographiques dans chacune des populations et en des échanges entre les populations d’un même ensemble, appelé métapopulation (Levins 1970).
Avec la fragmentation et la perte d’habitat, ce qui était une forêt continue devient des fragments forestiers résiduels, souvent de plus en plus petits et de plus en plus isolés. Lorsque la taille des fragments diminue, la densité des populations d’oiseaux (Schmiegelow
10
et al. 1997) mais également l’abondance des prédateurs (Chalfoun et al. 2002, Koprowski 2005) augmenteraient, au moins temporairement. Le succès d’appariement est l’étape du cycle de reproduction qui est la plus affectée par la fragmentation (Lampila et al. 2005), et la diminution de la taille des fragments réduirait le succès d’appariement (Villard et al. 1993, Burke & Nol 1998, Lee et al. 2002) ainsi que le succès au nid (Burke & Nol 2000, Stephens et al. 2003). L’approvisionnement des jeunes en nourriture peut parfois être perturbé (Zanette et al. 2000, Luck 2003), agissant ainsi sur la condition physique des oisillons (Hinsley et al. 1999, Huhta et al. 1999). La réduction de la taille des fragments agit également parfois sur la richesse spécifique, et la composition des communautés d’oiseaux devient de plus en plus variable (Schmiegelow et al. 1997, Boulinier et al. 1998a, 2001). La compensation des pertes liées aux gouffres par les habitats sources dépend de la taille de la source et de la fidélité au site (Donovan et al. 1995a). Chez les passereaux, la fidélité au site est typiquement élevée et la dispersion natale est très extensive (Tittler et al. 2006, Tittler et al. 2009). D’après Donovan et al. (1995a), si la source a un taux de croissance faible et que la fidélité est grande ou si la source a une croissance forte, alors la population arrive à se maintenir en supposant que la fragmentation ne soit pas trop importante.
L’isolement des îlots forestiers rémanents apparaît alors comme un facteur déterminant pour la dynamique de métapopulation et le maintien des populations dans les habitats gouffres. A la fois la théorie de biogéographie des îles (MacArthur & Wilson 1963, MacArthur & Wilson 1967) et la théorie de la métapopulation (Levins 1970) prédisent une perte de populations due à la fragmentation de l’habitat causée par une plus grande probabilité d’extinction et une plus faible probabilité de recolonisation des habitats isolés. La connectivité entre les fragments sources et gouffres conditionne alors la viabilité des populations (With et al. 2006).
Une conséquence indirecte de la fragmentation pour les oiseaux est la modification de la communauté de prédateurs (Schmiegelow & Mönkkönen 2002), et l’augmentation de la prédation et du parasitisme des couvées (Robinson et al. 1995, Chalfoun et al. 2002, Lloyd et al. 2005, Thompson 2007). En modifiant et en rendant accessibles certains habitats qui ne l’étaient pas auparavant (c’est-à-dire en augmentant la quantité de lisières), elle
11 augmente les opportunités pour les prédateurs ou les parasites de couvées d’accéder à ces habitats (Andrén 1995). L’effet de la fragmentation peut varier selon qu’elle est causée par la foresterie ou par l’agriculture (Bayne & Hobson 1997, 2002a), et les lisières agricoles ont souvent plus d’impact que les lisères forestières (Chalfoun et al. 2002). Dans les deux cas, l’augmentation de la quantité de lisières dans le paysage modifie la distribution des individus (Drolet et al. 1999, Villard et al. 1999, Villard et al. 2007), et la présence de zones à découvert conditionne les déplacements des individus à travers la mosaïque forestière (Desrochers & Hannon 1997, Rail et al. 1997, Bélisle et al. 2001, Bélisle & Desrochers 2002, Desrochers et al. 2011). Les lisières peuvent même parfois devenir des pièges écologiques en étant préférentiellement choisies par certains individus (Flaspohler et al. 2001). Cependant, bien qu’elle puisse diminuer le succès au nid (Paton 1994, Manolis et al. 2002), la quantité de lisières peut avoir un effet bénéfique en stimulant l’abondance des insectes (Helle & Muona 1985, Jokimaki et al. 1998, Harris & Reed 2002, Lampila et al. 2005).
La modification des caractéristiques forestières
La fragmentation en forêt boréale s’accompagne généralement d’une modification des caractéristiques forestières car elle est principalement due à la foresterie à grande échelle en Amérique du Nord (Imbeau et al. 2001, Schmiegelow & Mönkkönen 2002). La modification des attributs forestiers tels que l’âge des peuplements peut affecter la composition spécifique (Hobson & Schieck 1999) et la productivité des oiseaux (Rangen et al. 2000), notamment en influençant la composition et l’abondance des communautés de prédateurs (Fisher & Wilkinson 2005, Koprowski 2005). Smith et al. (2010) montrent d’ailleurs que la suppression des prédateurs de nids améliore le succès d’éclosion et d’envol ainsi que la stabilité de la population. Dans la période post-envol, la composition du paysage est déterminante, avec une survie des jeunes améliorée par des structures de végétation plus complexes (King et al. 2006). Finalement, le changement de la structure du paysage peut aussi avoir un impact au niveau évolutif, avec des adaptation morphologiques chez les oiseaux observées en réponse au dernier siècle de modification des paysages forestiers en Amérique du Nord (Desrochers 2010).
12
La gestion et l’aménagement forestiers au Québec
Dans le cadre du nouveau régime forestier du gouvernement du Québec (Ministère des Ressources naturelles 2013a), les initiatives de méthodes de gestion et d’aménagement alternatives aux méthodes traditionnelles d’exploitation sylvicole sont encouragées et prennent une place centrale. Ce nouveau régime se base sur le principe d’aménagement écosystémique qui consiste en « une approche d'aménagement qui vise à maintenir des écosystèmes sains et résilients en misant sur une diminution des écarts entre les paysages naturels et ceux qui sont aménagés afin d'assurer, à long terme, le maintien des multiples fonctions de l'écosystème et, par conséquent, de conserver les bénéfices sociaux et économique que l'on en retire » (Gauthier et al. 2008).
Un des projets pilotes a été mis en place dans la réserve faunique des Laurentides en 2006 et s’est achevé en 2010. Son objectif était de dresser une liste des enjeux de biodiversité dans ce territoire et de proposer des solutions aux problématiques liées aux enjeux. Ce projet a rempli son objectif final qui était de confirmer la faisabilité et la pertinence de l’aménagement écosystémique dans le but de l’appliquer à l’ensemble du Québec. Du point de vue de la faune, on peut relever des enjeux comme la protection des habitats fauniques ou encore des espèces en situation précaire. Concernant les oiseaux en particulier, outre les différents filtres bruts, on peut noter des enjeux particuliers (filtres fins) afin de maintenir certaines espèces comme la Grive de Bicknell (Catharus bicknelli) ou encore le moucherolle à côtés olive (Contopus cooperi), toutes deux présentes dans la zone qui fait l’objet de ce doctorat, la Forêt Montmorency.
La Forêt Montmorency (Figure 1), située dans la réserve faunique des Laurentides, illustre parfaitement les possibilités qui existent en termes de gestion intégrée des ressources et du territoire. Elle est gérée comme une forêt dite « modèle », avec des pratiques sylvicoles effectuées à l’échelle des perturbations naturelles. Elle travaille depuis sa création en 1966 à réduire les impacts négatifs causés par ces interventions. Les perturbations naturelles du domaine bioclimatique de la sapinière à bouleau blanc de l’Est (épidémies d’insectes et chablis) sont remplacées par des interventions contrôlées à l’échelle des perturbations
13 naturelles, et la foresterie pratiquée vise à recréer la mosaïque naturelle d’âge des peuplements (Figure 2). La Forêt Montmorency offre ainsi un cadre idéal pour étudier les effets d’une telle stratégie de gestion et d’aménagement sur les populations vivantes, tout particulièrement les populations d’oiseaux.
Figure 1. Domaines bioclimatiques du Québec méridional et localisation de la Forêt Montmorency.
14
Figure 2. Exemple de mosaïque d’âge des peuplements à l’échelle du paysage à la Forêt Montmorency. Source : André Desrochers.
Objectifs et organisation de la thèse
L’objectif général de ce doctorat est d’évaluer le succès de nidification d’une communauté d’oiseaux en forêt boréale aménagée, et d’étudier certains facteurs qui lui sont associés. J’ai divisé ce doctorat en trois chapitres. Dans mon premier chapitre, je cherche à estimer le succès de nidification d’une communauté d’oiseaux en forêt boréale aménagée à l’aide du transport de nourriture par les adultes comme indicateur de la présence de jeunes au nid. Cette estimation doit tenir compte de la détection imparfaite de ce comportement, les parents ne passant pas tout leur temps à transporter de la nourriture pour leurs jeunes. Afin d’atteindre mon objectif, j’adapte dans ce chapitre des modèles dits d’occupation développés au départ pour étudier le patron d’occupation spatiale (présence/absence) d’espèces d’intérêt (MacKenzie et al. 2002, MacKenzie et al. 2006) mais potentiellement adaptables à d’autres situation binaires telles que des individus nourrissant des jeunes ou non. Mon second chapitre utilise les connaissances acquises dans le premier chapitre ainsi que des données de transport de nourriture relevées depuis 1995 dans la zone d’étude afin d’analyser l’influence de plusieurs facteurs tels que la fragmentation du paysage ou encore
15 les conditions météorologiques sur la phénologie et le succès de nidification des oiseaux en forêt boréale aménagée.
Bien qu’elle donne une information sur le succès de nidification, l’observation du nourrissage des oisillons reste une mesure incomplète de la performance reproductrice. Elle informe sur la présence de jeunes à nourrir mais elle ignore la grande variabilité spatio-temporelle des facteurs qui peuvent affecter le succès reproducteur global tels que la prédation ou encore l’inanition des jeunes au nid. Il apparaît donc essentiel de compléter les observations de transport de nourriture avec une mesure du succès d’envol, mesure plus inclusive du succès reproducteur final. Dans mon troisième chapitre, j’utilise le suivi de groupes familiaux à la fin de la saison de reproduction pour estimer le succès d’envol d’une communauté d’oiseaux de la forêt boréale aménagée. Cette méthode a déjà été utilisée avec succès en forêt boréale (p. ex. Griesser et al. 2007). Cependant, il reste encore beaucoup à faire, notamment en ce qui concerne les déplacements des groupes familiaux et leur détection. J’ai donc développé un protocole d’échantillonnage et adapté des méthodes statistiques particulières (analyse du temps de survie) afin d’estimer la probabilité de détecter un groupe familial et de tenir compte de cette valeur pour estimer le succès d’envol de la communauté aviaire d’intérêt.
17
C
HAPITRE
1
E
STIMATING NESTING SUCCESS
:
A NEW APPROACH
18
Abstract
Indirect methods to estimate nesting success, such as the observation of parental provisioning, usually involve imperfect detection and bias. We developed a method to evaluate parental status and nesting success based on a novel combination of parental provisioning observations and hierarchical modeling. In the summers of 2009 to 2011, we surveyed 392 sites, each on three to four consecutive days at Forêt Montmorency, Québec, Canada. We assessed parental status of 2290 adult songbirds based on parental food provisioning. To account for imperfect detection of parental status, we applied MacKenzie et al.'s (2002) two-state hierarchical model, to obtain unbiased estimated of the proportion of sites with successfully nesting birds, as well as the proportion of adults with broods. To validate estimates of detection probability of parental provisioning, we monitored 16 active nests in 2010 and conducted parental provisioning observations away from known nests. The probability of detecting food provisioning was 31% when using nest monitoring, a value situated within the 11% to 38% range estimated by two-state models. The proportion of adults or sites with broods approached 90% and varied depending on date during the sampling season and year. This study offers a simple and effective sampling design for measuring avian productivity that could be implemented in national surveys such as breeding bird atlases. It also exemplifies the role of Eastern boreal forests as highly productive nesting grounds for songbirds.
Key words: nesting success, imperfect detection, two-state models, songbirds, boreal
19
Résumé
Les méthodes indirectes pour estimer le succès de nidification, telles que l'observation de l’approvisionnement des jeunes par les parents, impliquent généralement une détection imparfaite et un biais. Nous avons développé une méthode pour évaluer le statut parental et le succès de nidification basée sur une nouvelle combinaison d’observations d’approvisionnement parental et de modélisation hiérarchique. Au cours des étés 2009 à 2011, nous avons échantillonné 392 sites, chacun au cours de trois à quatre jours consécutifs à la Forêt Montmorency, Québec, Canada. Nous avons évalué le statut parental de 2290 oiseaux chanteurs adultes basé sur l'approvisionnement alimentaire des parents. Afin de tenir compte de la détection imparfaite du statut parental, nous avons utilisé le modèle hiérarchique à deux états de MacKenzie et al. (2002) de façon à obtenir un estimé non biaisé de la proportion de sites où sont présents des oiseaux élevant des jeunes, ainsi que la proportion d'adultes ayant des couvées. Pour valider les estimations de la probabilité de détection de l’approvisionnement par les parents, nous avons suivi 16 nids actifs en 2010 et mené des observations d’approvisionnement autour des nids connus. La probabilité de détection d'approvisionnement alimentaire était de 31% en utilisant la surveillance des nids, une valeur située dans la plage de 11% à 38% estimée par les modèles à deux états. La proportion d’adultes ou des sites avec couvées approchait 90% et variait selon la date au cours de la saison d'échantillonnage et l'année. Cette étude propose un plan d'échantillonnage simple et efficace pour mesurer la productivité aviaire qui pourrait être mis en œuvre dans les campagnes d’échantillonnage nationales comme les atlas d’oiseaux nicheurs. Elle illustre également le rôle de la forêt boréale de l'Est comme une région de nidification très productive pour les oiseaux chanteurs.
Mots-clés : succès de nidification, détection imparfaite, modèles à deux états, oiseaux
20
I
NTRODUCTIONEstimating reproductive performance of species or a community is important in both long-term monitoring programs and population dynamics studies. Traditionally, nesting success has been measured from nest monitoring (Martin & Geupel 1993), which necessitates the use of cameras or sensors, or that nests be visited frequently by field biologists (Dalley et al. 2009). Nest monitoring delivers a direct and reliable estimation of parameters such as nest survival (Rodewald & Yahner 2001), and the presence of nestlings (Robinson & Robinson 2001) and fledglings (Dalley et al. 2008). Some studies have monitored a large number of nests (Tewksbury et al. 1998, Robinson & Robinson 2001), sometimes for a large sampling area and a high number of bird species (Robinson et al. 1995, Tewksbury et al. 1998). However, nest monitoring is usually restricted to small areas (Martin & Geupel 1993, Sallabanks et al. 2000, Doran et al. 2005) and to one or a small number of target species at a time, generally over small intensively studied plots (Sallabanks et al. 2000). Moreover, despite their labor-intensiveness, nest monitoring studies may be subject to bias in the estimation of nesting success (Ball & Bayne 2012).
Indirect methods can be used to estimate nesting success extensively. They are often based on parental activity, such as food provisioning, to determine the reproductive status of observed adults in the field (Vickery et al. 1992, Villard et al. 1993, Gunn et al. 2000, Rangen et al. 2000, Ibarzabal & Desrochers 2005, Bonifait et al. 2006). However, these indirect methods suffer from imperfect detection, because adult birds reveal their parental status only part of the time. Thus to date, methods that are based on parental provisioning have only yielded negatively biased indices of nesting success (Doran et al. 2005).
MacKenzie et al. (2002) proposed a series of statistical tools to incorporate imperfect detection of dichotomous states such as occupancy, i.e., presence or absence of a target species from sites during the sampling period. They proposed a sampling protocol wherein repeated observations are made at several locations; this temporal replication would provide the data that are required to resolve the ambiguity between species absence and non-detection, when species are not observed. Occupancy models can be applied to other
21 dichotomous states with imperfect detection, such as nesting success, which is defined here as the production of nestlings. In principle, those two-state models can not only help estimate the overall productivity of avian populations, but also determine its variation through space and time, both of which are known to be important (Lack 1966, Newton 1989). Furthermore, two-state models can incorporate variation in the detection of behaviors that are used to infer nesting success. For example, food provisioning is known to vary with time of day (Best 1977, Knapton 1984, Freitag et al. 2001), weather conditions (Best 1977, Johnson & Best 1982, Wittenberger 1982), and prey availability (Pinkowski 1978, Geiser et al. 2008).
Here, we propose and validate a method to estimate parental status of a community of boreal forest songbirds based on a novel combination of repeated measures of parental provisioning and two-state occupancy modeling (MacKenzie et al. 2002). We apply this novel method to a boreal forest landscape, at two different levels: site, and species. We modeled parental status, i.e. the proportion of sites or adults with broods, as a function of date and year, and detection of food provisioning as a function of sampling conditions (weather and time of day). We deliberately limited the number of covariates in our models to provide a simple picture of parental status for the entire songbird community over a forest landscape.
M
ATERIAL AND METHODSStudy area
The study was conducted at the Forêt Montmorency, a research forest managed for timber and which is located 70 km north of Québec City (47°20’N, 71°07’W). The landscape is hilly, with elevations ranging from 600 to 1100 m (Darveau et al. 1997). Mean annual temperature is 0.3°C, and mean annual precipitation is 1589 mm, with 40% falling as snow (Environment Canada 2013). During the three summers of this study, mean ambient air temperature in Forêt Montmorency was 13.5C, which is similar to the long-term average
22
(Environment Canada 2013). Between 2009 and 2011, precipitation was unusually scarce. In particular, 2010 was especially dry, with total June and July rainfall measuring 70.6 mm and 113.5 mm respectively (mean 92.1 mm), compared to 136.6 mm between 1981 and 2010 (Environment Canada 2013).
Coniferous forests covered 52% of the harvestable territory and were mostly mature (61- to 80-years-old). Mixed forests represented 34% of total cover and were mostly young (21- to 60-years-old). The remaining area had no forest cover and corresponded mainly to clearcuts, windthrows and spruce budworm outbreaks (Laberge & Blanchette 2010). Mature forest stands were dominated by balsam fir Abies balsamea (L.) Miller, accompanied by black spruce Picea mariana, white or paper birch Betula papyrifera, and white spruce Picea glauca (Beauchesne 1991).
Sampling parental provisioning
We surveyed 224 point locations (called “sites”) that were located randomly on unpaved forestry roads and trails (Figure 3a), and 168 sites that were located on systematic transects dispersed over the entire study area (Figure 3b). We covered the entire range of forest succession stages (1 to > 100 years-old) using a combination of points placed randomly along forestry roads (minimum distance 200 m) and systematically along linear transects into the forest (distance 250 m). In 2009, three visits (sampling occasions) were conducted at each site, whereas in 2010 and 2011 we made four. Sampling was done from 05h00 to 10h00 EDT, between 28 May and 21 July each year, during days with no or little rain and no strong wind (< 3 on Beaufort scale) and, as often as possible, on consecutive days (43% of the time; otherwise, between 5 and 7 days to achieve the three or four visits at the same site). Two observers rotated every other day to survey sites. We visited each site only during one of the three years of study. On each sampling occasion, we used a playback of Black-capped Chickadee (Poecile atricapillus) mobbing calls to draw birds in, facilitate sighting and in turn, the detection of parental status (food provisioning: Gunn et al. 2000). We assume that birds responded to mobbing calls irrespective of nesting status because mobbing calls locally elicit strong responses even outside the nesting season (e.g. Bélisle &
23 Desrochers 2002, Desrochers et al. 2002), and the number of birds attracted to mobbing calls at a given site did not vary according to date in the sampling period, despite strong differences in the proportion of birds provisioning their young (unpublished data). We played the mobbing calls with a five-watt Pignose amplifier connected to digital audio player for a continuous duration of 15 min on each sampling occasion.
Figure 3. Sampling site locations on a) roads and trails, and b) systematic transects. Blue lines and areas are rivers and lakes, grey lines, unpaved forestry roads and trails.
Application of two-state models to parental status
We used MacKenzie et al.’s (2002, 2006) likelihood-based method with two states, which was developed for estimating the proportion of sites occupied by individuals or species when detection is imperfect. Here, we were interested in evaluating parental status. We estimated i, the probability of presence of nestlings at site i, and pi, the conditional
24
probability that parental status is detected at site i at a given time, given the presence of young.
We considered parental status at two different levels. First, we estimated the proportion of sites where there was at least one adult provisioning young, given occupancy by at least one species. Thus, for each visit at each site, we considered two outcomes, i.e. at least one adult carrying food, all species confounded, or no adults carrying food. Secondly, we considered the proportion of pairs with broods at the species level, given occupancy. At the latter level, a food provisioning detection history (at least one adult carrying food vs. not) was initiated for each species seen at a given site. If no individual of the species was seen at one visit, a missing data point was recorded to avoid confounding imperfect detection of parental status and imperfect detection of the birds themselves. None of the individual birds were seen on four sampling occasions; thus, we randomly deleted one of the missing data from each history of visual capture-recaptures in 2010 and 2011 and, therefore, worked with visual capture histories of three sampling occasions for this dataset.
We implemented two-state models with the Unmarked package 0.9-7 in R Version 2.15.3 (Ihaka & Gentleman 1996, Fiske & Chandler 2011, R Development Core Team 2013). This package enabled us to estimate the proportion of adults with broods, as well as the detection probability of food provisioning, given parenthood, as a function of several covariates. We were interested in a general assessment of parental status; thus, we kept the number of covariates to a minimum. We modeled the effects of Julian day (mean day of the visits to a given site), year and the interaction between these two regressors. Julian day-squared was included in an initial set of models, but its effect never approached significance. Thus, this covariate was ignored in the models presented here. Sampling time of day and meteorological conditions, i.e. hourly ambient temperature (C) and hourly rainfall (mm), were modeled because of their potential influence on parental provisioning. Weather data were obtained from the weather station < 1 km from the validation study area (Environment Canada 2013). With the null model and the full one (containing all covariates), we compared six models for each studied level of parental status (R code available in
25 Appendix
A
), based on the Akaike information criterion (AIC) (Burnham & Anderson 2002). The goodness-of-fit of the best model was tested with 1000 parametric bootstraps (Fiske & Chandler 2011).Validation of two-state models
Models and analytic tools used here are an extension of traditional closed-population capture-recapture theory (Otis et al. 1978, White et al. 1982). In the case of occupancy modeling, several assumptions need to be met; those assumptions can be adapted to the study of parental status and nesting success (Table 1). To avoid parental status change and, therefore, assume closure of parental status during the sampling period, we conducted visits during consecutive days as often as possible, i.e. 43% of the time (otherwise, between 5 and 7 days to achieve the three or four visits at the same site). Given that the entire nesting cycle lasts ~ 26 days from egg-laying to fledging in the species studied (Gauthier & Aubry 1996), not to mention days or weeks of fledgling care, changes in nesting state that were due to hatching in the course of a visual detection history must have occurred only rarely. However, changes in parental status due to nest failure in the course of a visual capture history may occur more frequently, thereby compromising the closure assumption. To address this concern, we compared the frequency of cases where parental status was detected initially but not on later occasions ("1-0" sequences in visual capture histories), to the converse cases ("0-1" sequences). We would interpret a greater frequency of "1-0" sequences as evidence of lack of closure due to nest failure.