Fonction et localisation de la PDE8A dans les cellules
ovariennes porcines et son implication dans la
stéroïdogenèse
Mémoire
Amel Lounas
Maîtrise en sciences animales
Maître ès sciences (M. Sc.)
Québec, Canada
Fonction et localisation de la PDE8A dans les cellules
ovariennes porcines et son implication dans la
stéroïdogenèse
Mémoire
Amel Lounas
Sous la direction de :
III
Résumé
Les nucléotides cycliques sont des seconds messagers intracellulaires possédant une grande importance dans la signalisation cellulaire du follicule ovarien. Les niveaux intracellulaires en nucléotides cycliques tel que l’adénosine monophosphate cyclique (AMPc) dépendent de leur synthèse, assurée par l’adenylyl-cyclase (AC) ainsi que leur dégradation par les phosphodiéstérases (PDE). Ces dernières appartiennent à la superfamille des métalophosphohydrolases, elles hydrolysent le groupement phosphate en 3’ des nucléotides cycliques pour produire un nucléotide 5’ phosphate. Dans les cellules ovariennes, plusieurs familles de PDE ont été identifiées, agissant comme modulateurs des taux intracellulaires de nucléotides cycliques.
Dans le présent projet, nous nous attarderons à étudier la signalisation cellulaire chez les cellules ovariennes impliquant l'AMPc. La régulation de ses concentrations intracellulaires affecte plusieurs processus physiologiques. Le projet s'intéresse particulièrement à une famille d’enzyme de dégradation de l'AMPc, la phosphodiestérase8A (PDE8A). Nous voulons donc valider la présence fonctionnelle de la famille de PDE8A dans les cellules ovariennes ainsi que la mitochondrie afin de comprendre l’implication de cette enzyme dans la physiologie cellulaire en s’attardant à la stéroïdogenèse, étant donné que les mitochondries sont un organite cellulaire essentiel pour la stéroïdogenèse parce qu’elles représentent le site de synthèses de plusieurs hormones stéroïdiennes. En effet, des récents travaux ont montré la famille PDE8 comme un régulateur de la stéroïdogenèse dans les cellules de Leydig.
Dans cette étude, nous avons montré que le transcrit de PDE8A ainsi que sa protéine étaient exprimés dans les cellules de la granulosa, les cellules du cumulus et l'ovocyte chez le porc. Ainsi, la protéine PDE8a a été détectée par western blot dans des mitochondries isolées à partir des cellules de granulosa et des complexe ovocyte-cumulus (COC). Une co-localisation entre le signal immunoréactive de la PDE8A et le marquage réalisé par mitotracker a été observée dans les cellules de granulosa et des mitochondries isolées. De plus, la présence fonctionnelle de la PDE8 mesurée en tant qu'activité AMPc-PDE sensible au PF-04957325 a été détectée dans des cellules de la granulosa et des mitochondries isolées supportant la présence fonctionnelle de la PDE8A dans les mitochondries isolées. De ce fait, cette observation soutient encore la localisation fonctionnelle mitochondriale de la PDE8A. Une association entre la mitochondrie et la PDE8A a aussi été
IV
démontrée par immunomicroscopie électronique qui a confirmé l’ancrage de la protéine sur la membrane mitochondriale externe.
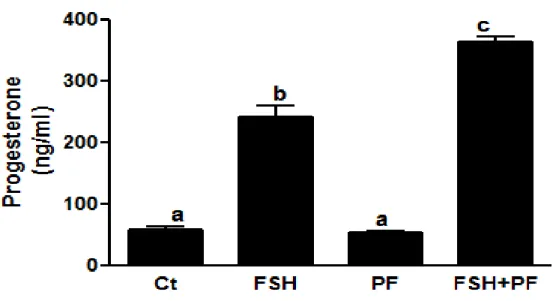
Pour évaluer l’implication de la mitochondrie dans la stéroïdogenèse, l'effet de PDE8A sur la production de progestérone a été mesuré dans les complexes ovocyte-cumulus en utilisant le PF-04957325 comme un inhibiteur spécifique de PDE8. Lorsque les COC sont cultivés sans FSH, un niveau basal de progestérone est mesuré. Par contre en présence de FSH une augmentation significative de sécrétion de progestérone est observée. En utilisant un inhibiteur spécifique de la PDE8 (PF-04957325) avec la FSH nous avons obtenu une augmentation encore plus importante de la sécrétion de progestérone.
Ces résultats démontrent la présence fonctionnelle de PDE8A dans les cellules de la granulosa et les complexes ovocyte-cumulus. Puisque les mitochondries sont l'une des localisations de PDE8A, l'effet de l'inhibiteur spécifique de PDE8 sur la sécrétion de progestérone soutient la contribution mitochondriale de PDE8 dans la stéroïdogenèse.
V
Table des matières
Résumé ... III Table des matières ... V Liste des tableaux ... VII Liste des figures ... VIII Liste des abréviations... IX Remerciements ... X AVANT-PROPOS ... XI
Chapitre 1 : Introduction ... 1
1.1. Structure et physiologie ovarienne ... 2
1.1.1. L’ovocyte ... 3
1.1.2. Les cellules du cumulus ... 4
1.1.3. Les cellules de la granulosa ... 4
1.1.4. Les cellules de la thèque ... 5
1.1.5. Stéroïdogenèse ... 6
1.1.6. Folliculogenèse ... 8
1.1.7. Atrésie ... 11
1.1.8. Ovulation ... 12
1.1.9. Lutéinisation ... 13
1.2. Maturation du complexe ovocyte-cumulus ... 14
1.2.1. Maturation de l’ovocyte ... 14
1.2.2. Expansion du cumulus ... 14
1.2.3. Régulation de la reprise méiotique ... 15
1.2.4. Mécanisme de reprise de la méiose ... 15
1.3. Phosphodiéstérases ... 16
1.3.1. Classification ... 16
1.3.2. Distribution des phosphodiéstérases dans l’ovaire ... 22
1.4. Compartimentalisation de l’AMPc dans la mitochondrie ... 22
1.4.1. La mitochondrie ... 23
1.5. Hypothèse et objectifs de travail ... 29
1.6. Bibliographie ... 30
Chapitre 2 : cAMP-specific phosphodiesterase 8A is localized in mitochondria of ovarian cells and regulates progesterone synthesis ... 43
VI
2.1. Résumé ... 44
2.2. Abstract ... 45
2.3. Introduction ... 47
2.4. Materials and methods ... 49
2.4.1. Chemicals ... 49
2.4.2. Ovary collection and tissue recovery ... 49
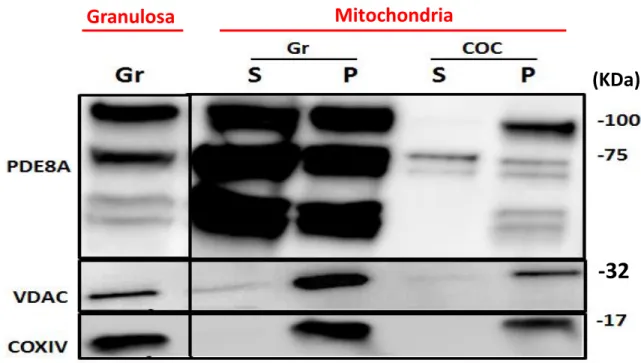
2.4.3. Isolation of mitochondria ... 49 2.4.4. RT-PCR ... 50 2.4.5. Western Blotting ... 50 2.4.6. Immunohistochemistry ... 51 2.4.7. Immunofluorescence ... 51 2.4.8. ImmunoElectron Microscopy ... 52 2.4.9. PDE assay ... 53 2.4.10. Progesterone quantification... 53 2.5. Results ... 54
2.5.1. PDE8A expression in ovarian follicular cells ... 54
2.5.2. PDE8A proteins are present in ovarian follicle ... 54
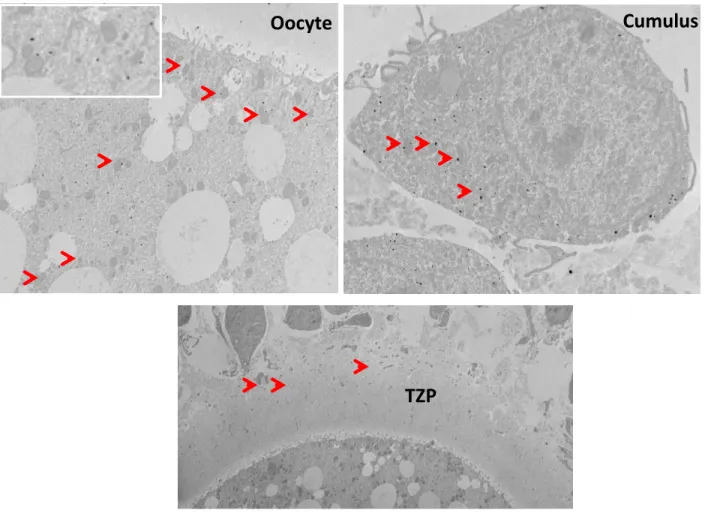
2.5. 3. PDE8A immunolocalisation ... 55
2.5.4. PDE8A and Mitochondria ... 56
2.5. 5. Effect of PDE8 inhibition on progesterone secretion during IVM ... 60
2.6. Discussion ... 62
2.7. Conclusion ... 66
2.8. Acknowledgments ... 66
2.9. References ... 67
VII
Liste des tableaux
Tableau 1 : Primers used to performed PCR amplifications ... 50 Tableau 2 : Immunoelectron microscopy localisation of PDE8A in cumulus-oocyte complex ... 60
VIII
Liste des figures
Figure 1.1 : Représentation schématique d’un follicule ovarien au stade antral (Avec obtention des droits
d’auteurs) ... 2
Figure 1.2 : La stéroïdogenèse ovarienne et la théorie «2 cellules- 2 gonadotrophines » adaptée ... 8
(Avec obtention des droits d’auteurs) ... 8
Figure 1.3 : Les principales étapes du développement d’un follicule ovarien ... 9
(Avec obtention des droits d’auteurs) ... 9
Figure 1.4 : Régulation du développement folliculaire par les deux facteurs FSH et GDF9 ... 12
(Avec obtention des droits d’auteurs) ... 12
Figure 1.5 : Structure comparative des PDE (Avec obtention des droits d’auteurs) ... 17
Figure 1.6 : Structure des 11 familles de PDE (Avec obtention des droits d’auteurs) ... 21
Figure 2.1 : PDE8A transcript detection by RT-PCR in ovarian cells ... 54
Figure 2.2 : Western blot of PDE8A in follicule cells ... 55
Figure 2.3 : Immunolocalisation of PDE8A in Cumulus-oocyte complexes ... 55
Figure 2.4 : Western blot detection of PDE8A in isolated mitochondria ... 57
Figure 2.5 : Immunofluorescence detection of PDE8A in granulosa cells and isolated mitochondria ... 58
Figure 2.6 : Immunoelectron microscopy localisation of PDE8A in cumulus-oocyte complex ... 59
Figure 2-7 : Effect of PDE8A inhibition on progesterone synthesis in cumulus-oocyte complexes during maturation ... 61
IX
Liste des abréviations
5’-AMP 5’-Adénosine monophosphate 5’-GMP 5’-Guanosine monophosphate AC Adénylyl cyclase
AMH Hormone anti-mullérienne
AMPc Adénosine monophosphate 3’5’-cyclique ARNm ARN messager
BMP15 Protéine morphogénétique osseuse 15 Ca2+ Ion calcium
CDK1 Kinase cycline-dépendante 1 Cx37 Connexine 37
Cx43 Connexine 43
CYP11A Cytochrome P450 coupant la chaine latérale CYP17 Cytochrome P450 17α-hydroxylase/C17–20 lyase CYP19A1 Cytochrome P450 aromatase 19A1
DAO Dérivés actifs de l’oxygène EGF Facteur de croissance épidermal FGF Facteur de croissance des fibroblastes FSH Hormone folliculo-stimulante
GDF9 Facteur de croissance et de différentiation 9 GMPc Guanosine monophosphate 3’5’-cyclique IBMX 3-Isobutyl-methyl xanthine
IGF-I Facteur de croissance semblable à l’insuline I IGF-II Facteur de croissance semblable à l’insuline II LIF Leukemia inhibiting factor
LH Hormone lutéinisante
MPF Facteur de promotion de la phase-M PDE Phosphodiestérase
PKA Protéine kinase dépendante de l’AMPc PI3K Phosphoinositol 3-kinase
PKB Protéine kinase B PKC Protéine kinase C PGF2α Prostaglandine F2α
TNF Facteur de nécrose tumorale
TOM Translocase de la membrane externe VDAC Canal anion-sélectif dépendant de la tension
X
Remerciements
J’aimerais remercier en premier lieu mon directeur de recherche François Richard de m’avoir accueilli dans son laboratoire, je le remercie aussi pour son orientation, ses encouragements, ses conseils et surtout sa présence durant tout mon projet de maitrise.
Je remercie les deux professionnels de son équipe Christine Guillemette et Alexandre Bastien pour leur aide sans oublier notre chère technicienne des sciences animales Isabelle Laflamme.
Je tiens à remercier les étudiants de notre département ainsi que ceux du Centre de recherche en reproduction, développement et santé intergénérationnelle (CRDSI) pour toutes les discussions innovantes.
Finalement, je voudrais remercier ma grande famille surtout "ma mère" qui est d’une importance majeure dans ma vie, mes frères, ma sœur et spécialement mon mari.
XI
AVANT-PROPOS
Ce mémoire contient un chapitre rédigé en anglais sous forme d’un article scientifique dont je suis l’auteur prinicipal. L’intitulé de l’article inséré est -cAMP-specific phosphodiesterase 8A is localized in mitochondria of ovarian cells and regulates progesterone synthesis-. Les coauteurs sont les chercheurs François J. Richard, Marie-EveTremblay ainsi que leurs professionnelles de recherche Christine Guillemette et Nathalie Vernoux. L’article sera soumis pour publication dans la revue Scientific Reports.
1
2
1.1. Structure et physiologie ovarienne
L’appareil reproducteur femelle est constitué principalement par deux ovaires représentant les gonades femelles, où sont produits les ovules. L’unité de base de l’ovaire est le follicule ovarien qui est formé avant la naissance. Ce dernier, soutient les fonctions physiologiques principales de l’ovaire tel que la production d’hormones sexuelles et la croissance de l’ovocyte qui aboutit à l’ovulation une fois que le follicule est mature. Après l’ovulation, le follicule ovarien se transforme en corps jaune qui joue un rôle durant la gestation (1).
Comme le montre la figure 1.1, chaque follicule ovarien comprend un ovocyte entouré par des cellules somatiques permettant la croissance et la maturation d’un ovocyte. On retrouve de l’intérieur vers l’extérieur les cellules du cumulus et les cellules du granulosa qui forment un épithélium interne pseudo stratifié. Ce dernier possède une fonction stéroïdogénique importante. La dernière couche de ces cellules somatiques est représentée par les cellules de la thèque qui forment un tissu stromal avec une fonction stéroïdogénique différente de celle des cellules de granulosa. L’ensemble de ces cellules somatiques soutient la croissance de l’ovocyte et facilite l’ovulation (2).
Figure 1.1 : Représentation schématique d’un follicule ovarien au stade antral (Avec obtention des droits d’auteurs) (3)
3
1.1.1. L’ovocyte
L’ovocyte, la cellule germinale femelle, est localisé dans une niche appelée le follicule ovarien. Il est considéré comme la plus grosse cellule de l’organisme (100 µm de diamètre), il apporte son génome haploïde ainsi que la quasi-totalité de son cytoplasme pour contribuer au développement embryonnaire, contrairement au spermatozoïde qui ne participe qu’avec son génome. De ce fait, l’ovocyte est capable d’assurer la croissance précoce de l’embryon et cela selon son état de maturation (4). Cette maturation est indispensable pour le développement embryonnaire parce qu’elle permet au gamète femelle d’acquérir les compétences requises pour être fécondé, se développer en embryon et enfin de donner naissance à un nouvel individu sain (5).
L’ovogénèse est un processus qui débute dès la vie embryonnaire, et reste actif jusqu'à la fin de la vie reproductive de la femelle. Durant ce processus, le gamète femelle passe par différentes étapes de croissance, différenciation et maturation et cela au sein du follicule ovarien (6).
Pendant la vie fœtale, les cellules germinales primordiales qui colonisent les gonades embryonnaires donnent naissance à des ovogonies. Ces dernières, prolifèrent dans les cordons ovigères et entrent en méiose. Ce passage d’ovogonies en prophase de la première division méiotique est marqué par leur transition en ovocytes et un arrêt de prolifération (7). À ce stade, les ovocytes s’entourent d’une unique couche de cellules somatiques aplaties, les cellules de granulosa et d’une membrane basale, pour former les follicules primordiaux. Ces derniers représentent une réserve ovarienne d’ovocytes qui s’épuise au cours de la vie de la femelle. La folliculogenèse est le processus comprenant le développement et la croissance des follicules primordiaux jusqu’à l’ovulation, ce qui permet la libération d’un ovocyte fécondable (8). La folliculogenèse et l’ovulation sont deux mécanismes hautement régulés par l’axe hypothalamo-hypophysaire. Ce dernier contrôle la sécrétion des gonadotrophines FSH et LH suite aux différents stimuli endogènes et exogènes. On note aussi le rôle crucial de l’hormone anti-mullérienne (AMH), sécrétée par les cellules de granulosa des follicules en croissance, dans la gestion et la régulation du recrutement des follicules pour éviter l’épuisement rapide de la réserve ovarienne d’ovocytes (9).
Durant la folliculogenèse, l’ovocyte subit la maturation et prépare son cytoplasme afin d’acquérir les compétences requises pour la reprise de la méiose. Cet ovocyte se trouve dans un follicule dominant destiné à l’ovulation sous l’effet des gonadotrophines. Une fois que la méiose est reprise, elle sera
4
poursuivie jusqu’au stade de métaphase de la deuxième division méiotique, stade auquel l’ovocyte sera de nouveau bloqué (10).
1.1.2. Les cellules du cumulus
Une fois au stade antral, une sous population des cellules de la granulosa se différencie en cellules du cumulus. Ce sont les cellules qui entourent l’ovocyte et établissent avec lui une étroite communication par des jonctions perméables (gap junction). Les molécules de petits tailles ( 1KDa) passent à travers ce réseau de jonctions pour participer à la transmission des signaux intracellulaires au sein du complexe ovocyte-cumulus.
Les cellules du cumulus régulent et contribuent au processus de maturation de l'ovule. En effet, la prolifération et la différenciation des cellules du cumulus sont contrôlées en grande partie par l'ovocyte et cela par la sécrétion de certaines protéines, telles que GDF9 (facteur de croissance et de différenciation 9) et la BMP 15 (protéine morphogénétique) (11).
Les cellules de cumulus se caractérisent par une haute capacité proliférative et une faible activité stéroïdogénique mais leur faible expression du récepteur LH reste la caractéristique principale par comparaison avec les cellules de la granulosa. Une expansion importante des cellules de cumulus est caractérisée par leur prolifération qui est sous l’influence des gonadotrophines ovulatoires. Ces dernières, interviennent dans la stimulation des cellules de cumulus pour sécréter une matrice extracellulaire, qui est composée entre autres d’acide hyaluronique (12, 13, 14). L’expansion du cumulus est très importante pour la fertilité, ce processus est considéré comme une empreinte moléculaire qui peut aider à prédire la qualité de l’ovocyte (15).
1.1.3. Les cellules de la granulosa
La croissance folliculaire passe par trois stades : primaire, secondaire et tertiaire. Les changements morphologiques qui surviennent lors de cette croissance sont marqués par l’installation des cellules de granulosa et cela dès le stade follicules primaire. Une seule couche de cellules de granulosa de structure aplatie entoure l’ovocyte bloquée à la prophase I dans un follicule primordial au repos. Mais une fois que la croissance de ce dernier est activée, les cellules de granulosa adoptent une forme cubique. En revanche, plus de deux couches de cellules de granulosa entourent l’ovocyte dans un follicule antral. De ce fait, le stade de croissance des follicules ovariens peut être déduit à partir du
5
nombre de couches de cellules de granulosa (16). Les modifications structurales de cellules des granulosa sont accompagnées par l’acquisition de spécificités fonctionnelles par lesquelles, elles agissent sur les fonctions physiologiques de l’ovaire. Durant ces stades de la croissance de follicules ovariens les cellules de granulosa développent un réseau de communication avec les cellules adjacentes et avec l’ovocyte grâce à l’expression de la connexine 43 (Cx43) et la connexine 47 (Cx47) (17). Le rôle de ces deux dernières dans le développement folliculaire et ovocytaire a été mis en évidence par l’inactivation des gènes correspondant à chacune des deux familles chez la souris. L’altération de la croissance du follicule ovarien ainsi que l’ovocyte chez ces souris knock-out a été observée, ce qui confirme l’importance de la communication intercellulaire dans la physiologie du follicule ovarien (18, 19, 20).
1.1.4. Les cellules de la thèque
Les cellules de la thèque se divisent en deux couches cellulaires : la thèque externe et la thèque interne, toutes les deux sont des cellules de type stromal. Les cellules de la thèque externe sont compactes et moins vascularisées, elles ne sont pas douées d’une fonction sécrétoire tandis que la thèque interne est caractérisée par une haute activité stéroïdogénique. De ce fait, elle est pourvue d’une grande vascularisation lui permettant d’assurer sa fonction sécrétrice des stéroïdes (progestérone, androgènes) lors de processus de la folliculogenèse (21). Durant la croissance folliculaire, les cellules de la thèque sont stimulées par la LH contrairement aux cellules de la granulosa qui sont contrôlées par la FSH puis la LH (22).
Les cellules de la thèque s’installent autour du follicule ovarien dès les premiers stades de développement folliculaire (follicules primordiaux et primaires), elles proviennent de la différenciation des cellules mésenchymateuses qui délimitent le follicule primordial (23). De ce fait, les cellules de la thèque sont d’une grande importance dans la croissance et la différenciation des cellules folliculaires durant toute la folliculogenèse. Durant la croissance folliculaire certains facteurs sécrétés par les cellules de la granulosa comme la FSH, IGF-I et II et GDF-9 sont essentiels pour la différenciation des cellules de la thèque ainsi que dans la régulation de leur activité (24).
Au stade de follicule secondaire, les cellules de la thèque contribuent aussi dans le développement ovocytaire par l’établissement d’une vascularisation autour de l’ovocyte. Plusieurs signes marquent la maturité folliculaire telle que la multiplication et la différenciation des cellules de la thèque ainsi que
6
l’acquisition de l’activité stéroïdogénique qui est associée à la distribution des récepteurs à la LH à leur surface. Cette stéroïdogenèse est considérée comme la principale fonction des cellules thécales qui assurent la production d’androgènes à partir du cholestérol une fois qu’elles sont stimulées par la LH (25, 26). Par la suite, ces androgènes seront utilisés par les cellules de la granulosa afin de synthétiser les œstrogènes (27). Cette biosynthèse nécessite le recrutement de certaines protéines clés synthétisées par les cellules de la granulosa et les cellules de la thèque. Ces dernières, expriment plusieurs enzymes qui contribuent activement à la stéroïdogenèse, telle que la CYP11A qui est responsable de la conversion du cholestérol importé dans la mitochondrie en prégnenolone (28). Par la suite la CYP17, une autre enzyme thécale intervient pour transformer la prégnenolone en androstènedione qui sera utilisée par les cellules de la granulosa afin de produire l’œstrogène (29). L’activité stéroïdogénique des cellules thécales est maintenue jusqu’ à l’ovulation, après ce stade les cellules thécales subissent des changements morphologiques et fonctionnels lui permettant de participer à la formation du corps jaune en association avec les cellules de la granulosa. Cette nouvelle entité cellulaire est chargée de produire la progestérone qui est une hormone essentielle pour la gestation (30).
1.1.5. Stéroïdogenèse
La stéroïdogenèse est une caractéristique principale des cellules de la granulosa. Elle se résume dans la synthèse des estrogènes à base d’androgènes déjà produits par les cellules de la thèque interne. Ce processus de biosynthèse nécessite l’intervention de deux gonadotrophines « FSH et LH ». La première hormone, contrôle l’aromatisation des androgènes en agissant sur les cellules de la granulosa qui expriment des récepteurs à la FSH, tandis que la deuxième gonadotrophine, stimule les cellules de la thèque interne qui possèdent des récepteurs à la LH, pour produire les androgènes (l’androstènedione et la testostérone). C’est la théorie «2 cellules- 2 gonadotrophines » (31). Au début de la croissance folliculaire, les cellules de la granulosa marquent une faible expression des récepteurs à la FSH, qui seront de plus en plus abondants vers la dernière phase de développement folliculaire. Par la suite, quand le follicule ovarien atteint sa dominance, les cellules de la granulosa auront la capacité d’exprimer les récepteurs à la LH (32). Toujours au cours de cette dernière phase de croissance folliculaire, les cellules de la granulosa sont aussi capables de synthétiser une enzyme appartenant à la famille de cytochrome P450, (CYP19A1). Par un mécanisme d’hydroxylation, cette aromatase contribue dans la conversion d’androgènes en œstrogènes chez les cellules de la
7
granulosa. L’œstradiol 17β est obtenu à partir de la testostérone, par contre l’estrone est synthétisé à partir d’androstènedione (33). Deux autres enzymes appartenant aussi à la famille de cytochrome P450 sont synthétisées par les cellules de la thèque interne et cela dès leur formation au cours des premiers stades de développement folliculaire (34). Ces enzymes sont CYP11A1 et CYP17A1 qui ont un rôle majeur dans la synthèse des progestagènes et des androgènes. La première enzyme assure la conversion du cholestérol en prégnénolone (35). La deuxième par contre est responsable de la synthèse de la progestérone à partir de la prégnénolone (36). Plusieurs facteurs endocriniens peuvent contribuer dans la régulation de ces voies de synthèse en agissant sur les cellules folliculaires ainsi que les hormones clés de la stéroïdogenèse. L’IGF-I et –II (facteurs de croissance ressemblant à l’insuline), sont des principaux modulateurs des effets des deux gonadotrophines « FSH et LH ». Ils régulent l’action de ces deux hormones sur les cellules cibles « cellules de granulosa et cellules de la thèque ». Cette voie de régulation affecte l’activité stéroïdienne de ces deux cellules folliculaires (37). Un autre système de régulation est exercé par deux autres peptides l’inhibine et l’activine. La production d’androgènes par les cellules de la thèque est régulée positivement par l’inhibine A qui stimule la synthèse d’androgènes en présence de la LH pour une production optimale d’estradiol (38). Tandis que le rétrocontrôle négatif est assuré par l’activine qui agit pour inhiber la production d’androgènes dans les cellules de la thèque (39, 40). À son tour, l’activité aromatase exercée par les cellules de la granulosa pourrait être une cible de régulation pour l’EGF (facteur de croissance épidermique). Ce dernier possède un effet mitogène sur les cellules de la granulosa et peut aussi inhiber leur activité aromatase (41).
8
1.1.6. Folliculogenèse
Le stock de follicules primordiaux passe par deux grandes phases de développement folliculaire, la folliculogenèse basale et terminale, durant lesquelles se déroule la croissance folliculaire. Différents stades sont marqués durant cette croissance, le stade de follicule primordial, le stade primaire et secondaire, puis le stade des follicules antraux. Ces transitions sont caractérisées par des aspects morphologiques spécifiques à chaque stade de croissance folliculaire. La croissance de l’ovocyte contenu dans le follicule en croissance accompagne ces transitions ainsi que la multiplication et la différenciation des cellules folliculaires pour la mise en place de plusieurs compartiments cellulaires telle que des cellules de la granulosa et les cellules de la thèque. Ces phases de croissance aboutissent à l’ovulation pour certains follicules ou à l’atrésie pour la plupart des autres (43).
Les facteurs qui déclenchent et régulent les premiers stades de ce processus de folliculogenèse restent inconnus. Au cours de la première phase de la folliculogenèse, les cellules du follicule ovarien
(42)
Figure 1.2 : La stéroïdogenèse ovarienne et la théorie «2 cellules- 2 gonadotrophines » adaptée (Avec obtention des droits d’auteurs)
9
« cellules de granulosa » expriment peu de récepteurs à la FSH. De ce fait, la folliculogenèse basale est indépendante des taux de la FSH vu que cette dernière présente un faible effet stimulateur sur les cellules cibles. En revanche, la folliculogenèse terminale est hautement dépendante des deux gonadotrophines, FSH et LH qui possèdent un rôle majeur dans la croissance et la différenciation folliculaire (44). En plus de ces deux hormones, certains facteurs stimulateurs ont une véritable contribution au cours de développement folliculaire. La protéine BMP15 et le facteur GDF-9 sont sécrétés par l’ovocyte dès les premiers stades de développement folliculaire pour réguler la progression et le déroulement de la folliculogenèse basale ainsi que la stimulation de la prolifération et la survie des cellules de la granulosa (45). Les facteurs de croissance des fibroblastes (FGF 2 et 7) sont aussi exprimés par l’ovocyte, ils sont impliqués dans la survie et la croissance folliculaire ainsi que la formation de l’antre au stade follicule tertiaire (46). La participation de « Leukemia Inhibiting Factor (LIF) » ainsi que « Stem Cell Factor » semble aussi essentielle pour l’initiation et la progression du développement folliculaire au cours de la folliculogenèse (47).
Figure 1.3 : Les principales étapes du développement d’un follicule ovarien (Avec obtention des droits d’auteurs) (48)
10
1.1.6.1. Recrutement
La prochaine étape de la folliculogenèse est le recrutement des follicules ovariens qui ont subi un développement folliculaire et qui ont atteint une taille de 0,2 mm chez la souris, 2 mm chez les primates, 3 mm chez le porc et 4 mm chez la vache. Ces follicules sont recrutés pour accomplir leurs croissances et se rendre à l’ovulation. De plus, ces follicules expriment des récepteurs aux gonadotrophines et dépendent de la présence de ces hormones pour la poursuite de la croissance (49). La FSH est considérée comme l’hormone clé du mécanisme de recrutement, son niveau plasmatique augmente graduellement durant le recrutement folliculaire. C’est à ce stade que les cellules de la granulosa répondent à la stimulation par la FSH et acquièrent la capacité à aromatiser les androgènes en œstrogènes. Les follicules recrutés doivent être suffisamment sensibles à la FSH pour passer à une autre phase de croissance (50).
1.1.6.2. Sélection
Selon l’espèce, un ou certains follicules recrutés réussissent à franchir l’étape de sélection pour devenir des follicules prè-ovulatoires alors que les autres follicules du groupe subissent l’atrésie (51). Le mécanisme de sélection est manifesté par une chute des taux de sécrétion de la FSH plasmatique ainsi qu’une augmentation de la production d’œstradiol. Ce dernier est en association avec l’inhibine exercent une régulation négative sur la synthèse de la FSH par l’axe hypothalamo-hypophysaire. La grande diminution de la concentration en FSH empêche le maintient en vie de la totalité des follicules recrutés. Cependant, selon l’espèce un ou plusieurs follicules recrutés vont être sélectionnés (12-20 pour le porc, 1 seul pour la vache) afin de poursuivre la croissance et devenir des follicules ovulatoires (52).
1.1.6.3. Dominance
Un ou plusieurs des follicules ovariens préalablement sélectionnés (selon l’espèce) ont acquis la capacité de compléter leur croissance folliculaire et devenir un follicule dominant. Durant cette phase, les cellules de la granulosa de follicules sélectionnés expriment des récepteurs à la LH. En association avec l’augmentation de la pulsatilité de la LH, le follicule sélectionné est capable de poursuivre le développement folliculaire et devenir un follicule dominant malgré les faibles taux de la
11
FSH. De ce fait, la LH est indispensable pour maintenir la survie du follicule dominant qui est capable de provoquer la régression puis la dégénérescence des follicules en croissance (53).
1.1.7. Atrésie
Lors de processus de sélection folliculaire, la majorité des follicules ovariens subissent une involution traduite par la diminution de leur taille folliculaire jusqu`à la disparition totale. Cette régression folliculaire ou atrésie est un mécanisme physiologique normal permettant d`éliminer certains follicules qui ne sont pas encore au bon stade de développement folliculaire. Ainsi, les follicules les plus disposés à être éliminés par atrésie sont ceux qui ne sont pas capables de répondre à la LH pour compléter leur croissance en raison d’absence des récepteurs de cette gonadotrophine sur les cellules de la granulosa (54).
Dans le follicule ovarien, les cellules de la granulosa sont les premières cellules folliculaires ciblées par l`atrésie, cette dernière va déclencher un mécanisme d`apoptose au sein de ces cellules afin d’assurer leur dégénérescence (55, 56, 57). Tout d`abord, l`atrésie induit un arrêt de la prolifération des cellules de la granulosa, par la suite elle provoque une disparition de l’activité aromatase associée à une diminution de la production d’œstradiol et d’inhibine au sein de ces cellules (58). Par conséquence, les cellules de la granulosa subissent une dégénérescence cellulaire par un processus apoptotique qui touche à ce stade les cellules thécales (59). Ces dernières présentent par contre une diminution de la production d’androgènes, cette réponse est considérée comme une étape préliminaire qui précède leur élimination par apoptose (60). L`ovocyte reste la dernière cible avant la disparition complète du follicule ovarien par atrésie (3).
Plusieurs facteurs peuvent contribuer dans la régulation du devenir folliculaire (croissance ou atrésie) suite à l’existence d’un équilibre majeur entre la croissance folliculaire et le processus d’atrésie au sein des follicules ovariens. Cela est dû à une harmonie établie entre l’expression et l’action de deux groupes de facteurs qui peuvent englober certains régulateurs endocriniens ainsi que des régulateurs intra-ovariens. Un groupe de ces facteurs module la prolifération et la croissance folliculaire tandis que l’autre assure la programmation de la mort des cellules folliculaires par apoptose (61). La croissance folliculaire est indépendante des gonadotrophines au cours des premières transitions folliculaires (du follicule primordial au follicule préantral). En revanche elle est régie par la participation directe des facteurs intra-ovariens comme le GDF9 qui est très
12
indispensable pour le développement folliculaire au cours des premières phases (62). À partir du stade antral, l’effet des régulateurs endocriniens sur la croissance folliculaire tel que la FSH devient crucial pour la poursuite de développement folliculaire (63), ce qui rend ces phases étroitement dépendantes de la présence de la FSH. Dans certains follicules l’expression des facteurs qui maintiennent la croissance folliculaire est défavorisé ce qui entraine l’atrésie folliculaire après l’expression des activateurs de l’apoptose dans ces follicules.
1.1.8. Ovulation
L’ovulation est un processus qui a lieu en réponse à un pic de l’hormone gonadotrophine LH sécrétée par l’hypophyse. Cette phase consiste à la rupture de la paroi du follicule ovulatoire pour libérer un ovocyte mature et fécondable. De nombreuses cytokines inflammatoires (interleukines, TNF) ainsi que des histamines et des prostaglandines sont communes entre la réaction inflammatoire et l’ovulation, de ce fait cette dernière est considérée comme une réponse de type inflammatoire (65). En réponse à un pic de LH, des agents inflammatoires provoquent une vasodilatation associée à une stimulation d’enzymes protéolytiques qui assurent la dégradation de la matrice extracellulaire pour faciliter la rupture de la paroi folliculaire et la libération de l’ovocyte (66). L’ovulation se manifeste par plusieurs aspects morphologiques et métaboliques, tel que la transformation des cellules de la granulosa et de la thèque en cellules lutéales (67). Ces dernières
Figure 1.4 : Régulation du développement folliculaire par les deux facteurs FSH et GDF9 (Avec obtention des droits d’auteurs)
13
contribuent à la synthèse de la progestérone qui est une hormone indispensable au maintien de la gestation. L’augmentation des taux de la progestérone est associée avec la diminution de la sécrétion d’œstradiol par les cellules folliculaires. Ces changements hormonaux sont accompagnés d’une diminution suivie d’une disparition des récepteurs à la FSH sur les cellules de la granulosa (3). Après le pic de la LH, l’ovocyte reprendra sa division méiotique pour atteindre le stade métaphase II, il est donc prêt à être fécondée. Après l’ovulation, le reste du follicule ovarien se transforme en corps jaune qui sécrète la progestérone.
1.1.9. Lutéinisation
Après l’ovulation, une structure bien particulière est constituée à partir du follicule ovulatoire pour former le corps jaune. Ce dernier tire son origine des cellules folliculaires qui subissent des transformations majeures suite à l’ovulation. Le corps jaune est considéré comme une glande endocrine assurant la sécrétion de progestérone afin de préparer l’implantation du zygote et soutenir la gestation. En revanche, le corps jaune est éliminé par un mécanisme de lutéolyse en absence de gestation (68).
Après l’ovulation et durant la formation du corps jaune, les cellules de la granulosa augmentent en taille et se remplissent de vésicules lipidiques lui permettant de se transformer en larges cellules. Ces dernières se situent au centre du corps jaune, tandis que les cellules qui se retrouvent dans la périphérie sont plus petites. Ces cellules périphériques proviennent de la transformation des cellules de la thèque en cellules lutéales. La restructuration tissulaire des cellules folliculaires et leur différenciation semblent être nécessaires à la formation du corps jaune (69).
Durant la phase lutéale, les changements morphologiques des cellules lutéales qui forment le corps jaune sont aussi associés à des modifications de l’activité sécrétrice.
Les cellules de la granulosa réduisent la sécrétion des enzymes de la stéroïdogenèse, afin d’acquérir d’autres enzymes indispensables pour la production de la progestérone telle que la 3β- hydroxystéroides déhydrogénase (3β HSD) (41). Toutefois, chez certaines espèces, la production de l’œstradiol reste toujours maintenue par les cellules lutéales (69).
Il existe un équilibre entre le développement et la régression du corps jaune qui dépend de l’action des facteurs lutéotrophes (favorisant la persistance du corps jaune) et des facteurs lutéolytiques (favorisant la régression du corps jaune). Parmi les facteurs lutéotrophes les plus reconnus, on
14
retrouve l’hormone gonadotrophe lutéinisante (LH) et la prolactine (70). Cependant, la prostaglandine F2α (PGF2α) est considérée comme un agent lutéolytique chez la plupart des espèces étudiées (71).
1.2. Maturation du complexe ovocyte-cumulus
1.2.1. Maturation de l’ovocyte
Plusieurs aspects morphologiques marquent la maturation de l’ovocyte qui est induite par un pic ovulatoire de LH (72). Cette maturation englobe la maturation nucléaire, la maturation cytoplasmique et la maturation moléculaire. Durant cette phase de maturation, l’ovocyte qui a été bloquée en prophase I depuis le stade de follicule primordial, est maintenant prêt à compléter la méiose. Le processus de la maturation nucleaire débute par la disparition de la membrane nucléaire qui se poursuit par une condensation des chromosomes et la formation de la métaphase I (MI). Ces modifications sont complétées par l'émission du premier globule polaire et bloquée en métaphase II. La maturation cytoplasmique et la maturation moléculaire sont caractérisées par d’autres aspects tels que la réorganisation du cytosquelette, la redistribution des mitochondries autour de la région nucléaire, un changement morphologique des gouttelettes lipidiques ainsi qu’une variation dans la composition du cytoplasme en métabolites (glutathione, cystéine, adénosine triphosphate …) (72). Ensembles, ces trois types de maturation confèrent à l’ovocyte son aptitude à supporter avec succès la fécondation et par la suite le développement embryonnaire. On parle d’acquisition de la compétence au développement (73).
1.2.2. Expansion du cumulus
Au cours de la maturation, les cellules de cumulus subissent une importante expansion qui correspond à un changement de l’organisation des jonctions perméables entre les cellules permettant les échanges. Cette modification résulte d’un accroissement des espaces intercellulaire menant à la dissociation des cellules du cumulus et d’une diminution des connexine 43 dans les jonctions communicantes (74). Durant cette expansion, les cellules du cumulus développent une matrice extracellulaire composée principalement d'un réseau d'acide hyaluronique grâce à une enzyme Hyaluronan synthase 2 (HAS2). Cette matrice extracellulaire est solidifiée par certaines protéines de liaison de l’acide hyaluronique dont versican (gène stimulé par le facteur nécrotique tumoral 6 ) et la pentraxine 3 ou l’inhibiteur de la trypsine inter-α (65). Ainsi, la matrice extracellulaire
15
contribue à la protection du complexe ovocyte-cumulus lors de son parcours dans l’oviducte et facilite aussi le processus de fécondation (75). La stimulation de ce processus d'expansion est également assurée par certains facteurs de croissance (EGF, IGF1) et des facteurs ovocytaires (GDF9) qui peuvent agir sur les cellules du cumulus pour moduler la synthèse d’acide hyaluronique et donc la matrice extracellulaire (43).
1.2.3. Régulation de la reprise méiotique
La reprise de la méiose par l’ovocyte correspond à la transition G2/M du cycle cellulaire. Cela est associé à l’activation du facteur de promotion de la phase-M (MPF) (76). Ce dernier est un hétérodimère composé d’une sous unité catalytique, la kinase cycline-dépendante 1 (CDK1) et d’une sous unité régulatrice, cycline B. Le profil d’activité du MPF présente deux pics entre la reprise méiotique et la métaphase II. La première phase montre une activité élevée du MPF qui baisse lors de la transition méiose I et II suite à la dégradation de la cycline. Cette dernière est neosynthétisée durant la deuxième phase, ce qui augmente l’activité du MPF de nouveau lors de la métaphase II jusqu’à la fécondation (77). La synthèse et la dégradation des cyclines B1 et B2 peuvent être considérées comme des niveaux de régulation de l’activité du MPF qui dépend de la formation de complexe cycline B- CDK1. Ainsi, l’état de phosphorylation de ce complexe est une autre cible de contrôle de l’activité MPF.
1.2.4. Mécanisme de reprise de la méiose
Le blocage méiotique des ovocytes dans les follicules ovariens semble avoir une relation avec les niveaux intra-ovocytaire en AMPC (78, 79). Gràce aux jonctions communicantes, des échanges
d’AMPC peuvent être établis entre l’ovocyte et les cellules de cumulus afin de maintenir un niveau
élevé en AMPC. Chez l’ovocyte, l’adénylate cyclase (AC) et la phosphodiéstérase 3A (PDE3A)
contribuent respectivement dans la synthèse et l’hydrolyse d’AMPC (80). L’inhibition de la PDE3A par
le GMPc transféré des cellules de la granulosa participe entre autres dans l’augmentation des niveaux d’AMPC en inhibant son hydrolyse (81). De ce fait, la régulation de ces enzymes peut aussi moduler les niveaux élevés d’AMPC. Ces derniers, maintiennent l’activation de la protéine kinase
AMPC dépendante (PKA) qui assure un contrôle négatif du MPF. Cela va empêcher la reprise
16
Avec le pic de gonadotrophines ovulatoires, les taux d’AMPc intra-ovocytaire baissent considérablement. Cela est associé avec une diminution importante de la perméabilité inter-cellulaire bloquant ainsi l’entrée d’AMPC dans l’ovocyte. Ces changements physiologiques favorisent la
maturation ovocytaire et la reprise de la méiose en présence des taux faibles en AMPc intra-ovocytaire (82).
1.3. Phosphodiéstérases
1.3.1. Classification
Les nucléotides cycliques sont des seconds messagers intracellulaires qui sont impliqués dans la signalisation cellulaire en réponse à différents stimuli. La 3′,5′- adénosine monophosphate cyclique et la 3′,5′- guanosine monophosphate cyclique sont les deux formes les plus connues des nucléotides cycliques. Leur synthèse est assurée respectivement par l’adenylyl-cyclase (AC) et la guanylyl- cyclase (GC) tandis que leur dégradation est considérée comme la principale fonction des phosphodiéstérases (PDE). Ces dernières appartiennent à la superfamille des métalophosphohydrolases, elles hydrolysent le groupement phosphate en 3’ des nucléotides cycliques pour produire un nucléotide 5’ phosphate. De ce fait, les PDE possèdent un rôle essentiel dans la régulation cellulaire et le maintien de l’homéostasie en agissant sur la transmission du signale par les nucléotides cycliques (83). Jusqu’à présent, 11 familles de PDE ont été identifiées chez les mammifères (PDE1- PDE11), codées par 21 gènes (1 à 4 gènes pour chaque famille). Ces protéines ont été classées selon certaines propriétés biochimiques telles que la séquence en acides aminés, la sensibilité à divers nucléotides cycliques et les agents pharmacologiques ainsi que les propriétés cinétiques de ces enzymes (84). Les gènes des PDE codent pour de multiples protéines grâce à l’épissage alternatif ou par l’utilisation de multiples promoteurs qui engendrent la synthèse de plusieurs isoformes. Chez les mammifères, certaines familles de PDE hydrolysent spécifiquement l’AMPC (PDE4, PDE7 et PDE8) tandis que d’autres hydrolysent seulement le GMPC (PDE5, PDE6 et
PDE9). En revanche, les familles (PDE1, PDE2, PDE3, PDE10 et PDE11) peuvent hydrolyser les deux nucléotides cycliques (AMPC et GMPC) (85).
17
Avec l’alignement des séquences, les 11 familles de PDE présentent une structure de base commune (Figure 2.5) avec un domaine catalytique en C- terminal très conservé et un domaine régulateur en N-terminal qui varie amplement d’une famille à une autre. Cette organisation confère à chacune des familles des propriétés biochimiques spécifiques qui pourront contribuer à leur régulation. Le domaine catalytique contient des motifs communs HD(X2) H (X4) ainsi que le site de liaison des ions Zn2+ ou Mg2+ (86). Les PDE peuvent fixer les nucléotides cycliques grâce au domaine N-terminal qui est spécifique à chaque famille de PDE (87).
La famille PDE1 comporte trois gènes : PDE1A, PDE1B et PDE1C qui peuvent générer plusieurs isoformes. Cette famille est activée en présence de calcium par une protéine thermostable, la calmoduline. La PDE1 semble être exprimée en grande quantité dans le cerveau, ainsi que dans le cœur, le muscle (squelettique et lisse) et autres tissus périphériques. Chez la PDE1, la localisation tissulaire ainsi que l’hydrolyse des nucléotides cycliques diffèrent d’une sous famille à une autre (89). Parmi les différente isoformes, la PDE1C est exprimée majoritairement dans le cerveau et le cœur
18
(90). Cette variante hydrolyse les deux nucléotides cycliques avec la même affinité tandis que la PDE1A et PDE1B présentent une grande affinité spécialement pour le GMPC (91).
Une autre famille PDE2 a été identifiée, les trois isoformes de cette classe (PDE2A1, PDE2A2, PDE2A3) sont codées par un même gène (92). Les deux nucléotides cycliques peuvent être hydrolysés par la PDE2 avec une légère affinité pour l’AMPC. En revanche, la stimulation de cette
classe est assurée surtout par le GMPC. Ce dernier exerce un effet allostérique en se fixant sur le
domaine GAF de la PDE2, cela contribue dans la régulation des deux nucléotides cycliques (93). La localisation cellulaire de chaque isoforme dépend en particulier de la région N-terminale qui diffère d’une forme à une autre, alors qu’elles sont caractérisées par une partie C-terminale conservée. La PDE2 est majoritairement présente dans le cœur, le cerveau et le foie mais elle peut aussi se retrouver dans d’autres types cellulaires. Au sein de ces cellules, les différentes isoformes se partagent entre la fraction cytosolique (PDE2A1) et la fraction membranaire (PDE2A2/3) (94).
Deux gènes codent pour les isoformes de la PDE3 qui sont PDE3A et PDE3B. Cette famille peut hydrolyser l’AMPC ainsi que le GMPC, par contre ce dernier pourrait contribuer dans l’inhibition de
cette PDE en rentrant en compétition avec l’AMPC (95). La partie C-terminale de la PDE3 comporte
des acides aminés additionnels par rapport aux autres PDE, ce qui pourrait intervenir dans la régulation physiologique de cette classe d’enzymes. Ainsi, chez les plaquettes sanguines humaines il a été démontré que la PDE3 possède la spécificité d’être activée indépendamment des voies de phosphorylation PI3K/PKB tandis que sa phosphorylation requiert l’activation de la protéine kinase C (PKC) (96). Les niveaux intracellulaires en AMPC peuvent être équilibrés grâce à la PDE3A qui est
considérée comme la principale PDE chez l’ovocyte de mammifères (97).
Les membres de la famille PDE4A sont spécifiques à l’AMPC parce qu’ils hydrolysent spécifiquement
ce nucléotide dans la cellule (98). Les PDE4s sont codées par quatre gènes notés A, B, C et D qui peuvent générer plus de 20 isoformes par épissage alternatif (99). La partie N- terminale de cette famille d’enzyme est caractérisée par la présence de régions hautement conservées appelées (upstream conserved region : UCR). Ces dernières contribuent principalement dans la régulation de PDE4 suite à sa phosphorylation par la PKA (100). De ce fait, cette phosphorylation est capable de réguler l’activité catalytique de la PDE4. Les domaines catalytiques de ces PDE possèdent des sites de liaison avec certaines protéines lui permettant de se retrouver dans différents compartiments cellulaires (101).
19
La famille de PDE5 hydrolyse spécifiquement le GMPC, elle est représentée par trois isoformes
nommés respectivement PDE5A1, PDE5A2 et PDE5A3. Ces trois derniers sont codés par un même gène (102). L’activation de la PDE5 est induite par sa phosphorylation suite à la fixation du GMPC. La
PDE5 a rapidement été connue suite à son implication dans le traitement des troubles d’érection masculine (103). Cette anomalie est corrigée par l’utilisation de sildenafil ainsi connu sous le nom viagra, un inhibiteur spécifique de la PDE5 (104).
La famille PDE6 englobe des enzymes qui sont retrouvées chez la rétine et qui hydrolysent exclusivement le GMPC. Elle est essentiellement impliquée dans un mécanisme de phototransduction
dans la rétine (105). Cette famille est représentée par trois gènes (PDE6A, PDE6B et PDE6C) et comporte aussi deux sous-unités PDE6 γ et PDE6 δ. L’activité de la PDE6 est en étroite relation avec ces deux sous unités qui régulent la stimulation de la PDE6 lors d’une cascade de phototransduction induite par le GMPC. Ce processus demeure régulé par la fixation de la sous unité
PDE6γ sur la sous unité α de la transducin, cela va permettre l’hydrolyse du second messager GMPC
par cette famille de PDE et la transmission du signal (106, 107).
Les PDE7 hydrolysent spécifiquement l’AMPC, elles sont codées par deux gènes PDE7A et PDE7B
et sont principalement exprimées sur les cellules lymphocytaires ainsi que le cerveau. Cette famille de PDE est impliquée dans la réaction inflammatoire, elle stimule la prolifération cellulaire des lymphocytes T afin de déclencher un processus inflammatoire (108). De ce fait, l’utilisation de la PDE7 comme cible thérapeutique contre l’inflammation a marqué un grand intérêt (109). Cette approche est basée sur l’inhibition de cette enzyme par des inhibiteurs spécifiques qui vont réduire l’intensité de l’inflammation en réduisant l’hydrolyse de l’AMPC par la PDE7. Des inhibiteurs
spécifiques de la PDE7 sont toujours en développement afin de traiter certains troubles neurologiques (110).
La PDE8 est la famille de PDE qui nous intéresse dans ce projet de recherche. Elle hydrolyse spécifiquement l’AMPC avec une grande affinité comparativement à la PDE4. Ainsi, cette famille de
PDE est insensible à l’IBMX, un inhibiteur non spécifique des PDE (111). Cette classe comporte spécifiquement deux domaines dans la portion N-terminale, soit le domaine REC et le domaine PAS. Le domaine REC ressemble aux domaines « receveurs » des systemes de signalisation retrouvés chez les bactéries tandis que le domaine PAS se trouve dans de nombreuses protéines de transduction de signal (88). La famille PDE8 consiste aussi en deux gènes qui codent pour deux
20
isoformes : PDE8A et PDE8B. Il existe cinq variants pour la PDE8A (1-5) obtenus par un épissage alternatif, l’expression de la PDE8A1 est la plus abandante par rapports aux autres variants (2-5). A son tour, la PDE8B possède 4 variants (1-4) (88). L’expression des deux isoformes (PDE8A et PDE8B) est spécifique aux tissus, elles sont présentes dans les cellules stéroïdogènes ce qui confirme leur implication dans la régulation de la production des stéroïdes (112). Ainsi, la PDE8A semble impliquée dans l’activation de la réponse immunitaire par la stimulation des cellules T (113). Dans les cellules de Leydig, la PDE8B est largement distribuée dans le cytosol tandis que la PDE8A est plutôt localisée avec la mitochondrie (114).
La famille PDE9 hydrolyse spécifiquement le GMPC avec une grande affinité comparativement aux
autres PDE. Un seul gène codant pour un unique membre PDE9A, a été identifié. Cette famille est exprimée dans plusieurs tissus tels que le cerveau, les reins, la rate ainsi que la prostate (115). L’utilisation des différents inhibiteurs connus n’a donné aucun effet sur la PDE9A. De ce fait, cette famille a été caractérisée comme insensible à une grande variété d’inhibiteurs. En revanche, des perturbations dans la reconnaissance chez les rongeurs ont été induites suite à l’utilisation d’un nouvel inhibiteur sélectif (BAY73-6691) (116).
Dans la même période, une autre famille de PDE, la PDE10 a été identifié. Cette dernière hydrolyse les deux nucléotides cycliques avec une plus grande affinité pour l’AMPC (117). Cette famille
d’enzyme est exprimée par un seul gène (PDE10A) et représentée par quatre isoformes (PDE10A1, PDE10A2, PDE10A3, PDE10A4). La distribution de la PDE10 est très restreinte comparativement aux autres PDE, elle est localisée surtout dans le cerveau ainsi que les testicules (118). De ce fait, cette PDE a montré un grand intérêt comme cible thérapeutique dans certaines maladies neurologiques telle que la schizophrénie en utilisant des inhibiteurs spécifiques qui ciblent l’activité hydrolytique de cette famille de PDE (119).
La PDE11 est la dernière famille de PDE qui a été identifiée, elle hydrolyse les deux nucléotides cycliques. Un seul gène code pour quatre isoformes qui présentent des différences dans la partie N-terminale (120). Les fonctions biologiques de cette famille ainsi que sa distribution sont mal connues en raison d’absence d’un inhibiteur spécifique pour la PDE11 (121).
21
22
1.3.2. Distribution des phosphodiéstérases dans
l’ovaire
L’activité catalytique des phosphodiéstérases a été détectée dans différentes cellules du follicule ovarien (122). Une étude chez le rat a rapporté la présence d’une activité PDE dans les cellules de granulosa, suite à une stimulation avec le Ca2+. Ces résultats reflètent la contribution de la famille
PDE1 qui est activée par le complexe Ca2+/calmoduline (123). Une autre étude chez le porc a montré
le rôle primordial que présente la PDE3A durant le processus de la reprise méiotique, cela était aussi associé avec la localisation de cette PDE dans l’ovocyte ainsi que son importante activité pour la maturation et la reprise de méiose (124, 125). Des variants de PDE4 ont été mis en évidence dans les cellules de granulosa et les cellules de la thèque. En revanche, une altération de la transmission du signal ovulatoire par les cellules folliculaires a été observée chez des souris déficientes du gène de PDE4D (126). Ces conclusions laissent suggérer l’implication de cette enzyme dans la fertilité. Ainsi, une expression fonctionnelle de la famille PDE8 dans le complexe ovocyte-cumulus a été démontrée chez le bovin, en se basant sur l’insensibilité que présente cette enzyme à l’IBMX (127). Le rôle précis de PDE8A dans l’ovaire n’est pas encore clair en raison d’absence d’inhibiteur spécifique pour cette classe (128). En revanche, la présence de cette enzyme dans les cellules du cumulus ainsi que l’ovocyte suggère qu’elle pourrait être impliquée dans la reprise de la méiose (52). En conclusion, toutes ces études nous permettent de considérer la PDE3 comme une PDE prédominante dans l’ovocyte tandis que la PDE8 et la PDE4 seraient prédominantes dans les cellules somatiques du follicule ovarien. Ainsi, la détermination de la distribution des PDE dans l’ovaire permet de mieux comprendre les voix de signalisation des nucléotides cycliques au sein des cellules folliculaires. Or, elle pourrait générer une nouvelle cible thérapeutique en ciblant chaque classe d’enzyme avec des inhibiteurs spécifiques (127).
1.4. Compartimentalisation de l’AMPc dans la
mitochondrie
La mitochondrie possède un rôle majeur dans la production d’énergie au sein de la cellule (129). Elle assure la régulation des différents processus physiologiques tels que le métabolisme énergétique, la régulation du cycle cellulaire ainsi que l’apoptose (130). De ce fait, cet organite cellulaire semi autonome pourrait occuper une place importante dans la signalisation cellulaire. Cependant, les
23
cascades de signalisation au sein de la mitochondrie restent peu connues. Certaines études ont montré la contribution de plusieurs molécules telle que l’AMPC dans ces mécanismes de signalisation
(131). L’adenylyl cyclase, une source de synthèse de l’AMPC a été mise en évidence dans la
mitochondrie, ce qui laisse suggérer une origine intra-mitochondriale de ce nucléotide cyclique (132). Ainsi, la régulation de ce dernier implique la contribution de la PDE, une enzyme de dégradation. Une étude récente chez les rongeurs a montré la présence de l’isoforme PDE2A dans la mitochondrie et cela dans le cerveau et le foie. Il a été rapporté que cette PDE est impliquée dans la régulation de l’activité de la chaine respiratoire (92). Une autre étude sur la régulation de la stéroïdogenèse dans les cellules de Leydig a également localisé la PDE8A dans la mitochondrie chez la souris (114). De ce fait, les PDE pourraient occuper un rôle crucial dans la signalisation cellulaire au sein de la mitochondrie.
1.4.1. La mitochondrie
1.4.1.1. Structure
Les mitochondries sont des organites semi-autonomes avec une taille qui varie de 1 à 10 µm de long et de 0,5 à 1 µm de large. Le nombre de mitochondries par cellule dépend du type cellulaire ainsi que des besoins énergétiques (133). Il y a entre 500 et 2000 mitochondries par cellules. Cependant, chez la souris on estime le nombre de mitochondrie par ovocyte à 90000 mitochondries. En revanche, chez l’humain ce nombre varie considérablement durant le développement folliculaire ainsi que la croissance ovocytaire (134).
Toutefois, ces organites présentent des formes et des structures extrêmement variées. Elles adoptent une forme allongée dans les cellules somatiques telles que les cellules hépatiques chez les humains (135). Tandis que les mitochondries des cellules germinales (ovocytes) sont petites et d’une forme spécifiquement ronde (136).
Ainsi, les mitochondries sont constituées de deux compartiments (un espace intermembranaire mitochondriale et une matrice) délimités par deux membranes mitochondriales (interne et externe) (137).
- La membrane externe : cette membrane présente des similarités avec la membrane plasmique des eucaryotes. Elle est sous forme de bicouche lipidique contenant environ 60 % de protéines et 40 % de lipides. Ainsi, elle est perméable aux molécules avec une taille inférieure à 10
24
kDa telles que des anions, des cations, différents métabolites (acides gras, pyruvate…) et des nucléotides. Cette caractéristique de perméabilité est assurée grâce à la contribution d’une protéine transmembranaire VDAC (Voltage-Dependent Anion Channel). Cette dernière, se présente sous forme d’un canal non spécifique pour faciliter le passage des molécules. En revanche, le transport spécifique des molécules avec un grand poids moléculaire est assuré par des protéines transmembranaires les translocases (TOM : Translocase of the Outer Membrane) (138).
- La membrane interne : contrairement à la membrane externe, la perméabilité de cette membrane est moins importante. Cependant, sa composition diffère de celles des autres membranes biologiques, sa teneur en protéines est de 75 % et 25 % en lipides. Cette particularité provient de la présence en quantité importante d’un phospholipide, cardiolipine. Cela réduit la perméabilité de cette membrane aux molécules polaires ainsi que les ions. En revanche, le transport des molécules à travers la membrane interne requiert des transporteurs spécifiques. Ainsi, cette membrane présente des invaginations qui forment des crêtes mitochondriales, renfermant la majorité des enzymes de la chaine respiratoire ainsi que l’ATP synthase (139).
- La matrice : est un compartiment mitochondrial interne délimité par la membrane interne. Elle renferme plusieurs enzymes impliquées dans le cycle de Krebs ainsi que l’oxydation des acides gras. Elle comporte aussi l’ADN mitochondrial, les ARN de transfert et les mitoribosomes.
- L’espace intermembranaire : ce compartiment se trouve entre les deux membranes avec une composition proche de celle du cytosol. Très riche en protons ainsi que de nombreuses protéines (140).
1.4.1.2. Fonction
Dans les cellules eucaryotes, les mitochondries sont considérées comme la principale source d’ATP en transformant les nutriments en énergie utilisable selon un processus de phosphorylation oxydative (141). En plus de leur rôle déterminant dans le métabolisme énergétique, les mitochondries contribuent aussi à la signalisation cellulaire en fournissant de l’ATP, la différenciation cellulaire ainsi que l’apoptose (142).
25
Pour répondre à ses besoins, la cellule nécessite de l’énergie produite par la chaîne respiratoire de la mitochondrie sous la forme d’ATP. Ce processus implique l’oxydation des nutriments cellulaires ainsi que des molécules organiques, cela va générer un gradient électrochimique qui résulte en une phosphorylation de l’ADP en ATP (142). Le système de phosphorylation consiste en une chaine respiratoire mitochondriale ainsi que l’ATP synthase. Cette chaine respiratoire comporte quatre complexes protéiques localisés sur la membrane mitochondriale interne et subdivisés en différentes sous unités.
- Complexe I: aussi connu sous le nom NADH: ubiquinone oxydoréductase. C’est la première enzyme de la chaine respiratoire contenant le premier accepteur d’électrons " la flavine ". Elle catalyse la réaction de réduction de l’ubiquinone par un transfert d’électrons du NADH. Ainsi, avec un poids moléculaire de 980 kDa, cette enzyme est considérée comme le plus gros complexe enzymatique membranaire avec une structure en forme de L. Ce dernier possède un domaine hydrophile comportant le site catalytique de l’enzyme (143). Cependant cette enzyme comporte quarante-six sous-unités, certaines sont codées par l’ADNmt alors que d’autres sont codées par le génome nucléaire. En revanche, plusieurs études ont montré l’implication de ce complexe dans le développement de certaines atteintes neurodégénératives tel que la maladie d’Alzheimer. Cela est lié avec la survenue de certaines mutations qui vont engendrer une augmentation de la production d’espèces réactives de l’oxygène (DAO) par ce complexe (144).
- Complexe II : ou succinate déshydrogénase, le composant le plus simple mais il demeure très important dans la mitochondrie. Grâce à son activité oxydoréductase, le complexe II implique la connexion du cycle de Krebs à la chaine respiratoire suite à un transfert d’électrons du succinate à l’ubiquinone (145). Ce complexe est composé généralement de quatre sous unités (A à D), codées par quatre gènes appartenant au génome nucléaire. Un domaine enzymatique hydrophile est constitué par les deux sous unités A et B qui sont la flavoprotéine et la protéine fer-soufre. Ces deux dernières représentent ainsi la fraction active de l’enzyme. En revanche, les deux sous unité (C et D) forment le domaine hydrophobe qui assure l’ancrage membranaire du complexe (135).
- Complexe III : c’est l’enzyme ubiquinone-cytochrome c oxydoréductase aussi connue sous le nom « complexe bc1 ». Ce complexe est ancré dans la membrane mitochondriale interne, il est considéré comme un transporteur d’électron de l’ubiquinol (forme réduite) au cytochrome c. Dans la membrane mitochondriale interne, cette enzyme est organisée sous forme de dimère. L’unité
26
structurale de ce dernier est un monomère formé de onze sous-unités qui sont presque toutes codées par le génome nucléaire (146). Le transfert d’électrons est assuré par les trois sous unités le cytochrome b, le cytochrome c1 et la protéine Rieske Fe-S. Cependant contrairement aux autres sous unités, seulement ces trois dernières contiennent des groupements prosthétiques (135).
- Complexe IV : ou cytochrome c oxydase, c’est le dernier complexe de la chaine respiratoire. Il est impliqué dans la catalyse de transfert des électrons du cytochrome c déjà produit par le complexe III, à l’oxygène(O2). Ce complexe enzymatique est composé de treize sous-unités,
certaines sont codées par l’ADNmt alors que les autres sont codées par le génome nucléaire (146). Cependant, le centre catalytique de cette enzyme est constitué par les deux premières sous unités (COX I et COX II) possédant chacune un groupe héminique et deux centres de cuivres CuA et CuB. Ainsi, le transfert d’électrons dans le complexe est assuré par le centre catalytique de l’enzyme tandis que le pompage de protons demeure le rôle de la troisième sous unité. En revanche, la fonction des autres sous unités reste encore mal connue (147).
1.4.1.2.2. L’apoptose
L’apoptose est un processus physiologique permettant d’induire une mort cellulaire programmée suite à un signal. Ce mécanisme est activé pour éliminer des cellules endommagées, mutées ainsi que des cellules âgées. Cette auto-destruction cellulaire est en équilibre avec le processus de prolifération cellulaire afin d’assurer la survie des organismes. Ainsi, le contrôle majeur de l’apoptose est gouverné par la mitochondrie qui est impliquée essentiellement dans l’activation de la voie intrinsèque de l’apoptose. En revanche, cette voie participe dans l’activation des caspases, une famille de protéases spécifiques. Ces dernières sont responsables de la dégradation cellulaire ainsi que la formation de corps apoptotiques (148).
Plusieurs études ont démontré que la maturation ovocytaire ainsi que le développement embryonnaire sont aussi affectés par l’apoptose, principalement induit par la mitochondrie. Cette activité apoptotique représente un mécanisme de contrôle de qualité qui est déclenchée en présence de conditions non optimales (149).