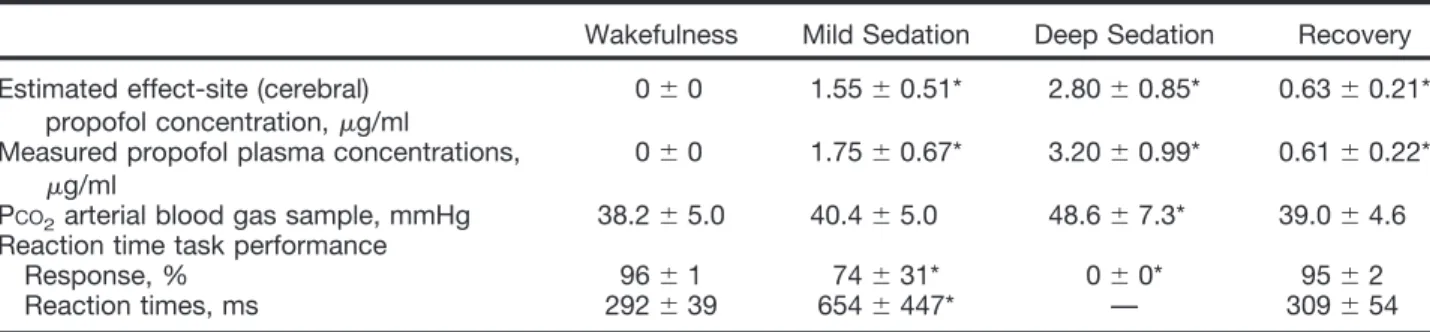
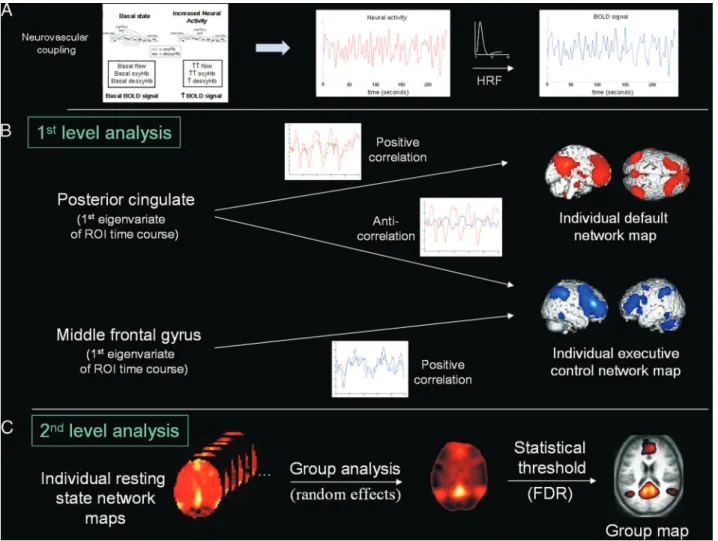
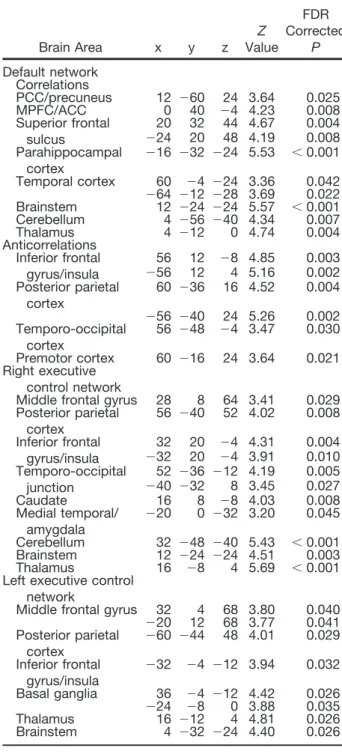
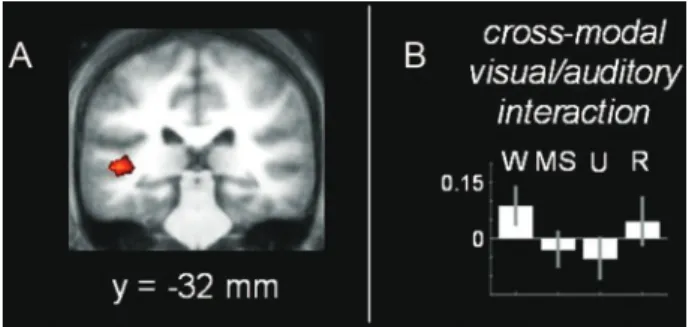
Breakdown of within- and between-network resting state functional magnetic resonance imaging connectivity during propofol-induced loss of consciousness.
Texte intégral
Figure


Documents relatifs
Different groups of people have to be considered: the employees working in the nearby areas might want to use it at lunch hour to meet each other and enjoy the
In this work, we introduce a novel approach for dynamical functional network connectivity for functional magnetic resonance imaging (fMRI) resting activity, which
L‟exploration de ces mêmes phénomènes dans un groupe de patients avec une durée de maladie plus élevée (8 ans en moyenne) révèle une augmentation de la connectivité
Neuropsychological measures (episodic memory, both immediate and delayed recalls, as measured with the Stories and Family Scenes CMS subtests) and psychopathological
In the go/nogo condition, we manipu- lated the time during which proactive inhibitory control had to be sustained by varying the delay between the beginning of a trial and the go
At each channel use, the transmitter aims to send informa- tion to the receiver for which reliable decoding at the highest achievable secrecy rate is guaranteed. As the transmitter
Our preliminary results demonstrate, using datasets coming from different databases, that a network embedding approach, combined with standard classification methods, provides
In this section, we take into account the ability of grouping individuals within two independent groups (e.g., the lead team and trail team) and the degree of interaction between
