(
.
.
l '
(
.
(
INHlélTION DE LA CROISSANCE DE 'NEISSERIA GONORJllJOEAE o '
PAR
DES STAPHYLOCOQUES COAGULASE-NEGATIFS
ISOLES DE LA FLORE UROGENITALE
..
.1par
, l LUCIE LAFOND
Mémoire prêsentê ~ la Facultê des études supêrieures et, de _ recherche, en vue de l' obtenti on du grade
de ma'ître ès sciences (M.Sc.)
< • •
,
Dèpartement de Mi clfobi 0 1 agie " , ' Collège'Macdonald de'l'universitê McGill
Montréal, Qùêbec, Canada Septembre 1981
.
' \ " ,.
1· l..
ft
((
O ' Short .1tle: ,INHIBITION DU GON~COQUE PAR DES STAPHYLOCOQUES
..,..
.
.
..
\
" 1l
~ ,i i '11
t '
~,
f1
f
J l l,
..
, '
(
M.Sc.
..
/ RESUMELUCIE lAFOND
Mi crobipl ogieINHIBITION
DE LA tROISSANCE DE
Neisseria gonorrhoeaePAR DES STAPHYLOCOQUES COAGULASE-NEGATIFS ISOLES DE
LA FLORE UROGENITALE
'
Pann; les microorganismes Neisseria 90no,~;hoeae, nous
négatifs isolês 'de la' flô;~ urogênitalè. Les activitês
antigonococci-~
---ques observées sur milieu solide semblent attribuables ~ des substan-- - ' " , _ _ _ _ _ _ _ _ _ _ _ ;1
, ces inhibitrices et leur production a variê en fonction de la
composi-..
tion du milieu de culture utilisê. Des activitês sous forme soluble ont êté obtenues de surnageants de cultures sur mil ieux semi -sol ides
.
. ainsi qu'en mil ieux liquides et celles-ci ont êt~ différenciées en '
,J'
deux types, soit bact~ricic\e et bactériostatique. En comparant la sub-stance bactêriostatiqte produite par l'isolat 66 à la substance
bacté-1 t1
, 1 •
ricide produite par " isolait 7 qui avaft déjà été purifiée, nous avons montrl! qu'il s'agi s sa it de Ideux 1 i poprotêi
n~s
• semb 1 ab 1 es" la 'pa rt ie"" 'protêique des inhibiteursiJ
et 66 semble être .re~ponsab"'è
'de'
"'ac,t--iVltê,,,,,,, , /~
",
anti gonococci que et possédai t un effe,t:':b' ~"'~~me--1tarrsn~es deux cas. que 66 appara'it différent des inhibiteurs gonococciques, produits par les staphylocoques, préc~emment rapportés
,. . dans 1 a 11 ttérature. , t " ' ,
.
1l
!
l t ~{
M.Sc.
ABSTRACT
LUC·lE LAFOND ) ". Microbiology • ,
INHIBITION OF Neisseria gmiorrhoeae GROWTH BY
COAGUlASE-NEGATIVE STAPHYLOCOCCI ISOLATED
FROM UROGENITAL FLORA
Among the microorganisms known to inhibit Neis8em gonorrhoeae
~' f\
.
growth, we studied coagulase-negative staphylococci isolated from the urogenital flora. The antigonococcal activities observed on solid medium seem to be a~tr;buted to inhibit()ry su~stances, and the;r production was related to the cultu,re'medium uséd. Activities in a
• ' 4 " , ' /
solùble form were obtained frqm>'supernatants of semi -sol id and 1 iqui d cul tures, and these were", dÙ;;erenti ated ; nto two types,
..
bact~
ostat; c", \\ .
and
bactêrici~aT. 'ê~mparison
of the bacteriostatic substance produced-.
,• A ~;
by isohte 66 with tbe bactericidal substance producedôy isolate 7
---.-4 'which had been already' purified, has revealed that they were two similar
(
lipoproteins. The protein component of inhibitors 7 and 66 seems to"be ,responsible for the antigonococcal activity and was found bac1;erici~al
in both cases. The bacteriostati c itlhibi tor appeared different from , al1 other gonococcal inhi bitors, produeed by staphYl oeoeci, al ready
'.
•
descri bed ; n the 1; terature.
.
.
'1 )' \ L•
" l 11
1 !,
( \r
, \ ',1 " 1~I
. i> .
) "
"
i,
,
( '
J,
,"
REMERe 1 J;MENTS
Je ~gire r~mercier sincêrement toutes les personnes qui ont
, ,
contribué de prês ou de 'loin' \ ,
a
,la réalisation de ce travail,a
-
)l'élaboration de ce m~moire, et tout particuliêrement:
, -)
Les docteurs Roger Knowles,et R.A. McLoed. directeurs du
D~par-,
.
tement de microbiologie du Collêge Mac~onald de l'université McGill, pour m'aval r acceptée conme candi date
a
1 a mattri se.Le docteur Vincent Porte1ance, directeur du Centre de recher,che
1
en bactêriologie (C .R.B.) de
.
li Institut Armand-Frappier (I.A. F.), pour,
son appui et sein e!"couragement. '
~
Le ~octeur Samjr Antoine S~heb, professeur au C.R.B. de l' I.A.F.,
.
pour la direction de 'ce travail effectué en collaborationlavec le ('
"-docteur Jean~ Guy Bisaillon.
Le docteur Jeân. Guy Bi saillon, professeur au C .R.B. de l' LA.F'."
..
pour avoir assumé l'entiêre responsabilité de la direction de ce
tra-,
vail suite au d~cêssubit du docteur S.A. Saheb.
le docteur Réjean Beaudet, professeur au C.R.B. de "LA.F., pour son aide et ses précieux conseils,' particulièrement lors de la
purifi-"
cation et de la caractérisation des produits inhibiteurs.
v
.
" , \, , 1 , ,'j
',' , . , J J c i1
" > ..
(
(
(,
.
' 1 y;f "Le"d.oct~ur
Michel Syl'véstr'e;, pr.tifesseur 'auC.'R.B\'
de "l' 1.A.~.,
pour ses conseils et ses encouragements.Mesdames Jocelyne Charron-Morin et Rita Alary ainsi que ,monS\ieur
, .
• r J ..
~OUiS "Ra.fHle, pOl.lr
y
ai "de",~~chnil'que pr~cieuse apport~e, au, cours de ce traVhn.,' - "6,
\ "
.
Mademoiselle ~y'via-G-lrardon, pour son aide lors de
la
~actylographie, de ce m~moi re •~:...- - ~
les,~tudiants du C.R.B:,'Richard Degr~, Michel Frenette, Andr~ Morin
--et ~enoit' Picard,/pour leur appui moral constant.
L'Institut Annpnd-'Frappier et la Direction G~n~rale de l'Enseignement _ Supérieur, 'pour mt avo;
t
accordé des bourses me permettant de poursui vre "1 mes études. ,"
.
, 1 tIF ) , , " ~ l, l'"
1 • ,"
,
1 1 \, , , , , ! ;,.
1 ! , i ;j
o
(
(
r,
a
TABLE DES MATIERES (
TITRE ••• ,. ••••••••• .- ... , .• RE SUME • ~ •• , • • • • • • • • • • • .. • • • • • • •••• Il • • • • ~ • • ,. • • • • • . . . • • • • • • • • • • • • • • • , ~ ABSTRAC T ••.• ,. •••••••••••••••••• ,. ••••• ~ ... " ••• ~ ••• . . DEDICACE. f ' l • • • • • • • • ,. .. ,. • • • • • • • • • • , • • • • • • f • • • • • I r ' • • ' . . . . REMERe 1 EMENTS ••••••••••• : ••••• ,. •• , i • • • • • • • • • • ' . . . ,. • • • • • • TABLE DES
LIS1
DES LISTE DES MATt E RE S. • • • ••• " • • • .. • • • • • • • • • . • • • • • • • • .. • • .J ••.•... TABLEAUX •••••••••••••••••••••• ~ ... . FIGURES •••••••••• ~ ••••• , .... Il • • • • • • • • " • • • • • • • • • • • • • • • • • ~ . . INTRODUCTION •••••••••••••••••••••••••.••••• ~ •• '.'Y" .'. ~ .•.•.•• :' ••..
. REVUE DE LITTERA~URE ,. •••••••••••••••••••••••••••••••••••••••••••• . vi; ii ii; ivv
viix
xii 1 3 1. Substances antimicrobiennes produites par les staphylocoques. cf" 51.1 1.2 1.3 1.4 Les staphylococcines ... f.'t . . . . l es an tl , 'b' a t' l ques ...•••...•....•..••...•... ~
les enzymes bactériolytiques ... ' ... . Autre .••....•••.•..••••••••••••••••••.••••••••••••••••••
5 8 . 9 11 2. Inhibition microbienne de N. gonorrhoeae... ... 11
" ,
2.1 Inhibition in vitl'o de N. gonol'rhoeae ... ... ,. 12 2.2 Substances antigonoc'occiques d'origine microbienne... 16 2.3 Inhibition in vivo de N. gonol'1'hoeae... ... 19 MATER! EL ET METHODES ... ". .. • .. • 22 1. Souches bactéri ennes ..•... : .•....•.•...•... , ... .
' .
.
'1.1 Isolats urog~nitaux ...•••....••••..• , •••. , ..••...•• 1 2 • Ne7,,88er1.,Q gonoT'p. oeae ••••• ••• ' ••••••••••••••••••••••••••• • . h ' \.
1 .3 Conse rv a t ; on. ; ... . 1 .4 Pr~cu l tures ... :-:\. ' .•...•.••..• ~ ..•.•• 1.5 Standrdisation des suspensions.',' .••••••.••••.••....• '. .• 2. Milieux
de~lture
... .-...~
... :2 • 1 Mi 11 eu x so 1 i de s ....•.•.•••••••••••••••••••••••••••••• l' • •
2.2 Milieux sem; -solid4s ...• . ,. ....•.•.•. t ...• f • • •
2.3 Mi lieux l ;qui des ... . , '23. 23 23 23 24 25 26 26 28 29 0
!
l
~î'
\ i , i ~ {(
1,
)
" il(
j
(;,
-3. Dé",", t ra t i
on
de.
l' a ct i vité an tiinococ
ci que, ur mi lie ux " solf~s ... p. ••••••••.•••••••••••••• ~ •••••••••••••••••••• : ••3. 1 Méthode di recte ... ., .... ~ ... ;. .... A . . . .
).2 Méthode "f1ip-floP agar overlay"-.~ ... ~ ... .
1.
Production de surnageants antigonococciques .•....••...•....JI
4.1 A partir de milieux solides ... . 4\2 A parti r de mil ieux li qui des . .' ... .. 4.3 A partir de milieux semi-solides.' ... .
vili 2Q 29 30-31
...".
31 32 1 0 5. DHe~tion de l'activité antigonococcique des surnageants.. .... 335.1 Diffusion en milieux solides ... .' ... 33
5.2 Concentration minimale inhibitrice: CMI1... 34
5.3 Concentration minimale inhit1itrice: CMI2... 34
5.4 Unités, àrbitraires... ... ... ... ... .... 35
~ 1 • 6. Cint:!tique
re
prod,ucti on: ... .. 6.1 En milieux liquides ... : .. 6.2 En milieux semi-solides ... " 7 C· • lnc ~t' lque d" lnac lva t . t . lon ... , ... - c ~8. DHermination de la concentration minimale qui tue plus de 99%
des ce 11 u l es de N. gononhoeae ...•...•...
-....
9. Caractérisation prê1iminaire ... ..
9)1'
abi lité aupH ... .
9.2 Stabilité à la chaleur ... ..::
~!: ~~:~; ~~" d~" p~i
d;"';';
i
~~~i ~;;~
.
p~;" ~i t;~f;i t;~t;~~:
: ::::
10. Puriflcation ...• ' ...•....•..•....••...••.... 11. Spectres ultraviolets ...•...•.•... 12. Traitement à l'ur~e ... : ...•...•....••... ; .. 13. Dosage des protéi nes ... ~ ...•... ~ .. ~ ... .14. Electrophorêse ... .
14.1 En gel d'agarose ... .
14.2 En gel de polyacrylamide ... .' ... . 15. D~termi nation du po~ ds mol écul ai re ... .. 15.1 Par filtration sur gel mol~cu1aire ... .. 15.2 Par ~lectrophorêse en gel de polyacrylamide en pr~ser'lce
d~ SDS .• ". ••.••.••.•..•••.••..•••.••••••..•••••..•••••.••• 36 36 37 38 39 ,39 39 40 40 40 41 43 43 ·44 44 44 46 47 47 48
{
-r 1 1 i 1 11.
.'...
..
>,(.
(
q 1~x
, . 1 ' . ( 16 P d• rOUl S c 't h" lmlq~s""~".,.".",,,""""" \ ' .. ' .. " " " · · ' · · · " " " 49 . A • ' • 51 , RESUL T TS. 1 . , • • • • • • • • ~ • If • • • • ' • • • • • • • • ! • • • • • • • • • • • • • • • • • • • • ~ . . . . ~ ,,1. Production d'activitéS antigonococciques sur milieux solides;. c, 52-2. Obtenti on dl act; yit~s anti gonococci ques sous forme 1 i qui èle • • .. 58 3. Caractérisation préliminairé et pu.rification ... ' ... :. 75 DISCUSSION ••• ! • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • ~ • • • • '
194 .,
, /
1. ,Production d'activités antigonococc;ques sur milieux solides.. 95 2. Obtention d'activités antigoQococciques sous fOl'illJe liquide.... 98
<
..
.
3 .. Caract~risation préliminafire et purification ... ' 108 CONCLUSION .•••••••••••••••••••••• ,. ••••.••••••••••• ) •.•• " • • . • • • • • • • • . 116 BIBLI()GRAPHI'E .•••••••..••••••• , ••••••.•••••..•• l • • • • • "' • • • • • • • • • • • • • • '\ 118 -, " ,.. fJ 12 ORIGINAUTE DU TRAVAIL... • ... ... 8 , , " , l
d ,
\\
1 1 1 •1
{..
.i
l.
~ 1 ~, , . 1 1'-
~....
f.
~. 1 , TABLEAU l 2(
4 5 7 f 1 ) 8..
('
j ,r ,~ !.. 't v -, / .,.
LISTE DES TABLEAUX
't PAGE
x
\
\
'.
,;Microorganismes inhibant la croiss~nce de
N. gonoZ'Z'hoeae in vitro
13~
Effet des différents mil jeux de culture sur , la production d'activit~s antigonococciques
par les isolats urog~n;taux .
lJf
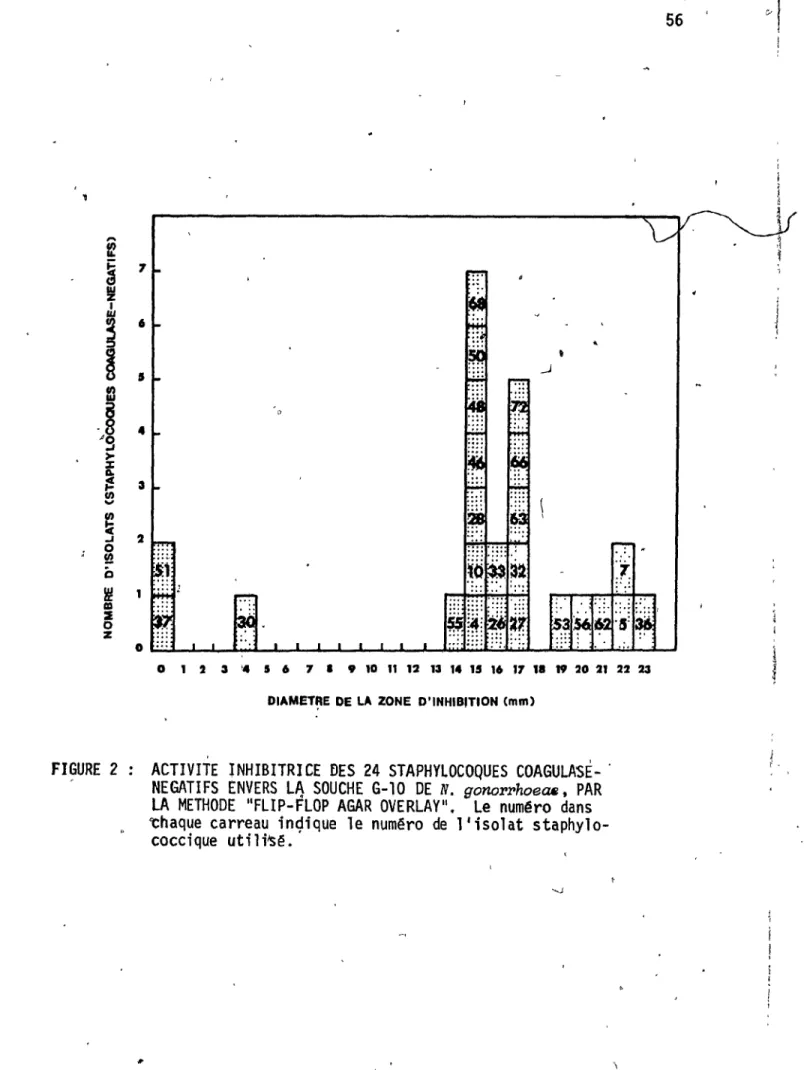
Effet du nombre dIU.F.C. de l'isolat inoculê sous ,forme de "spot fi sur 1 a pr~duct; oh d'
acti-lX
-vités antigonococciques C
..
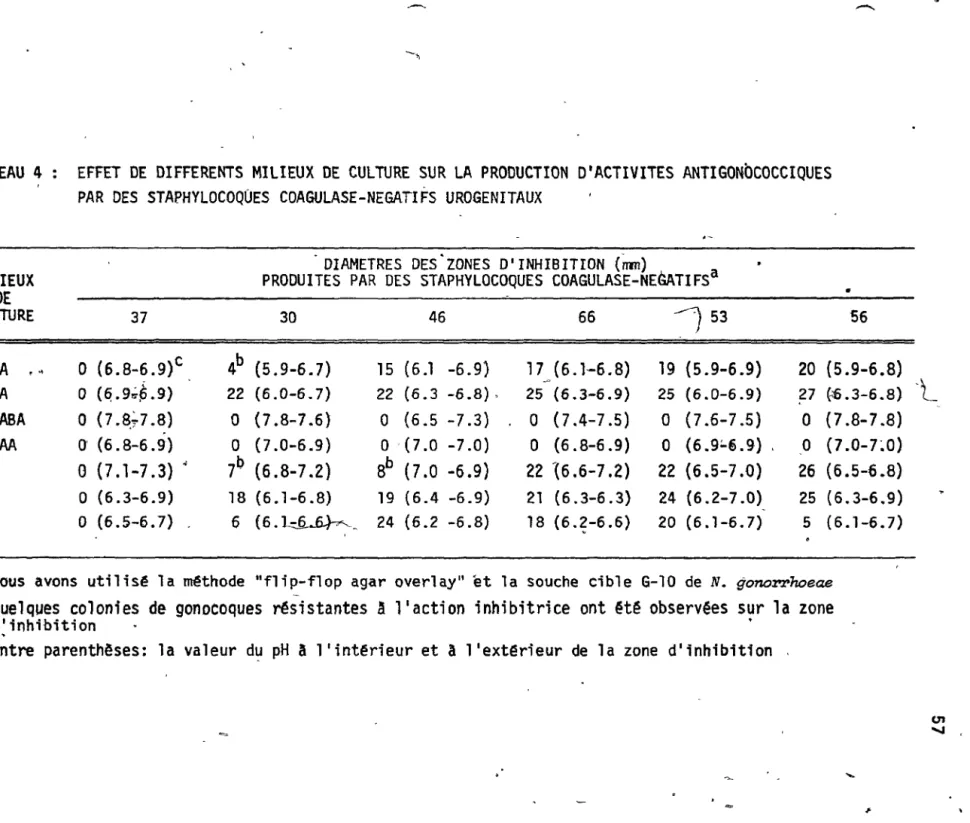
~- ~érents m,l • e'ux de cul tùt~ sur la
productionod'activit ococciques par'-des st~phy19coques coagulas négati!,s urogê-nitaux
~ ... _---... <ï J t
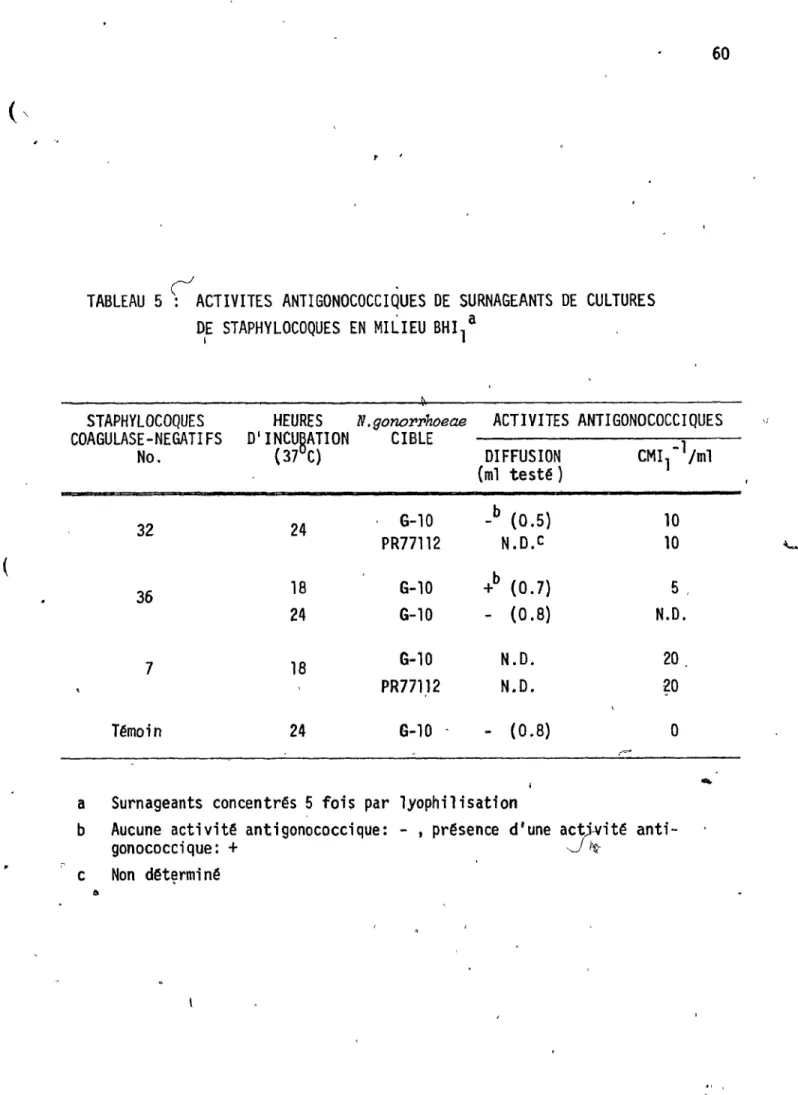
Activit~s ;n~~e surn eants de 0
cultures de staphylocoques en~
,
.
"
Effet de diff~rents mili~ux de cultures liquide sur la production d'activités antigonococciques
par l'isolat 32 f 53 54 57 60 61
'Détermination quantitatfve des activités anti- 66 gonococciques des sutnageants provenant des
cultures de staphylocoques
Détermination quantitative des activités anti- 71
, p
gonococClques des surnageants de culture de ,
"
, .
staphylocoques coagulase-négatifs
1
0
(-.
TABLEAU
9 ~ , "-. 10 11(
._1'
," i ' Puri fi cati on produi te pa rBH1l
Puri fi cation prOdui_te par BH!l ~ o - • de l'activité antigonococcique l'isolat 66 en milieu semi"solide"
de "act ; vi té ant i gonococc';, que l'isolat 7 en m"'eu semi-solide
DétenninaÙon de la concentration minimale, en unités arbitraires par ml, nécessaire pour
tuer plus de 99% des U.F.C. de la souche G-10
de N. gonorrhoeae inqculêes xi 1
!
1PAGE
.l
J ! J 781
!
,1 79 93 ).
FIGURE 1 2 3 ( 4 5 6 7 xii
....-LISTE DES FIGURES
0:> ' PAGE
.
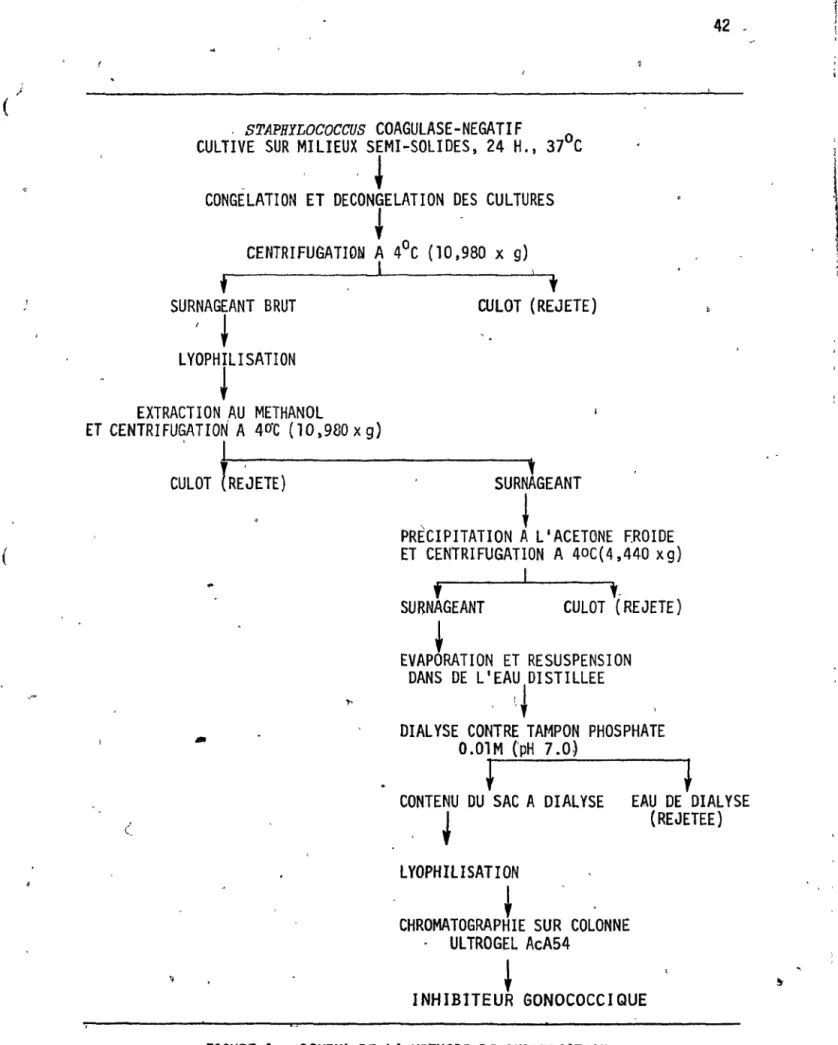
Sch~ma de la m~thode ~e purifiGation
,
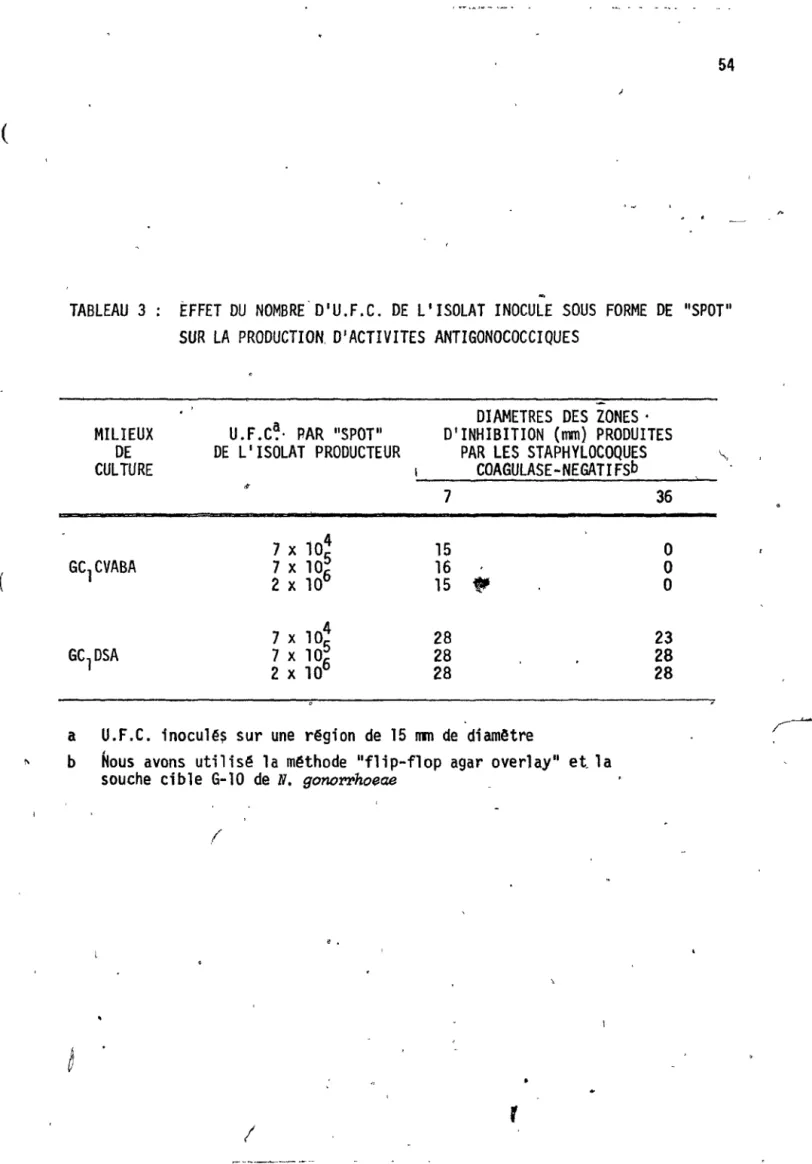
Activit~ inhibitrice des 24 staphylocoques coagulase-né,gatifs envers la souche G-10 de'
-
"N. gonol'l'hpeae, par la, méthode IIflip-flop , agar overl ay"
,
.
Ci nét i que de produe;tî on de l'actÙité anti-gonococcique du staphylocoque coagulase-néga-. ""tif 32, en milieu SHI, liqu;'de
Cinétique d'inactivation de la souche 6-10 de
N. gonorrhoeae par différentes concentrati~ns
de surnageant inhibiteur provenant d'une cultu-re de 18 heucultu-res de' l'isolat 32 en milieu BHI liquide'
Cinétique d'inactivation de la souche G-10 de
.
. ' N. gonol'1'noeae par différentes concentrations de surnageant inhibiteur provenant d'une culture de 24 heures de l'isolat 37 en milieu semi-solide
42 56
62
64
68
Effét du mili~u de culture sur~a croissance de 3 69 la souche G-10 de N. gonol'l'hoeae
.
'Pourcentage de densité optique de cultures de la sou- 72 che G-10 de N. gono1'rh~eae 'après 6 'heures
d'incuba-t.i on en présence de di fférentes concent rat; ons de s'urnageants tnhi biûteurs
: 1
'1
-(
((
.
-, , . \ "••
•
J
xiii fi FIGURE PAGE ( j 8 9· 10 11 12 13l<Çi~ét;ques'd'inactivation de' la souche G-10 de"
N. gono~rhoeae par deux types de surnageants: bactéricide '(isolat 37) et bactériostiitique (isolat 66,) .
74
.Cinétiques
de~pr~9uction
de l'activitê'anti- 76 , gonococcique de deux isolàts sta~hylococciques:l'isolat 37 (bactérici~e) et l'isolat 66
(bactério-s~atique)
Filtration
mOl~c~l~~re
sur UltrOgel'AèA54 de sur-nageants antigonococci~ues ~ialysésCourbe d'étalonnage obtenue par chromatographie de protêines de poids moléculaires connus sur Ul troge l AcA54
E1ectrophorêse en gel de polyacrylamide, en pré-. . sence de 0.1% SOS, ijes fractions AB des isolats 7 et 66
Spectres u1 travi 01 ets des
f~a~ti
onsAB
p'rovenant des isolats 7 et 6680
82
84
85
14 "QEle~rophorese en gel d'agarose des fractions 'AB ' .87
des iso1at~ 7 et 66
15 Filtration moléculaire sur Ultroge1 AcA54 des 88
16
"
fractions antigonococc;ques
AB
Courbes d'étalonnage ob~enue par chrOma~graPhie de 'Protéines de poids molêcula.ires conn~ sur' ) Ultt"99:1 AcA54 , . " \ . ' cp \II 89 f ,
,.
l l ,;'.
l
; j i(
(
FIGURE.
.
PAGE1 17 Electrophorêse en gels d'agarose des' fractions 91
/11,2,' 3 (U1.tr~el AcAS-4, 4 M u~êe) des isolats
'7 et 66
...
, ,.xiv
@ 1,)
1 1 1 1 \,1
t 1 )\
1
: , l -l 1 j J ; 1\
~.
(
(
•
l' ,INTRODueTION
la gonorrhêe figure parmi les infèctions bactêriennes les plus répandues. Au Quêbec, envir.on 5,000 nouveaux cas de gonorrhée sont ,décllrês annuellement, mai s on estime qu l'une personne ,~ur.200 en est
atteinte, soit environ 30,000 individus (Statistiqtle C~ada). A, .
travers le monde, on estime à plus de 70 millions de cas de gonorrhée
,
par ann~e (Roberts, 1977).
.,
les gonocoques sont devenus graduell~ment de plus ,en plus
résis-[ .. ~
tants
a
la pénicilline, a la tétracycline et aux autres antibiotiques durant les deux dernières décades (Jaffe et al., 1976; Sparling, 1972), et jusqula prêsent, aucun vaccin efficace contre plusieurs souches deNei88eria gono~rhoeae nia été développé. De cette situation pour le
"
moins inquiétante, il apparatt que d'autres mesures de contrôle de cette maladie sont nécessaires (Morriss et al., 1978).
1
Il est reconnu que tous les gens qui ont été exposés A N.~nor
rhoeae ne développent pas nécessairement ,une
g~~Orrh'ée
"clinique';'en~
identifiable. L'activité'antigonococcique' de certaines espèces bacté-riennes, présentes dans les voies ur~gênitales, serait une des explica-tians possibles de ce phénomène (Kraus et al., 1976), d'autant plus
,0 • • .,.
qu'on ne peut apparemment acqUérir que peu ou pas d'immunité contre cette
l ma1a4ie.
.
~ l\
)
• J 1..
((
..
(..
,.
-f~lusi'E!urs microorganismes de la" flore urog~nitale sont reconnus
p
J
pour' lnhiber la croissance de N. gonoPl'hoeae in vi.tro. Kay~ et Levison (1977), Saigh et at. (19?8j et Ingraham et ~t. (1978) ont,montrê que les staphyl ocoques ~tajent parmi l es pl us fréquents inhi bitèu'rs du 90nO-coqueo. Mais bien que ces activitês antïgonococciques aient êté ample~nt démontrées in-vitro, les substances impliquées .dans ces inhibitions ont
rarement été clairement identifiées .
.
.
L 1 étude de telles substances fabriquêes par les stapnylocoques
" '
2
pourrait donc ~ntribuer à la co~prêhension d~s phênomênes et m~canismes
intervenant
da~ l~s
relations entre N. gonorrhoeae et les staphylocoques de la flore urogénitale, et 'pourrait aussi êventuillement contribuer au dêveloppement de substances qui pourraient, @tre ~pplicables au niveau du "diagnostic, de l'épidémiologie et de la thérapie de l'infection gonococcique.-
.• J
If--~_-,
-(
"; 'v '13/
, (),.,.,.
( REVUE DE LITTERATURE . " j (..
,;., ~ ... ' """
, ,< ,.'
l '. i\
"\
~
,j
"1 , 1 ; ,"
L'interférence bactérienne est connue depuis ,longtemps. les premiêres descriptions d,'une intéraction ant'a'goniste entre diffé-rentes bactéries remontent probablement
a
celles de Past~ur et 'Joubert (1877) qui.ont noté, l'effet inhibiteur de plusieurs bact,é-'.
' .ries sur BaailluB anthro.ais. Ces premi êres 'études ont ét~ cependant ,
orientées davantage sur les implications biologiques que sur le
,
mécanisme d'action mis en cause ou la caractérisation chimique des agents inhibiteurs.
Aujourd'hui, nous savonS que 1 'jnhibition peut être due
a
uneJI>
compétition nut~itive,
a
la création de conditions défavorablesa
la croissance tel qu'un efret du pH ou encore A l'élaboration de sub-stances antimicrobiennes (Tagg et al., 1976; Shinefield et a~ .. , 1972).les substances bac~ériennes inhibitrices incluent une grande variété de produits tels que les antibiotiques "classiquesn de faible poids mo1écul ai re, 1 es produi
ts..,
ou déchets métabo~ i ques, les bactéri ophages, les bactériocines, les enzymes et les agents lytiques. '~.~En' ce qui nous concern,e, nous nous sommes intéressés aux produits inhibiteurs fabriqUéS par des staphylocoques isolés de la flore uro-gênitale et" plus particuHêrement ~ ceux possêdant une activité anti~
> • • gonococcique.
4 ; .
"
(
(
,\
"
, a
1. SUBSTANCES ANTIMICROBIENNES PRODUITES PAR lES STAPHYLOCOQUES
,
Cette section de la revue de littérature a été inspirée par la.
,
revue de Tagg et aZ. (1976) sur les bactériocines des bactéries
a
Gram-positif et par celle d~ Korzybski 'et al. (1978) sur les anti-biotiques.les staphylocoques sont reconnus pour produi re, pl us ieurs substan-• ces anti'bactériennes incluant staphylococcines, antibiotiques et
en-zymes bactêriolytiques, sans compter que plusieurs chercheurs n'on't pas caractérisé précisément les agents inhibiteurs responsables des effets observés.
u 1
1.1 LES STAPHYLOCOCCINES
Frédéricq (1946) a été le premier
a
utiliser le terme staphy-, ,lococcine; il a remarqué que les effets inhibiteurs associés aux staphylocoques ressemblaient
a
ceux attribuables· ux col lcines'd'EBcherichia coti.
/
les staphylococcine~ sont des bactêrioc1nes produites par les
bactérie~ du genre stapnylocaccus. Il n'existe pas de définition , universellement acceptée pour ce groupe de substances, mais on
-Il ~ :
s'entend généralement pour l~ur donner les deux propriétés suivan-tes: la présence d'une portion protéique essentielle et biolegi-- ,
b
5
quement activ~~
e:
un mode d'action bactéricide (Tagg et al., 19?6)./ /
1
. 1
'I-l.
; '. 1 1 1·!
1(
1"--/
9
D'autres propd~tês peuvent ou non être p~sentes tels quolun spe'ctre d'action êtroit limité aux espèces homplog~es, un atta-chement
a
un, r~cepteur cellulaire spécifique, la' production de) '
') ,
bactêriocine déterminée génétiquement par un plasmide,
l
l,immunité de la cellule hôtea
la bactérioéine et enfin la product;on,d~ bactérjocine par biosynthèse ·létale ..Plusieurs staphylococcines ont été décrites et classifiées en se basant sur leur spectre d'inhibition (Frédéricq., 1946; Halbert et al,., 1953; Ivanov, 1970; Pulverer et Sieg, 1972;
Jetten et Vogels, 1973). Probablement à causè des difficultés rencontrées pour produire (Barrow, 1963a; Barrow, 1963p; Gagliano et Hinsdill, 1970) et ;sole~su et Wiseman, ,1967; Lachowicz,
~ 1965; parker et Simmons, 1959) plusieurs staphylococcines en gran-des quantités, peu de choses sont connues sur leur nature, leur génétique et leur mode d'action (Dajani 'et aL, 1970; lachowicz et Walczak; 1968; Moore, 1970; Rogo1sky et aL, 1976)~
Deux bactériocines produites par des souches de
s.
epide~midi 8 'ont été étudiées plus intensément. Jettel1 et aL (l972a, ,
b,c) ont travaillé sur la'staphylococcine 1580. Il s'agit d'un i complèxe l;poprotêin~-carbohydrate d'un poids molêculaire se
situant entre 150.000 et 400.000 daltons et qui peut ~tre dissocf~ en sous-unités d'environ 20,000 daltons. , la'bac~ériocine agit sur
" '
6
'.
r,
/ 1 '~.
~ ,les cellules sensibles e~ inhiban simultanément les synthêses d'acide's nucléiques et ~s~lJroté .nes. La s~~phylococcil\e inhibe
,
f..,.~~-aussi le tr~sport du glucose, ~e l'acide glutamique, des ions
4
. : ... o
rubidium et d~ lJe-nit,~heny
a-
galactoside. Quanta
L~chowiczo
"
et al., (1966, 1968), ils orlftravaï'llé sur-la staphyloccocine A.
o
Il s'a~it cette fois <t-~un polypeptide thermolabile qui possède' deux fractions biologiq~ement actives: la fra'ction 1 (34,000 dal-tonsl et" l'i1'l1act.:i;en 1 l (9,700 daltons).
.
'ç'-:2t
Des inhibit parulie autreespêce, soits. aureus,
ont été classifiés co bact~r,ocines et ont fait l'objet d'études - plus poussées. La staph~loco cine (~14) isolée par Gagliano et
"Hinsd111 (1970) apparait à 1.' état brut comme un 'complexe li popro-- téinepopro--carbqhydrate de 200,000 daltons, mais l'activité inhibitrice
semble être associée' à la partie protéique de la molécule. Ce com-plexe peut être dissocié en sous-unités de 12,500 daltons qui càn-servent leur activité. La staphylococcine 462, par ailleurs, est une lipoprotéine composée de sous-unités de 9,000 daltons qui
in-hiben~ les ~ynthêses des protéines et des acides nucléiques. Toutefois, son action sur les staphylocoques et les streptocoques
est"bact~r;ostatique au lieu de bact~ricide (Hale e~ Hinsdill, 1
1973; 1975). Tout comme les staphylococcines produites par S.
o
epidermidis , ces staphylococcines ont un effét inhibiteur sur
un grand nombre dé bàctériest ~ Gram-p~sit{f incluant S.·~U8,
7 , 1
\
s.,
, r(
1 •
t
(
(
malS pas sur les bactéries
a
Gram-négatif.Il a étê suggéré que des bactêriocines produites par
s.
au-l'BUS type phag;,quë 71, peuvent avai r un œ'rtai n rôle
a
jouer .dans des infections mixtes de la pe'au (Crowe et al.. , 1973; Dillon, 1968). Dajani etaZ. (1970) ont isolé d'un,s.
tmreUS type phagique 71,8
une substance bact~ric;de pour S. aureus et plusieurs autres
espê-1
ces à Gram-positi,f mais sans'effet sur S. epidermidis et les bac- "" ~téries â Gram-négatif. Cette subst~nce, la staphylococcine CSS, de
nature proU;;que provoque la d~gradation de l'A,RN chez les cellules
"
sensibles ainsi gu'un arrêt immédiat de la synthèse des protéines . et de l'ADN Bos causer de lyse cependant.
,
/
.1 .2 LES ANTIB
{on
qVES
t J ~ " , , , ,tt i
..
'P1~sieurs
substances inhibitricesfabriqU~
des staphy-.~
10COqUe\poss~dent
un faible poidsm01~lâ~
et sont considêrêesco~
ltant des antibiotiques.H~~
Wiseman (1967, 1971) ont isolé deux souches de S. ep~~dis produisant des substances/ /
antibiotiques nonvnées ép;'dermidines. Ces épldennidines inhibent la croissance de S. autoBUS et celle de plusieurs autres bactéries
, G
a
Gram-positif mais possèdent peu ou pas d'activité inhibitricesur
la croissance de N. gonor;hoeae.Hsu
et Wiseman (1967, 1971) ont montré que ces substances. étaient des complexes formés de deux ~lê!l1ents: l'un formant un sêdiment avec ZnC12 et l'autreo
,
/ Î '--- i , ,1",J
..
"
donnant une
frac~'~OlUble
avec le zinc. les mêmes auteurs\
(1972) ont montré que les épidermidines Al' ~2" 81 et 82 sont
des peptides de ~oids moléculaire semblable va~iant de 1,200 à .-1
1,400 daltons. Ces molécules peptidiques contiennent chacune , .
11
résidus dIacides aminés et possèdent une structure cyclique~
\,'
9
/-La mycobactocidine est un antibIotique, probablement une glyco- , / ' protéine, produit par une~souche de S. epider.rnidis (Fregnan et
S~ith,
1962) actif~ontre
les mycobactériJs. Une autre souche des.
eI?idermidis isolée de la flore de la p'eau, produit un pol eptidede faible poids moléculaire
â
spect~e largê (bactéries àram-posi-tif et
a
Gram-négatif) et possédant une activité ?nti otique (Marsh 1, 1
et ,~elwyn, 19771).
Gardner (1949) a étudié un antibiot' ue protéique thermostable isolé de S. aureuB et possédant des opriétés to~iques contre
plu-~
sieurs espêces bactériepnes. Dlautres antibiotiques d'origine staphy- Jlococcique ont été décrit par Jones et Edwards (1966) et Loeb et
al. (1950), mais c
"",f'"
/ '
inh'ibitiurs n lont pas ét~ bien caract@risés.~ ,
6 -"
/
~T~ ~ ~
/
;:~
.. /.HS ENZYMES BAClER IOL Y 7, / / / Dans l~~r c1assificatfon des produits inhibiteurs.synthHisés / " , / par' les stap 1
l~çoques,
Jetten et Vogels (1973) ont réuni dans le//~.- t ~
,/ grout>e A, les substances causant 1 a lyse des cell u1es et hydrolysant
Cl •
\
. l
!
1J
j 1\
, , -, " ,~:
1 ... 4f. ~ 1 , ) l, ! ' l. !
i
i ! \ \ , -----
- ---_._---
--,.
.
•
•
(
la paroi cellulaire. Ils ont d~fini ces substances comme des
enzymes lytiques. Schindler et $chuhardt (1964)oo~t observê
' r
qu'une souche de S. staphyZoZytiaue pr.oduisait un facteur
anti-"
bactérien qui lysait aussi bien les cellules vivantes que les
cellules mortes de S. aure,U8. Ce facteur a été appelé lysosta::. __
phine et semble être une peQtidase de poids moléculaire de 32,000
daltOns qui sem!?1e avoir une activité spécifique contre les ponts
.
'polyglicine des paTois cellulaires des staphylocOques.)-a lysozyme'
(endo,-~-N-acétylmuramldase) produite par S. œœeU8 a êtê dêcrite
par Hawiger (1968). Il s'agi-t--,d'une glycosidase qui agit sur les
.
,
cellules de MicrocOCCU8 l.,ysodeikticus et qui est aussi capable de
dissoudre la couche glycopeptidique de la paroi cellulaire de
plu-"-..
sieurs bactéries.
Une autre enzyme lytique produite par .S. aureus a ;Hê décrite
par Wadstrljm et Hisatsune (1970). Cette enzyme esC une
endo-(3-N-. 0
acétylglucosaminidase dont le poids moléculalre s~ situe autour de
70,0Q(j' daltons et- qui possède une activit~ lytique sur les cellules
de M .. tysodeikticu8.
L'hémolysine delta de S. aureuB ,q'ui 'lyse les globules rouges,
est reconnue aussi pour posséder une aGtivitê' antibiotique sur les ,
'
'bactéries
a
Gram-positif'(Hoffm~n et Streitfe,ld,,1965). L'agentl~iique
appelévi~olysine
par Ralston et al. (1955) etprod~it
par"
'\
, ! • j 1 1 10~,
~
. .S. aupeu8 ~git sur les ce~lules des staphylocoques, mais
seule-J
ment après altération de la paroi cellulaire ou apr,ês "sensibi-Hsation" des cellules par des bactériophages. Un agent lytique semblable et appelé CWSE-80 a Hê-décrit par Doughty et Mann , ,
~
(1967), tandis que' l'enzyme lytique ,nol1llJlée "staphy1ococcal phage-associated lysin" (PA~) par Sonstein et aL. (1971) possède un~
activité lytique contre plusieurs souches de S. ~UB "mais non contre d'autres espèces bactériennes. Cette enzyme produite~
S. aureu8 PS53 est une peptidas"e.
1
1.4 'AUTRERibble (1967) a montré que les staphylocoques
coagulase-né-«>
\ ,
gatifs inhibent les staphylocoques coagulase-positifs par un inhibiteur the'nnolabile du métabolisme de 1a nicotinamide.
1 D'autres auteurs ont décrit des activités inhibitrices produi-tes par les staphylocoques mais les substances impliquées dans de
-telles activités n~ont pas, été c1Lairement dHinies (Tagg et al, .... ,
. "
1976; Korzybski et aZ" 1978),
2.
INHIBITIONMICROBIENNE
DE N"gonor-rnoeaeCette sectionfe la revue de la littérature s'est largement ins-pirée de l'excellente ,revue' publiée par Ingraham et a~. (1978).
(
"
2.1 INHIBITION IN VITRO DE N. gonorrhoeaè
En 1967; Nartin et aZ. ont détecté deux fois pl us de cul
tu-res de gonocoques t provenant de prél èvements gén; taux, lorsque
1~ croissance des autres mi~roorganismes était supprimée par l'additidn au milieu d'antibiotiques sélectifs. Depuis cette ob-servation
" il a été d~montr~ que p1usi,eurs microorganismes norma-lement présents chez 1 1 homme ~ont des inhibitéurs fÏn vitrb du
go-nocoque (Tableau 1). p'armi ces inhi biteurs, on retr9uve de nom-., breux genres t des bactéries aérobies et ana~robies ai nsi que des
bactéries ~ Gram-pDsi tif et à Gram-négati f.
l i inhibiti on gonococcique a' été habituellement démontrée sur
~
mi1ieu solide par croissance séquentielle de la souche productrice (i.e. l'organisme soupçonné de produire une activité inhibitrice)
,
et de 11 indicateur (Le. une souche de N. gonol'l'hoeae ). La mé-thode "basal streak/cross streak" est un exemple de cette appro-che avec laquelle la souappro-che productrice et l'indicateur sont ense-menc~s ~ angle droit. En variant cette te-chnique, on obtient la 'm~thode "flip"-flop" qui consiste
a
inverser soigneusement lagélo-se sur laquelle la souche productrice s'est développée et
a
ense-mencer l'indicateur sur l'autre surface non utilisée de la gélose.Ces techniques peuvent être modi fiées de sorte 'llue l a souche
,
-productrice et' l'indicateur puissent cro'itre simultanément au lieu 12 1
(
•
(
TABLEAU 1: MIC~OORGANISMES INHIBANT LA CROISSANCE DE
N. gonoIT~oeae IN VITRO
..
,.
GENRE DE LA SOUCHE INHIBITRICE Aoi'fBtobactero Aer'omona~ Bacteroides Bifidobacte:r:>iwn Candida Citrobacter...
ctostridiwh . CorynebactePium Ente:robacter Eecherichia , Eubacteroiwn Fueôbactel'iwn Gaffkya Kleot)ie l la Lactobaci Hus Mic:rococcus Neisseria PeptococCU8 Peptostroeptococcus Propionibàcteroiwn Prot:eU8 Pseudomonas stapnytocoqCU8 \ streptococCU8 Se:r:ratia Vibno • REFERENCE ,'. Bisaill,on ei al. (1980) Geizer, (1968). MOrin et al. (1980b) - Kaye et al. (-1977) JJ
Bisaillon.et al.(1980);Hipp et al.(l974);Hipp et
al.(1975);Kayeetal.(1977). " .
Kraus et al. (1976) Kaye et
at.
(1977)Kraus (1975):Saigh et aZ.(1978);Shtibel (1976) .•
Bisai,llon et al.. (1'980); Kraus et al. (1976) .
Bisaillon et al..(1980);Gc;!izer (1968);Kraus (1975); Simpson et a'l. (1979) .
Mori n et aZ. (l980b
r ""
Kaye et ~77)Kaye -et aZ;(1917)
Bisai110n et aZ.~1980)
Morin et al.(1980a);Sâigh et al.(197B).
Bisaillon et al..(19'80)
Flynn .et al.(1972};Ingraham et al.(1978); Lawton
et al.(1976);Shtibe] (1976). •
Mo ri n et aZ. ( 1980b)
Morin et aZ.(l980b)
Mo ri n et al. (1980b) Kraus (1977) .
Geizer (1968) ;'Kraus et al.(1976); Morse et at.
(1976h Sidberry et al.(l977).
Bisai110n et al.(1980);Hsu et al.(1967);Ingraham
et aZ.(1978);Kaye et ~Z.(1977)jKraus et aZ.(l974);
Saigh et al. (1978) ;Shtibel (1976).
J ,
Bisaillon et çzl.(1980};Oajan'i et aZ.(1976);Kaye
et aZ.(1977) ;Kraus (l975);McBride et al.{19BO');
Morin et al. {1980b};Saigh et a3.(l978};$htibel
-, (1976). , Kraus (977) Ge1zer (196'8) ? • ,
.
13 ~ , , , 1 ..! , ,(
..
..t
1
d.e séquenti ~11ement. Toutefoi s, ces tec~niques requi èrent l' u- . "ti1isation d'un milieU contenant de l'agar et permettant la (,
"
croissance des deux microorganismes.
D'autres techniques uti1i·sent une seconde couche d'agar, laissant suggéret~que lïinhibition est due
a
la production de..
substances diffusibles pl~tôt qll.'A une compétition nutritive. Ces méthodes permettent aussi de tester des espèces productri ces (ex. anaérobies) exigeant des conditions de ctoissance-très dif-. férentes de celles' du gonocoquedif-.
,
Différents auteurs ont utilisé une ou plusieurs de ces
métho
-1 des pour démontrer l'inhibition in vitro de N. gonorrhoeae par
des mi croorganismes. ~'i sail1'~>n et al. (1980) ont montré que "
ac-..
t1vitê inhibitrice qe la souche productrice pouvait varier avec". la méthode utilisée pour mettre en évidence l'interférence. En , , 1974, Kraus et Ellis'on ont montré que deux souches de S.
epider-midi~ pouvaient inhiber la croiss.ance de N. gonorrhoeae in vitro
Ces deux souches avaient été isolées de l'urètre de deux homnes qui
, .
n'avaient pas développé d'infection gonococcique malgré des
expo-,
sitions répétées au germe. Kraus et Ellison (1974) ont montré
éga-4 ~
1ement què 39 des 75 honmes sains étudiés, possédaient dans leurs . 'urêtres, des souche's de S. epidermidis ayant la capac;U
d'inter-"
férer avec 1 a croi ssan.ce de N. gonorrhoeae i.n vitro.
14
.1.
t
,
(
( ; ;t .:\
En 1976, 'Sh,tibef a obseiié'que la croissance de certaines cultures deo·N. gonorrhoeae ~tait inhibée en} présence de di vers contaminants bactériens provenant de prélêvements urogénitaux. , Dans cette étude, l~s' esp~ces bâcté~iennes qui ont -inhib~ l'a croissance de N. gono!'moeae ont ~té par ordre de fréquen~e
,
N. meningitidis ~ S. epidermidis, Corynebacte!'ium s P ." S. aul'eU8"
streptococaus du groupe A et N. subf2ava. Shtibel (1976) a
, ,
suggéré que -cette inhibition était causée p'ar des bactériocines.
/1
Tou~ours en 1976, Kraus e~ontré que plusteu~~ bac- ) tér;es aérobies ou anaérobies facultatives frêquemment iden'tifÙ!eS dans les laboratoires cliniques, incluant S. au:I'eU8 et S. epid.er-;.
f
midis, inhibaient )a- croissance du gonocoque in vit1'o. Ils ont
aussi démontré que l 1 interférence gonococcique n'était pas un
phé-nomêne de 'tout ou rien mais variait
d~ l'inhibit~on
deqUél~ues
souche~
a
l'inhibition de plusieurs. Leur étude ainsi que celle de 8isaillon et aL (1980),;()nt indiqué que la capacité inhibitricedés souches n'êtait pas reliée au site anatomique de l'isolat; le pourcentage de f~rts·inhibiteurs parmi; les staphYlocoques a êt~
similaire pour différentes régions. Le m~canisme impl.iqué dans llinhihitifn invitro es.t inconnu et ne peut être dêterm;né de leurs donn~s. ". 15 j -1
j
,
f 11.
! 1 1 , ll
"" (
.'
(
J'
.
Kaye e~ Levisont en 1977, ont observ~ que les staphylocoques
sont les microorganismes, isolés des s~crHions vaginales :èt
cer-.. t ./' ~
vicales, qui interfêrent le plu's souvent avec le gonocoque. SaighO
~.
et aL (1978) et Ingraham et aZ. (1978), dans des étuçes similaires,
sur les bactéries aérob~es et anaérobieg facultati ves de 1 a flore gênita·le, ont troùvê que les staphylocoques Haient aussi parmi les plus fréquents inhibiteurs du gonocoque. Bisaillon et aL. (1980) ont montré que les st'aphy ocoques COagUlaSe-nêga~S 'représentaient la majorité des isol~ts uro énitaux inhibiteurs tu' ils ont étudiés . . ~ De pl,us, Bisai'llon et aL 1981) ont Illontré qué pl usieurs de ces
st~phylocoques possédaient à la fois un large spectre antigonococci-que et un spectre d' interférence étroit envers des bactéries
aéro-r'
btes et anaérobies représentant la flore urogénitale normale ..
"
2.2 SUBSTANCES ANTIGONOCOCCIQUES D'ORIGINE MICROBIENNE
'L'étûde des acti'vités antigonôcocciques a I!!té campl iquée par la difficulté d'ob,tenir ces ,activités sous forme soluble. Les
,)'
16
• l ,~
'''---études' de Dajani
et
aL.
(1976) sur les vi'ridil)es de plusieurs strep-'),~-e
.tocoques<a,lpha-Mmolytiquesont montré que malgr~ la faèilit~ avec "
~
laquellé on peut dêmontrer.la p"rêsence d'inhib'iteurs gonococ~iques
, .
diffusibles en milieu solide,' aucune substance inhibitrice so'luble n'a pu ~tre démontrée en milieu liquide. Par contre, l'homogênêi-sation -de cellules entières de St1'eptooocaus mitis a résultê en
.
, \ 0' i1
,/
1..
(
l'obtention d'un inhibiteur de N. gonoI'l'hoeae , la viridine B.
,
Cette bactériocine possède quelques propriêtês inha,bituelles telles qu'une thenno'labilit~, une activité ayant un êcart de pH êtroit et une absence de capacité d'absorption aux cellules sen-sibles .
..J Gei_zer (l968)
a
rapportê une inhibition,.,de la croissance du, )
.
gonocoque par de~ substances non identifiées et produites par
1
" v
de s _souches de Vibrio chotel'ae ~ E. co "li J ,tleI'omonaa hydl'ophy la,
y
Miaroaocaus sp. et Paeudomonas aerruginosa. Mc1JS"e et at. (19!6) ont isolé", caract~risê et identifiê une pyocine produite par
P. aerruginosa qui inhibe N. gonol'rhoeae. Hipp et al. (l974) ont démontré que Candida albioans produit une substance sol uble inhib,itri cè pour N. gonol'l'haeae. Pl us récenment, Simpson et Davis (1979) ont noté qu'une souche de E. coU pouvait empêcher la croissance de N. gonol'l'hoeae in v"Î,tl'O par la production d'~ne
substance inhibitrice bactériostatique, diffusible dans "agçr et pyant un poids
molé~ulaire
entre 1,200 et 2,000 daltons.--
..
" ~es travaux de Flynn et McEntegart (1972) sur l'inhibition
"
ihtra-espêces parmi les souches de N. gonoI'I'hoeae ont sugg~rê, que cet organisme est capable d'êlaborer des gonococcines. Walstad
et al. (l974) ont dêmontri'i une inhibition de la èroissance de"N.
, "
gOr/orrhoeae par certaines souches de gonocoque due
a
1 a producti'on\
17·
i ·
,1
;! . ,
(
1
(
-.
,.
\d'acides gras libres et de lysophosphatidyl~thanolam;ne, mais pas
a
de vêrHables bactériocines. Knapp et al,. (1975) ont aussi trouvé' 'une" substance inhi bi·tri ce non spécifique produite par N.(lonoX'X'hoe~e. Lawton'et al. ont mis en évidence pour la premiêre
,8.
fois en 1976 la production de bactêriocines typiques pail' N. (10--,
norrhoeae.
Miller et al" (1977) ont étudié les effets individuels des aci des gras sur 1 a cro; ssance, l'ut; 1 i sati on d' oxygêne et 1 a rêduc-, co
ti on de l' act; vitê ni cotinami de, adênfne dinucl êoti dE! (NAD) d~ N. 1
(10-norrhbeae. Il sont t rouv~ qu' une va ri ~té de compos és il cou r-tes chaînés'(Cl-C6) et'à longues,chatnes (C16-C20) inhibaient la croissance du gonocoque. De même, les études d',Ingraham et 1 aL
18
(1978) sur l'inhibition gonococti-que par N. meningitidis ont démon-~
tré que' les compos~s lipidiquesl solubles sont aussi des substances inhib;itrices. En effet, tout microorganisme nbérant de grandes
Q
quantités d'acides gras durant sa croissance,.peut inhiber le go-1
nocoque. Cependant, ces 'effets peuvent être contre,carrés par l' ad-dition d'amidon dans le milieu supportant la croissance de N. (/0-norrhoeae (Ley et a~., 1946) •
•
Les expêri ences dl l ngraham et aL (1978) avec S. epide:rmidie
.,
. interfêrant ave,c la croissance de N. gonol'rhoeae ont rê'v~n~ des
'"
difficul tés pour isoler d~s substances inhibitrices solubles. La
t,
·1
Q }
·
(
production d'inhibiteurs en milieu liquide a ét~ un ~chec, mais
'4:
~ IJ ~_
ils ont détecté de l'activité anUgonococcique soluble dan9s le
surnageant obtenu aprÈ!s centrifugation de milieux gélos~s qui
, î
~vaient supporté la croissance des staphylocoques. Ces rêsuJtats' '. .sont similaires
a
ceux de Jetten etaZ..{1972c) qui ont étéinca-o
pables d'obtenir avec des cultures liquides suffisamment de sta-phylococcine 1580 pour les étapes de pur'fication subséquentes. Ingraham et al. (1978) n'ont pas identifH! la substance acti~e
mais ils ont mentionné la possibilité qu'il s'agisse d'~ne staphy-lococcine similaire
a
celle êtudiée par Jetten et al. (1972c).Morriss et al,. (197,8) ont produit en milieu liquide une sta':'
. /
phyl ococei ne 5Bac Rl) ayant un ,~effet bactéri.ci de sur des souche's de gonocoques résistantes et sensibles
a
la pénicilline. Donc peu de substances antigonococciques d'origine microbienne, e't plus par-ticul ièrement celles prodJ,Jites patr"des microorganismes de la floreurogén~tale, ont Hé caractérisées et puri fiées, et les mécani smes d'aCtion de ces substjlnces deme~nt i!lconnus:
2.3 INHIBITION IN VIVO DE N. gonoIThoeae .
.
' ,La signifi'cation clinique de l'interf~rence gonococcique
dé-montr~e irt..,vitro est discutable sans études in vivo adéquates et
"
comp l émenta i re~ .
..
/~ ,
.
(
/ //
/ / 1 1 / 1 1L'implantation de chambres sous-cutan~es,chez le cobaye, permettan(la croissance simultanée de soùches productrices et
/ . .
de gonocoques/~i rul ents, a ~té ut il; sée pour êtudi er l' fnter-' ,
férence bJctêrienne in vivo. En uti1;'sant ce modêle, Kraus et'
al,. (19]f5) ont démontré que l'inhibition de la-croissance dè N.
gono rhoeae par d'autres bactéries, dont
s.
e)i.derm-ldis ets.
us pouvait survenir aussi in vivo. Ils ont montré aussi que,
/ quelques microorganismes t'rouvés non i"hibiteurs in vitro sont devenus inhibiteurs in'vivo. De même, il s ont démontré que l' i
n-~ 1
troduction d'une souche fortement t-nhibitrice dans la chambre sous-cutanée résultait en une élimination rapide du gonocoque et
.
prévenait l'infection avec N. gononohoeae. Enfin, ils ont mis en
évidence que l'interfére7 bactérienne avec la croissance du gonocoque variait e~nction de i) la capacité inhibitrice des bactéries impliquftés; ii} le nombre de bactéries inhibitrices
/
.
présentes; iirJ le nombre,de gonocoques , pr~sents et iv) la durée de " i
nter1i~ti
on mi crobienne . 1 1 , . / 1/ Evi demment, l a chambre sous-cutanée représente 'un envi
ronne-/
'/fuent bien-différent de celui du tractus urogénital, la 00 les \ , / interactions entre la flore indigêne et le 'gonocoque envahisseur
surviennent. Le modêle de Ward . , et a7" (1974) utilisant des trompes de fallope humaines, serait probablement un modè·le' plus appropriê pour êtudier 1linterfêrence' gonococcique.
1 20 ,1 r r.i'l
.
jf
,~
J(
..
( " ",.
Le r61e de l'inhibition micro~iennè dans la pathogênêse des infections gonococciques.chez l'homme n'est pas connu; cependa t
1
les travaux de Volk et Kraus (1973), Kraus et E1Hson (1974)
et"
" '
Saigh et al,. .(1978) s'uggêrent que " interférence Qactérien e peut conférer une certaine protecti on ,contre la ~onorrhée .
"
, le ou les
ml!ca~;sm.s. ;l11Pl'iqU~S
dans;'jnter'fê~nce
ilvtt'ro'
aussi "bien qu'in vivo avec N.~oYlOl'l'hoeae
sont iÎ'lconnus. Lei mécanismes~euvent
varier avec chaque espèce bactêrienne e.t " i,rl'hibition peuti~-1
/
pliquer plus d'un mécanisme (McCabe,1967). L'inMbition peut être, due
a
l'ac~dification d~
milieu,a
desproduitsflo~iques,'a
des~nzy-21
1
..
w mes bactériolytiques, des antagonistes métaboliques, des
ba~rioCines
ou
a
la combinaison de pl usieurs de ces facteurs(Ingraha~t
aZ., '1978). Ce travail a été entrepris afin d'étudier les activités antigono- l'cocciques produites in vitl'O par des staphylocoques coagulase-oêgatifs
isol~s de 'la 'flore urog~nitale. Le but de cette ~tlJde a êtê de trou-- ~ ver un isolat èt une m~thode de production permettant d'obtenir une
, \
activité antigonococcique élevée, pour éventuellement purifier et
ca-,
ractériser l'inhiBiteur produit.
1 ·f " '
'/',
l , y
".
1 l' , j / j/ / )/
(
.
,•
, , f .' ( "j
l
1 l1
, ~ .: ,.
1,
) 1 ~ 1 ,; 11
(
(, ( L 1 1 • - SOUCHES BACTERIENNES1.1
I~OLATSUROGENITAUX
Les soucpes utilisées ont Hê isolées de la flore urogéni.tale nonnale et sélectionn~es pour leur spe.ctre antigonococcique large
.
(Bisaillon et aL, 1980; 1981). Les 24 souches de staphylocoques
0(
coagulase~négatifs reten~es
pour cette étude ont aussi montréuin
spectre an~ibactérien étroit (Bisai11ori et al., 1981) et elles ont été numérotées de 1~ façon suivante: 4,5,7,10,26,27,28,30,32,33,
'-36,37,46,48,?0,51 ,53,55,56,62,63,66,68 et 72. Laosouche 61, un staphylocoque coagulase-positif et la souche 12, un streptocoque' , du groupe D ont aussi
ét~
util isées dans éertaines expériences '~~-1 .2 ,-NEISSEBIA GONORPJ1OEAEtt
L'a sou'che G-10 de N. gonorrhoeae a été utilisie comme cible car cette souche est 11 une des pl us sensibles que nous possédi ons (Bisaillon et aZ., 1980). La souche PR77112, productrice de péni-' cillinase, a été util;~ée dans quelques expériences (Bisaillon et al., 1981).
1.3 ,CONSERVATI:ON
Les isolats urogénitaux ainsi que les souches de N.
gonor-", ~ ~
rhoeae, ont {tê conser~s
a
-76°C sous forme 'de suspensions dansun milieu "Brain Heart Infusion" (BHI, 01fco) enr1cryi de 10% (v/v)
l, 1 ; , 0 , , ,
!
-, ,1
-.
, ' ," " 7 (
...
-! • 1 > ;1 d~glyc~rol. ,>les suspensions ont été prépar~es à l'aide de la masse bac"
Il là ' ,
térienne r~cfJpérée de cultures de 24 heures sur 'géloses',Columbia
, , " , .
contenant 7.5% (v/v) de s~ng de mouton (GS, ~nst;tut
Armand-Frap-"~
pier Production (IAFP)), pour les i~olats urog~nitayx et sur
gélo-Q / "
ses GC, (Gibco) enrichies de 0.1% (v/v) CVA (Gibco) et de 1.5% (v/v)
de sang de cheval hémolysé (GC1CV~BA) pour l'es gonocoques.
Certains isolats urogénitaux (souches 5,7,32,37 et 66) ainsi que les 'souches de N. gonorrhoeae ont Hé également conservés sous
Cil forme lyophilisée à 4°C. Ces lyophilisations ont ~té èffectu~es ~\ , l'a~ d'un appareH Virtis (modêle lO-147-MR-BA) et en présence
d'un
e~iPient
soit 2% (p/v) de lait~crémé
(Difco). ' 1.4 PRECULTURES
-Pour chaque exp~rience, une nouvelle suspension .des isolats
. !
urogéniJ;aux conservée.
a
-76°C a été décongelée, pui s O.f
ml de ., f
cette derniêre a été ensemencé' sur une gélos&GS et incubé 24 heu-res à 37°C. Un second passage sur milieu solJde ou liquide appro-prié a été effectué avant que ces microorganismes ne soient utili"
/'
·sés dans une expérience. la période d'incubation de ces précultures,
r
a.été d',environ 18 heures à 37°C.
(
p
(
Aussi, chaque semaine une nouvelle suspension de la souche
de N. gonorrhoeae dêsir~e, conservée à -76°C, a ét~ d~congelée
.
pui s ensemencée sur des géloses GC{VABA. La souche a été
mai.nte-\
nue par repiquages quotidiens non sélectifs sur milieu GC1CVABA et
inçubé~à
37°C enpr~sènce d~
S(de CO2 et de 70% d'humidité.'v h
Pour fin d'expérience, les souches de N. gonorrhoeae n'ont jamais été utilisées avant qu'il y ait eu un deuxième passage sur milieu
-GCrÇy~~A et la période d'incubation de ces précultures a été d'en-viron 18 heures à 37°C.
1.5 STANDARDISA1ION DES SUSPENSIONS
Des suspensions ont Hé pr~parées â l'aide de la masse bacté-rienne récupérée ;de précultures de N. gonorrhoeae sur géloses GC1CVABA ou 'de précultures des isolats urogénitaux sur géloses GS. Les suspen'sions, gonococciques ont été effectuées en milieu PBS (tampon phosphate 50 mM, pH 7.0; NaC1 140 mM) contenant du MgS04 2 mM et du CaC1
2 2 mM, tandis qùeo les suspens:ions des isolats
uro-,
25
g~nitaux ont été effectu~es dans de 1 ~eau physiologique (0.85%
(plv)
, NaC1). la deun$'it~ optique des suspensions gonococciques et des sus-pensions des isolats urogénitaux a ét~ ajustée à une valeur de 0.15 (1
x
10~
Unités Formatrices de Colonies (U.F.C.)/ml) à 540 nm avec un spectrophotomêtre Bausch et Lomb, Spectronic'21 (Saheb et al.,1979).- Pour la méthode ,de dêtection des unit~s arbitraires, la den-sitê opti~ue , des suspensions gonococciques a été ajustée,
a
une valeur de 1.5a
540 nm (1 x 109U.F.C./ml)~
...f \
,
î .'
(
2. MILIEUX DE CULTURE
/ (
Plusieurs milieux solides, semi-solides et/liquides ont été ùti-lisés pour la production d'activités antigonococciques. les diffé-rents mi,lieux ont Hé préparés avec"de l'eau distillée et le"s milieux contenant de l'agar ont été d'abord chauffés
a
ébullition ou jusqu'à., ,
ce que l'agar soit complètement dissous". Tous les milieux ont alors
/
été stérilisés
a
~autoclave A. une température de 121°C pendant 15 1•
minute$ (tel que recommandé par le fabriquant).
(
les suppléments (supplémen~ défini de Lankford et supplément CVA) ont été stérilisés par filtration à travers une membrane ayant une porosité de 0.20 ~m (Nalge Co., Rochester, N.Y.), et ils ont été
ajou
-tés stérilement aux milieux déjà stérllisés dont la température avait atteint envi rDn 50D
e.·
2.1 MILIEUX SOLIDES
.' Les différents m;l'ieux solides Ol)t été répartis, après stl~ri
lisation, dans des baltes de Pétri (15 x 100 mn) à raison de 25 ml'
par ba~te. Les 'milieux utllisés ont été les suivants:
Le.milieu
BHI,A
qui contenalt par litre d'eau distillée: Bacto-"Brain He'art Infusi on", 37 9 (Difco, lot #6p6140) Bacto-agar (Difco) 10 9 : ' If 1.
, 1 26(
Le milieu BHI
2A qui eonteriait par~ litre d'e~u distillée: Bacto-"Brain Heart Infusion". 37 9
(Oifco, lot #668294)
Bacto-agar (Difeo)
" .if
10 9
Le milie~ BHI~A qui contenait par litre d'eau distillée: ., "Brain Heart In~usion Broth~" 38 9
(Gibco, lot #870093)
Baeto-agar (Difço) 10 9
Le milieu OSA ("Dextrose Starch.Agar") qui contenait par litre d'eau distillée:
Prot~ose peptone no. 3 (Difeo) 15 9
Baeto-dextrose (Oifeo) 2 9
Amidon soluble (Difco) 10 9 _
Chlorure de sodium 5 9
Phèsphate de sodium dibasique 3 9
.
Bacto-gélatine (Oifco) 20 9
Bacto-agar (Difeo) 10 9
Le milieu pC,CVAA qui contenait par litre d1eau distillée: Base Ge (Gi beo, lot #970435A 36, 9
contenant 10 9/1 dl aga r)
Suppl~ment CV A' (Gibco)
-.
l ml
" ,
(
Le.milieu GC1CVABA ~tait un milieu GC1CVAA enrichi de 1.5% (v/v) de sang de cheval Mmolysé (IAFP).
Le milieu GC1DS~ qui contenait par litre d'eau distillée: r
Base
te
(Gibco, lot #970435A, 36 9 contenant 10 9/1 d'agarfSuppl~ment défini de Lankford 20 ml (tankford, 1950)
_~~.!"iJieu GC
2DSA qui contenait par litre d'eau distillée: Base Ge (BBl, lot #A2Dl RB , 36 9
contenant 10 9/1 d'agar)
Suppl~ment défini ,de Lankford 20 ml
Le milieu TSA qui contenait par litre d'eau distillée:
Baeto-"Tryptie" Soy
B1h"
(DHeo) 30 9Bacto-agar (Ili feo) 10 9
2.2 MILIEUX SEMI-SOLIDES
28
Les milieux semi-solides ~ti1isês ont été les s~ivants: BHIl' BHI2 et BHI3, tels que défipis précédemment et avec la différence qu'ils ne contenaient que 0.4% (g/l) d'agar. Les milieux ont été
répart~s; aprês 'stérilisation, dans des bottes de Pétri (15 x 100 mm)
a
raison de 20a
25 ml par boJte.'i . . -- -- -- -- / . )
1
J 1 ! '1 l(
.
2.3 MILIEUX LIQUIDES l'
Les milieux liquides utilisés ont été les suivants: BHI 1,
., ' 1
,BHI 2, BHI3',TSB, GC\CVAB et GC1DS, :els que définis précédemment, et avec la différence qu'ils ne contenaient pas d'agar. Les milieux ont ~t~ répartis
a
raison de 100 ml de milieu par erlen-meyer de 500 ml, avant d'être stérilisés.3. DEMONSTRATION DE L!ACTIVITE ANTIGONOCOCCIQUE SUR MIlIEUX SOLIDES
•
3.1 METHODE DIRECTE
Cette méthode est basée sur celle développée par Gratia (1946) et modifiée par Marsh et Selwyn (197'7). Les isolats urogénitaux
o '
a
tester ont été ensemencés, sous forme de "spot" d'environ 1.5 cm de diamètre, sur des mil ieux GC1·CVABA préalablement inoculés avec 0.1 ml de différentes dilutions décimales {1:l0, 1:100} provenant
29
d'une susRension standardisée de N. gono~rhoeae, de sorte
a
couvrir toute)la surface des géloses."
>
Les souches productrices et cibles ont été incub~es pour une période de 18
a
24 heuresa
37°C dans une atmosphère de 5% de CO2
i
et de 70% d'humidité, de façon
a
croitre simultanément. ,L'inter-férence bactérienne a été mise en évidence par la présence d'une zone d'inhibition du gonocoque autour de l'isolat urogénital. Le prérequis était que la ,croissance de 1 'isolat urogénital soit,


![FIGURE 3: CINETIQUE DE PRODUCTION DE L'ACTIVITE ANTI- b GONOCOCCIQUE DU STAPHYLOCOQUE COAGULASE-NEGATIF 32, EN MILIEU SHI] LIQUIDE](https://thumb-eu.123doks.com/thumbv2/123doknet/7656148.238006/77.918.72.907.26.1152/cinetique-production-activite-gonococcique-staphylocoque-coagulase-negatif-liquide.webp)


