Induction et stabilisation de protéines endogènes
chez la pomme de terre exprimant une cystatine
recombinante
Mémoire
Aurélie Munger
Maîtrise en biologie végétale Maître ès sciences (M.Sc.)
Québec, Canada
Induction et stabilisation de protéines endogènes
chez la pomme de terre exprimant une cystatine
recombinante
Mémoire
Aurélie Munger
Sous la direction de:
Dominique Michaud, directeur de recherche Russell Tweddell, codirecteur de recherche
Résumé
Des cystatines recombinantes employées en phytoprotection dans des variétés végétales trans-géniques ont été étudiées pour les effets d’interférence métabolique qu’elles causaient dans la plante hôte. L’objectif de ce projet était d’évaluer ces effets pléiotropiques à l’aide de
li-gnées de pommes de terre (Solanum tuberosum) exprimant des cystatines de céréales de façon
constitutive dans l’ensemble des tissus de la plante. Nos résultats ont montré que la cystatine du maïs CCII induit, dans les feuilles, une activation de protéines endogènes reliées au stress,
conférant notamment un effet protecteur contre le champignonBotrytis cinerea. La cystatine
du riz OCI, exprimée dans les tubercules transgéniques, a engendré un délai de leur germi-nation et une préservation des principaux nutriments en période d’entreposage. En somme, nos lignées de pommes de terre exprimant une cystatine recombinante démontrent des effets d’interférence métabolique à valeur agronomique éventuellement intéressante.
Table des matières
Résumé iii
Table des matières iv
Liste des tableaux vi
Liste des figures vii
Liste des abréviations viii
Remerciements xi
Avant-propos xii
Introduction 1
Mise en contexte générale . . . 1
Les cystatines en phytoprotection . . . 3
Les inhibiteurs de protéases . . . 3
Le cas de la pomme de terre, Solanum tuberosum . . . 13
Hypothèses et objectifs du projet . . . 15
1 Effets bénéfiques inattendus d’une cystatine de céréale dans des lignées transgéniques de pomme de terre, Solanum tuberosum 19 1.1 Résumé . . . 19
1.2 Abstract . . . 21
1.3 Background . . . 21
1.4 Results and discussion . . . 23
1.5 Conclusion . . . 32
1.6 Methods . . . 33
1.7 Bibliography . . . 37
2 Préservation des nutriments et inhibition transitoire de la germination dans des tubercules de pomme de terre exprimant des cystatines de céréales 43 2.1 Résumé . . . 43
2.2 Abstract . . . 45
2.3 Background . . . 46
2.4 Results and discussion . . . 47
2.6 Methods . . . 55
2.7 Bibliography . . . 58
Discussion et conclusion 65
Retour sur les Objectifs 1 à 3 . . . 66
Retour sur les Objectifs 4 à 6 . . . 68
Conclusions et perspectives . . . 70
Bibliographie 72
Liste des tableaux
1.1 Proteins up- or downregulated in CCII-expressing potato lines 9.4 and 10.4 . . 27
1.2 Inhibitory effect of diagnostic protease inhibitors on Botrytis cinerea secreted
proteases . . . 29
2.1 Starch and soluble sugars in OCI-expressing potato tubers stored at 4˚C for
0, 36 or 48 weeks . . . 52
A.1 LC-MSMS identification of leaf proteins up- or downregulated in CCII-expressing
Liste des figures
0.1 Représentation schématique des types d’inhibition enzymatique . . . 5
0.2 Représentation structurale d’une cystatine végétale, la cystatine de maïs CCII 7
0.3 Représentation schématique de l’effet inhibiteur d’une cystatine dans un
sys-tème plante-insecte. . . 11
0.4 Schématisation de l’hypothèse de travail . . . 17
1.1 Expression of CCII and potato multicystatin in leaves of transgenic potato lines 24
1.2 Expression of endogenous defense proteins in leaves of control line K and
CCII-expressing lines 4.1c, 9.4 and 10.4. . . 25
1.3 2-DE analysis of the leaf proteome in control line K and CCII-expressing lines
9.4 and 10.4 . . . 26
1.4 Secreted proteases of the fungal necrotroph Botrytis cinerea . . . 30
1.5 Impact of recombinant CCII and CCII-expressing potato lines on growth and
infection capacity ofBotrytis cinerea . . . 31
2.1 OCI expression in leaves and tubers of line K52, line K53 and control line K. . 48
2.2 Number, length and distribution of buds and sprouts on tubers of lines K, K52
and K53 stored for 48 weeks at 4°C. . . 49
2.3 Soluble proteins, potato multicystatin (PMC) and protease activity in tubers
of line K52, line K53 and control line K stored for different periods at 4°C . . . 50
2.4 Growth parameters of plants emerged from line K or line K52 tubers stored for
48 weeks at 4°C. . . 53
2.5 Relative amounts of patatin, PMC and Ser protease inhibitors (PI’s) . . . 54
3.1 Représentation visuelle des analyses effectuées sur le tubercule en fonction de
Liste des abréviations
2-DE Two-dimensional gel electrophoresis
Asp Aspartate
BSA Bovine serum albumin
Caspase Cysteinyl-aspartate-cleaving proteases
CCI Corn cystatin I
CCII Corn cystatin II
Cys Cystéine
DTT Dithiothréitol
E-64 l-trans-Epoxysuccinyl-leucylamido(4-guanidino)butane
IEF Isoelectric focusing
IPs Inhibiteurs de protéases
MCP Mort cellulaire programmée
OCI Oryzacystatine I
PAGE Polyacrylamide gel electrophoresis PCR Polymerase chain reaction
PMC Potato multicystatin
PMSF Phenylmethylsulfonyl fluoride PR-2 (PR-3) proteins Pathogenesis-related proteins 2 (3)
PVPP Polyvinyl pyrrolidone
RT PCR Reverse transcriptase PCR SDS Sodium dodecyl sulfate
SELDI TOF MS Surface-enhanced laser desorption ionization time-of-flight mass spectrometry
L’expérience ne se trompe jamais, ce sont nos jugements qui se trompent.
Remerciements
Après toutes ces années en tant qu’étudiante à la maîtrise, souvent interrompue par des élans de vagabondage, de maternité et de période professionnelle, j’ai un léger sourire en pensant à cette dernière décennie écoulée depuis mes débuts universitaires. Tout d’abord merci au Dr. Dominique Michaud pour sa grande qualité de vulgarisateur et son engouement pour ses projets de recherche, c’est en assistant à une de ses conférences que j’ai compris que je pouvais arrimer ma passion d’enfance et mon parcours académique dans une perspective professionnelle. Un remerciement particulier au Dr. Russell Tweddell pour son support et son expertise complémentaire en phytopathologie. Un petit clin d’oeil à Charles Goulet et à Jean-François Dubuc pour m’avoir abandonnée, seule, au laboratoire à mes débuts, ce fût finalement très formateur. Une ovation debout pour Marie-Claire Goulet, le phare des étudiants curieux en perpétuel égarement. Une accolade chaleureuse à tous mes collègues de laboratoire lève-tard qui ont prolongé ma phase HIGH de café matinal, un gros merci pour
les moments de complicité en fin de journée. UnPranayama à mon conjoint Patrick pour son
support inconditionnel et sa persévérance qui est devenue un modèle pour moi. Un câlin à mes filles Mathilde et Éliane, que j’aime de tout mon coeur, pour m’avoir permis de décrocher en temps opportun. Finalement, à tous les scientifiques qui ont la passion pour leur recherche, qui la vulgarisent, qui l’approfondissent, qui la mettent en pratique et surtout qui contribuent à faire de cette sphère d’activité un environnement dynamique, enrichissant et stimulant, je vous dis MERCI.
Avant-propos
Le présent mémoire intègre dans sa forme finale deux articles scientifiques rédigés en anglais pour publication dans des revues scientifiques arbitrées par des pairs.
Le Chapitre1du mémoire est le premier de ces articles, publié dans le journalBMC Plant
Bio-logy en novembre 2012. En tant qu’auteure principale, j’ai contribué au design expérimental
et à la réalisation d’une bonne partie des expériences décrites. Les expériences nouvellement réalisées s’ajoutaient à des résultats non publiés obtenus par les coauteurs Karine Coenen, Line Cantin et Louis-Philipe Vaillancourt, réanalysés ici à la lumière de mes hypothèses de recherche. Les coauteurs Charles Goulet et Marie-Claire Goulet ont participé pour leur part à la réalisation des expériences et à la rédaction de l’article. Frank Sainsbury et Russell Tweddell ont fourni des expertises complémentaires pour l’interprétation des données et l’écriture de l’article. Dominique Michaud, chercheur prinicipal du projet, a chapeauté et dirigé l’ensemble des travaux et préparé la version finale du manuscrit.
Le Chapitre 2 est aussi présenté sous la forme d’un article scientifique, soumis le 16 juillet
dernier au périodique BMC Plant Biology. En tant qu’auteure principale, j’ai contribué à la
réalisation d’une bonne partie des expérimentations, à la rédaction du manuscrit et à l’analyse des résultats, obtenus aussi en partie par les coauteurs Marie-Aube Simon et Moustafa Khalf. Dominique Michaud a dirigé l’ensemble des travaux de recherche reliés à cet article, puis rédigé la version finale du manuscrit. Afin de présenter un contenu cohérent, les articles sont présentés dans ce document sous leur forme originale, avec la section Matériel et Méthodes à la fin, suivie d’une bibliographie distincte pour chacun. Les références bibliographiques citées dans l’Introduction et dans la section Discussion et conclusion ont été regroupées à la fin du document, dans la section Bibliographie, afin d’en faciliter la consultation.
Introduction
Mise en contexte générale
La pomme de terre (Solanum tuberosum)est une culture importante pour l’alimentation
hu-maine à l’échelle mondiale, avec un taux de production record de 376 M de tonnes pour la
seule année 2013 (FAO,2013). Dans les dernières années, une augmentation de 1 M de tonne
produite par année a été observée mondialement, démontrant l’importance toujours grandis-sante de la demande pour cette culture. Le Canada, en 2013, a produit 4,6 M de tonnes de tubercules sur une superficie de 19 M d’hectares distribués sur tout le territoire, d’un océan
à l’autre (FAO,2013). Au Québec, la pomme de terre est la troisième production agricole en
importance et répond à 80 % de la consommation locale. Selon la Fédération des producteurs de pommes de terre du Québec, plus de 50 % de la production québécoise se retrouve dans nos assiettes, 20 % est destinée à l’industrie de la croustille et le reste utilisé pour les sous-secteurs
du prépelage et de la semence (FPPTQ,2013).
La pomme de terre est un produit versatile et un choix économique pour le consommateur, qu’elle soit consommée brute ou transformée. Considérée comme un aliment de base pour prévenir la malnutrition dans les pays en voie de développement, elle est un peu boudée par la population nord-américaine, souvent associée injustement à l’obésité et au diabète. En plus de contenir des ressources énergétiques importantes, la pomme de terre regorge de vitamines
essentielles, de minéraux et d’une quantité appréciable d’antioxydants (Camire et al.,2009).
Les problèmes de salubrité alimentaire pour les produits de la pomme de terre sont presque inexistants, bien que certains intrants ajoutés lors de la production et l’entreposage soulèvent des questionnements quant aux répercussions sur la santé. La pelure, en particulier, peut
contenir des résidus de nitrates ou des inhibiteurs de germination chimiques (Camire et al.,
2009), et des alternatives à ces produits sont recherchées afin de répondre aux inquiétudes
des consommateurs.
Selon le Répertoire des semences publié en décembre 2013, plus de 103 variétés de semences de pommes de terre ont été enregistrées pour le Québec, assurant une grande diversité génétique
des produits cultivés (Agri-Réseau, 2013). Ces variétés offrent des caractéristiques variées
et la forme du tubercule. Les semences montrent aussi des caractéristiques agronomiques diverses pour les agriculteurs, notamment pour la résistance aux maladies s’attaquant aux parties aériennes, aux racines ou aux tubercules. D’origines bactérienne, virale ou fongique, ou encore causés par des ravageurs arthropodes, les stress biotiques associés à cette culture revêtent une importance économique particulière pour le producteur, qui souhaite maximiser sa productivité en limitant l’émergence de maladies au champ et minimiser les pertes associées aux maladies pendant l’entreposage des tubercules. Le producteur désire offrir, aussi, un produit de qualité aux consommateurs, de belle apparence et avec une longue durée de vie tablette en épicerie.
La pomme de terre a été l’objet de nombreuses études en transgénèse depuis le milieu des années 1980, alors que les outils biotechnologiques utilisés à grande échelle aujourd’hui fai-saient leur apparition. Des variétés transgéniques de pommes de terre ont été développées
pour la résistance au doryphore (Leptinotarsa decemlineata), pour la résistance aux virus et
pour l’amélioration des propriétés nutritionnelles. La variété NewLeafTMdéveloppée par
Mon-santo, qui exprimait un gène de la bactérie Bt, a été commercialisée et cultivée au Québec entre 1999 et 2002, mais pour des raisons de marché liées aux controverses sur les cultures transgéniques n’est plus cultivée ni au Canada ni aux États-Unis. Malgré les controverses tou-jours vives, des travaux sont actuellement en cours afin de développer des variétés de pommes
de terre transgéniques résistantes au pathogènePhytophthora infestans, agent responsable de
la Grande famine en Irlande (GMOcompass,2008). Le séquençage du génome de la pomme de
terre, ses attributs génétiques et les outils biotechnologiques actuellement disponibles font de cette culture une candidate idéale pour l’amélioration génétique basée sur la biotechnologie (Barrell et al.,2013).
Dans ce contexte, le présent projet avait pour objet d’étudier des lignées de pommes de terre transgéniques dérivées de la variété Kennebec, une variété à chair blanche, exprimant des
in-hibiteurs de protéases (IPs) de la famille des cystatines (Benchabane et al.,2010). L’objectif
à la base du développement de ces lignées était de conférer à la plante hôte une résistance au doryphore de la pomme de terre via une inhibition de ses protéases digestives par l’effet inhibiteur de la cystatine exprimée de façon constitutive dans les feuilles et les autres tissus (Cloutier et al., 2000). Les cystatines forment un complexe avec les protéases digestives de type cystéine chez l’insecte ou l’agent pathogène ciblé, réprimant ainsi ses capacités
diges-tives et menant à des carences nutridiges-tives parfois mortelles (Broadway,2000). L’expression de
cystatines chez la plante, en revanche, soulève des questions sur sa biologie, dépendante aussi
de l’action de protéases de type cystéine potentiellement inhibées par les cystatines (Schlüter
et al.,2010). Si des études ont déjà démontré des effets potentiels des cystatines recombinantes sur le métabolisme des plantes transgéniques, très peu demeurait connu, à l’initiation de ce projet, sur les mécanismes expliquant ces effets.
Les cystatines en phytoprotection
Une première démonstration de l’implication des IPs dans la défense des végétaux a été faite par Clarence Ryan et son équipe en 1972, qui rapportaient alors une accumulation de ces protéines dans une plante soumise à l’herbivorie. Les auteurs démontraient que l’herbivorie par un doryphore chez la tomate occasionnait une accumulation systémique d’IPs dans les
organes aériens de la plante (Green and Ryan,1972). L’implication directe des IPs dans les
mécanismes de défense des plantes permettait par la suite de considérer ces protéines comme
un outil potentiel en transgénèse végétale à des fins de phytoprotection (Ryan,1990). Des
au-teurs rapportèrent, par exemple, les effets significatifs d’une cystatine de soja sur des extraits
de protéases d’insectes coléoptères (Hines et al.,1991), alors qu’une cystatine de maïs
expri-mée chez le riz montrait un effet inhibiteur sur les protéases digestives de plusieurs insectes (Irie et al.,1996). Des lignées transgéniques exprimant une cystatine étaient aussi créées,
prin-cipalement avec le transgène d’une cystatine du riz, l’oryzacystatine (OCI) (Masoud et al.,
1993; Sheng-Jun et al., 2004). Plusieurs espèces de plantes incluant des céréales, le colza, la pomme de terre et la patate sucrée étaient génétiquement transformées avec des transgènes de cystatines et des effets de protection notables étaient démontrés, notamment contre les
lépidoptères et les nématodes parasites (Gao et al.,2011;Lecardonnel et al.,1999;Vain et al.,
1998). Les cystatines végétales, ou phytocystatines, sont désormais exprimées couramment
dans des lignées végétales transgéniques pour la répression d’insectes herbivores ou d’agents pathogènes dépendants de protéases de type cystéine pour la digestion extracellulaire des
protéines alimentaires (Benchabane et al.,2010;Schlüter et al.,2010).
Les inhibiteurs de protéases
La protéolyse chez les végétaux
Malgré des développements prometteurs pour l’utilisation des IPs recombinants en phyto-protection, des questions demeurent quant aux impacts de ces protéines et à leur intérêt pratique sur le plan agronomique. En particulier, l’effet répresseur des IPs sur les protéases digestives des herbivores nuisibles a été étudié à de nombreuses reprises au cours des années (Zhu-Salzman and Zeng,2015), mais peu d’études se sont attardées à leurs effets éventuels
sur le métabolisme protéolytique de la plante modifiée (Schlüter et al.,2010). La protéolyse
est un mécanisme enzymatique crucial chez tout organisme vivant, qui intervient au niveau
physiologique et au cours du développement, y compris chez les plantes (Schaller,2004). Les
protéases, aussi appelées peptidases, sont les acteurs clés de ce processus. Une protéase est une enzyme qui a comme substrat une ou des protéines. Chez les plantes, les protéases assurent des rôles importants dans la sénescence, dans l’abscission foliaire, dans la mort cellulaire pro-grammée, dans la maturation des fruits et dans la mobilisation des protéines accumulées dans
le catabolisme des protéines de réserve dans une graine ou un tubercule est assuré par des protéases qui permettent de mobiliser les acides aminés nécessaires aux plantules en émer-gence lors de la germination. La protéolyse est essentielle aussi pour le maintien de l’intégrité cellulaire en permettant l’élimination des protéines anormales et inutiles par des voies de
dé-gradation sélectives ou non-sélectives, pour un recyclage ultérieur des acides aminés (Vierstra,
1996). Sur le plan biochimique, la protéolyse est un processus très complexe nécessitant un
contrôle serré de la séquence des événements et de la localisation du processus, autant dans la cellule que dans la plante entière.
Au niveau cellulaire, les protéases sont régulées par un contrôle rigoureux de leur expression au moment de la transcription et par une inhibition de leur activité après la traduction. La régulation post-traductionnelle des protéases est assurée par des IPs de nature généralement protéique, découverts en 1894 par Fermi et Pernossi dans des échantillons de plasma humain (Birk,2003; Fermi and Pernossi, 1894). Selon l’inhibiteur et la protéase en jeu, le processus
d’inhibition prend trois formes, schématisées à la figure0.1. Une première forme est l’inhibition
compétitive (A), où l’inhibiteur se lie de façon réversible au site actif de l’enzyme cible et entre en compétition directe avec le substrat, agissant ainsi comme un pseudo-substrat défini par sa charge, sa conformation et son affinité pour le site actif de la protéase. Contrairement à l’inhibition compétitive, l’inhibition non-compétitive (B) résulte de la fixation de l’inhibiteur à un site autre que celui du substrat sur l’enzyme, appelé site allostérique. Un complexe peut être formé avec l’enzyme libre ou avec le complexe enzyme-inhibiteur, rendant le complexe inactif dans les deux cas. L’inhibition incompétitive, ou anticompétitive (C), implique également une liaison de l’inhibiteur à un site différent du site de fixation du substrat, mais une liaison est possible seulement avec le complexe enzyme-substrat.
Sur une base plus spécifique, les protéases peuvent être classifiées en différents groupes selon trois critères majeurs : 1- la nature du site d’hydrolyse du substrat, 2- la nature chimique du site protéolytique de la protéase et 3- les homologies de séquences peptidiques entre protéases. La classification selon la nature du site d’hydrolyse est de loin la plus couramment utilisée pour la nomenclature des protéases dans la littérature. Dans ce cas, la spécificité d’hydrolyse des peptidases est liée à la nature des acides aminés qui se trouvent à proximité du site actif dans la conformation tri-dimensionnelle de la protéine. Selon la base de données sur les protéases MEROPS (merops.sanger.ac.uk), il existe sept clans permettant de différencier les types de protéases : les protéases de type aspartate, de type cystéine, de type glutamate, de type sérine, de type thréonine, les métalloprotéases et les protéases à mode d’action mixte. Dans la présente étude, notre attention s’est portée plus particulièrement sur la famille des protéases à cystéine de type papaïne, ou famille C1A qui, à ce jour, est représentée par plus de 3 000 séquences distinctes chez les plantes. Les protéases types de cette famille incluent la papaïne, les cathepsines B/D/H/L, les caspases et la bromélaine. Le mécanisme catalytique commun de ces protéases implique un acide aminé cystéine au sein de la triade catalytique
Enzyme Site actif S I
B
Enzyme Site actif S IA
C
S I Enzyme Site actifFigure 0.1 – Représentation schématique des types d’inhibition enzymatique (v. texte pour détails) : A) Inhibition compétitive, B) Inhibition non-compétitive, C) Inhibition anticompé-titive. S représente le substrat, I l’inhibiteur.
Les inhibiteurs de protéases compétitifs, comme par exemple les inhibiteurs des protéases végétaux, sont catégorisés en classes fonctionnelles calquées sur la classification des protéases. Ainsi, les inhibiteurs montrant une activité inhibitrice contre les protéases de type cystéine sont appelés inhibiteurs de type cystéine, parmi lesquels se retrouvent la leupeptine, la
chymo-statine, l’inhibiteur fongique E-64 et les cystatines de nature protéique (Barrett et al.,1982),
qui font l’objet du présent mémoire.
Les phytocystatines
Les cystatines sont des inhibiteurs de protéases de type cystéine isolées de sources aussi diverses que les végétaux, les animaux et une variété de microbes. Parmi les isoformes dé-couvertes, celles d’origine végétale, appelées phytocystatines, ont été étudiées le plus souvent dans une optique de phytoprotection. À ce jour, plus de 300 séquences ont été décrites chez les plantes et sont répertoriées dans la banque de données NCBI GenBank, et il est maintenant admis que leurs rôles ne touchent pas que l’inhibition des protéases exogènes, mais aussi celle
des protéases endogènes impliquées dans divers processus physiologiques (Benchabane et al.,
2010).
La première caractérisation d’une cystatine végétale a été réalisée par l’équipe de Soichi Arai,
au Japon, au milieu des années 1980 (Abe and Arai,1985). Isolée à partir de semences de riz,
cette protéine a été nommée Oryzacystatine (Abe et al.,1987), puis rebaptisée Oryzacystatine
I (OCI) après la découverte d’une cystatine homologue baptisée Oryzacystatine II (OCII) (Abe et al., 1991; Kondo et al., 1990). La structure 3D d’OCI a été élucidée partiellement
par cristallographie (Kudo et al.,1998) en 1998, puis décrite plus précisément sur la base de
données de résonance magnétique nucléaire en 2000 (Nagata et al.,2000). Les phytocystatines
montrent une structure sans pont disulfure, comportant une hélice alpha et un feuillet bêta
antiparallèle à cinq brins (Figure 0.2).
Des recherches effectuées chez différentes espèces végétales ont permis de recenser d’autres
phytocystatines, notamment chez la pomme de terre (Waldron et al.,1993), le soya (Misaka
et al.,1996), l’ananas (Shyu et al.,2004b), la papaye (Song et al.,1995) et le maïs (Abe et al.,
1995). Il est désormais bien établi que les cystatines végétales établissent une inhibition de
type compétitive avec leurs protéases cibles, tel qu’illustré par une modélisation in silico des
interactions établies entre les deux protéines (Benchabane et al.,2010;Sainsbury et al.,2012).
De nombreuses cystatines chez les plantes
Le groupe de Soichi Arai a adopté une approche similaire à celle utilisée pour OCI afin d’isoler de nouveaux homologues chez le maïs. Une protéine de 135 acides aminés bordée d’un signal peptide en position N-terminale a alors été isolée et nommée CCI, pour Corn Cystatin I (Abe et al., 1992). Des essais d’inhibition in vitro avec des protéases modèles ont montré
Figure 0.2 – Représentation structurale d’une cystatine végétale, la cystatine de
maïs CCII. Les structures secondaires sont ici mises en valeur, à savoir un feuillet bêta en
bleu, l’hélice alpha caractéristique en rouge et deux boucles en position basale qui pénètrent dans le site actif de l’enzyme cible (Image : Gracieuseté du Dr. Juan Vorster, U. Pretoria, Af. du Sud).
et al.,1994). La même équipe découvrait, un an plus tard, une autre cystatine chez le maïs,
la cystatine CCII, plus particulièrement active contre les cathepsines L (Abe et al., 1995)
similaires à celles retrouvées chez les insectes herbivores. Par la suite, huit autres cystatines
ont été identifiées chez le maïs (Massonneau et al., 2005), et il est maintenant établi que
le génome des végétaux est pourvu de séquences multiples pour les cystatines, présumément utiles pour l’inhibition d’une vaste gamme de protéases à cystéine impliquées à divers niveaux
chez la plante (Benchabane et al.,2010;Girard et al.,2007).
Le rôle des cystatines chez les plantes
À l’échelle du métabolisme, les cystatines ont pour fonction de réguler le turnover protéique, c’est-à-dire la durée de vie des protéines, par leur action de répression sur les protéases de type cystéine. Ces inhibiteurs peuvent contrôler des protéases dans la plante elle-même (inhibition
in planta, ou endogène) ou inhiber les protéases extracellulaires d’organismes en interaction
avec la plante (inhibitionex planta, ou exogène). Le nombre de variants de cystatines dans le
génome d’une même espèce et leur profil d’expression distinct d’un organe à l’autre ou sous diverses conditions environnementales suggèrent une diversité de fonctions pour ces protéines (Benchabane et al.,2010).
Les connaissances actuelles concernant l’identité précise des protéases ciblées par les cystatines
en sont à leurs balbutiements. La modélisation in silico et l’acquisition de connaissances sur
la structure des protéases permettent de suggérer des pistes, mais l’implication des cystatines dans la régulation des protéines endogènes chez les plantes a été principalement inférée indi-rectement par transgénèse. La majorité des cystatines identifiées jusqu’ici ont été découvertes
par l’étude de profils d’expression génique suite à un stress. La répression (ou le silencing)
des gènes de cystatines correspondants, ou à l’inverse leur surexpression dans la plante, ont
ensuite permis de formuler des conclusions quant à leurs rôles éventuels (Benchabane et al.,
2010).
Les fonctions des cystatines in planta
Chez les végétaux, les cystatines sont impliquées dans la régulation de plusieurs processus physiologiques en lien avec les rôles multiples attribués aux protéases de type cystéine, comme la régulation de la mort cellulaire programmée, la réponse aux stress abiotiques et le contrôle du turnover protéique dans les organes de réserve.
Turnover protéique dans les organes de réserve
La germination des graines et des tubercules est contrôlée par des protéases de type cys-téine qui assurent le catabolisme des procys-téines de réserve au moment opportun. Pour utiliser les acides aminés stockés et permettre la production des nouvelles protéines nécessaires au développement de la nouvelle plante, ce processus protéolytique est finement géré via des
signaux hormonaux et la présence d’inhibiteurs de protéases à action endogène (Weeda et al.,
2010). Les légumaines, une sous-famille des protéases à cystéine, activent d’abord des
pro-téases à cystéine de type papaine, qui elles entament la dégradation des protéines de réserve pour ainsi fournir une source d’azote et d’acides aminés libres aux organes de la plantule en développement. Il a été démontré que certaines cystatines montraient aussi une affinité
pour ces légumaines (Martinez et al., 2007; Martinez and Diaz, 2008), qui s’exprimeraient
pour empêcher le processus de germination pendant la période de dormance. Chez le blé, il
a été démontré qu’une cystatine régule des protéases à cystéine dans les semences (Kiyosaki
et al., 2007) et il est maintenant suggéré que l’initiation de la germination dans les organes de réserve est associée à une diminution de la teneur en cystatines, de manière à permettre aux protéases à cystéine d’entamer le catabolisme des protéines accumulées dans l’organe
de réserve (Shyu et al., 2004a; Valdes-Rodriguez et al., 2007). En revanche, lorsque le gène
d’une cystatine a été surexprimé chez Arabidopsis, la germination s’en est trouvée retardée, alors qu’une suppression de l’expression du même gène rendait à l’inverse la germination plus
précoce (Hong et al.,2007; Hwang et al.,2009). Ces études suggéraient, dans l’ensemble, un
Réponse aux stress abiotiques
Les cystatines joueraient aussi un rôle de protection contre les stress abiotiques (Benchabane
et al.,2010), associés notamment à des conditions environnementales défavorables comme un apport en eau limité, une luminosité déficiente, des températures extrêmes, des radiations solaires intenses ou des carences en certains minéraux. Des conditions environnementales s’éloignant de la normale peuvent amener la plante à réagir pour se protéger, notamment via des processus protéolytiques induits en conditions de stress cellulaire. L’implication des inhibiteurs de protéases dans la réponse aux stress abiotiques a été déduite à plusieurs reprises par des travaux rapportant une surexpression systématique de cystatines en condition de stress.
Par exemple, la cystatine de l’amaranthe AhCP1 a été identifiée dans les racines de la plante en réponse à des stress abiotiques comme un déficit en eau, une haute concentration en
sel, le froid ou la chaleur (Valdes-Rodriguez et al., 2007). Chacun de ces stress induisait
l’expression du gène de la cystatine, bien que la localisation cellulaire finale de la protéine différait selon le stress et l’organe impliqué. En comparaison, sept isoformes de cystatines ont été identifiées chez le maïs suite à des traitements thermiques. L’expression de ces isoformes
était modulée en fonction des tissus de la plante et du stress appliqué (Massonneau et al.,
2005). Chez Arabidopsis, deux isoformes de cystatines ont été identifiées suite à des stress de
sécheresse, de chaleur, de froid ou de blessures mécaniques (Hwang et al., 2010). Zhang et
al. identifiaient chez la même plante des cystatines surexprimées suite à des stress hydrique et salin, et démontraient que leur surexpression dans des lignées transgéniques procurait une
tolérance supplémentaire à ces mêmes stress (Zhang et al., 2008). Chez l’orge, la cystatine
HV-CP1 est surexprimée en conditions d’anaérobiose, lors d’un choc de froid ou suite à un
choc thermique (Gaddour et al.,2001). Des observations semblables ont été rapportées chez
la châtaigne d’eau (Pernas et al.,2000), le haricot (Diop et al.,2004), le blé (Christova et al.,
2006), la roquette de mer (Megdiche et al.,2009), la betterave sucrière (Wang et al., 2012),
la pomme (Tan et al.,2014), et plusieurs autres études montrant des effets de protection aux
stress abiotiques associés à l’expression d’une cystatine recombinante (Schlüter et al.,2010).
Régulation de la mort cellulaire programmée
La mort cellulaire programmée (MCP) est un autre processus où les cystatines joueraient un rôle de régulation. Ce processus implique la mort dirigée d’une cellule, suite à divers stimuli qui enclenchent une série d’événements cellulaires contrôlés et régulés à différents niveaux. Ce processus très utile au développement de la plante se manifeste aux toutes premières
étapes de son développement, puis jusqu’à sa mort (Pennell and Lamb, 1997). La MCP
joue un rôle dans la germination et le développement des graines, dans la reproduction pour la détermination du sexe de la plante, et dans les interactions entre le pollen et le pistil (Greenberg,1996). Chez la plante en croissance, la MCP est impliquée dans le développement des tissus végétatifs comme les feuilles et les racines. Elle est au coeur de la formation du
xylème et elle est impliquée activement dans les processus de sénescence pour le recyclage
des métabolites (Williams and Dickman,2008). La réponse aux stress biotiques et abiotiques
implique aussi la MCP, qui permet dans ces situations de limiter les dommages causés à l’organisme. Par exemple, la réaction hypersensible, un processus de MCP impliqué dans les interactions plantes-pathogènes, est une réaction de défense rapide activée lorsque certains agents pathogènes attaquent des tissus spécifiques. Des études ont fait état d’une interaction
entre les cystatines et les protéases initiant le processus de MCP (Belenghi et al., 2004;
Bonneau et al., 2008; Chichkova et al., 2004; Danon et al., 2004). Bien que le mécanisme d’interaction soit encore incompris, les cystatines ont clairement un rôle modulateur dans
l’établissement du phénomène et une surexpression de ces protéinesin planta aurait un impact
généralement répresseur sur le processus (Beers et al.,2000;Belenghi et al.,2003;Rojo et al.,
2004;Solomon et al.,1999).
Les fonctions des cystatines ex planta
Depuis la découverte du rôle des IPs dans la défense des plantes par l’équipe de Clarence Ryan, les voies de défense aux stress biotiques ont été étudiées par plusieurs groupes de recherche au fil des années. Les stress biotiques auxquels sont soumis les plantes impliquent une gamme d’organismes dits nuisibles, en particulier les insectes herbivores et les agents pathogènes. Les cystatines font partie du large arsenal de composés utilisés par les plantes pour se défendre contre ces organismes et il est maintenant connu qu’elles jouent un rôle actif dans la réponse à plusieurs stress biotiques.
Résistance aux insectes
L’effet des cystatines dans la défense des plantes est lié à leur impact sur l’activité des pro-téases digestives chez les organismes visés. Le tube digestif des insectes herbivores, notamment chez les coléoptères, montre une activité protéolytique digestive principalement de type
cys-téine (Murdock et al., 1987; Soares-Costa et al., 2011). La digestion des tissus foliaires dans
l’intestin de ces herbivores nécessite un cocktail enzymatique pour briser mécaniquement et chimiquement les structures de la feuille. Pour les plantes d’intérêt agronomique, une dimi-nution de la surface foliaire par l’herbivorie cause des pertes de rendement considérables, et l’inhibition des enzymes digestives des insectes ravageurs par l’expression de cystatines recom-binantes s’est avérée une approche potentiellement utile en phytoprotection depuis les années
1990 (Haq et al.,2004; Schlüter et al., 2010). À l’échelle moléculaire, les cystatines agissent
comme un pseudo-substrat en formant un complexe réversible avec les protéases digestives de type cystéine dans le tube digestif de l’insecte cible, réprimant l’activité protéolytique et
causant des carences nutritives chez le ravageur (Broadway,2000)(Figure0.3). Ce « scénario
idéal » pour la plante peut toutefois être contrecarré par des phénomènes adaptatifs chez
l’in-secte, qui sécrète rapidement des protéases insensibles à la cystatine recombinante (Bouchard
de-Protéases de la plante Protéases de l'insecte IP IP IP Inhibition Problèmes de croissance Compensation PLANTE INSECTE
Figure 0.3 – Représentation schématique de l’effet inhibiteur d’une cystatine dans un système plante-insecte.
meure prometteur pour la lutte aux insectes, grâce au développement d’inhibiteurs toujours
plus performants (Schlüter et al.,2010).
Résistance aux champignons
Les cystatines végétales montrent aussi des effets adverses contre les champignons phytopatho-gènes, en causant un arrêt de la croissance du mycélium ou l’inhibition de la germination des spores. Bien que le mécanisme d’action des cystatines ne soit pas encore bien déterminé, des études ont clairement démontré des effets antifongiques pour les cystatines recombinantes. Par exemple, une étude réalisée sur le taro, un légume racine qui démontre une grande résistance aux agents pathogènes, a révélé un effet inhibiteur de la cystatine CceCPI sur six espèces de
champignons des genresFusarium et Sclerotinia (Yang and Yeh,2005). De même, la cystatine
Csc de la châtaigne a réprimé la croissance mycélienne du champignon nécrotrophe Botrytis
cinerea par l’inhibition de 50% de ses protéases extracellulaires (Pernas et al.,1998). Il a été suggéré que l’activité antifongique des cystatines pouvait être reliée à l’inhibition de protéases
de type cystéine nécessaires à la formation des parois cellulaires du champignon (Soares-Costa,
2002). Dernièrement, une cystatine du kiwi a été identifiée et son activité antifongique reliée
à l’inhibition de la germination des spores des agents pathogènes Alternaria radicina et B.
cinerea (Popovic et al.,2012). D’autres études ont démontré une diminution de la croissance mycélienne chez des plantes surexprimant une cystatine sans inhibition apparente des pro-téases du champignon, des observations suggérant des effets indirects des cystatines via une
modification possible de la plante transformée pour les exprimer (Dutt et al.,2012;Martinez
Résistance aux virus
Des effets indirects pourraient aussi expliquer l’effet antiviral des cystatines chez les plantes. Plusieurs hypothèses ont été émises pour expliquer l’impact négatif de ces protéines sur des virus pathogènes. Une étude a suggéré, par exemple, que des variétés transgéniques de ta-bac résistantes à certains virus, comme le virus de la mosaïque du tata-bac, étaient protégées directement par la cystatine, qui inhiberait une protéase de maturation du virus et en
compro-mettrait du même fait la réplication (Gutierrez-Campos et al.,1999). Selon d’autres auteurs,
la cystatine supprimerait la mort cellulaire programmée dans la plante sans même inhiber les protéases du virus, et aurait ainsi un effet antiviral indirect par un effet d’interférence sur le
métabolisme cellulaire de la plante-hôte (Gholizadeh et al.,2005).
Quoi qu’il en soit, les cystatines ont des fonctions importantes pour le métabolisme de la plante, jouant notamment un rôle non négligeable dans les réponses aux stress. Les interac-tions entre les végétaux et les agents pathogènes sont toutefois très complexes et évoluent rapidement, rendant souvent la compréhension de leurs effets plutôt ardue. Il est à prévoir néanmoins que nos connaissances sur la régulation des protéases végétales, y compris celles impliquant les cystatines, s’approfondiront dans le futur et nous permettront de développer des stratégies toujours plus efficaces en phytoprotection.
Des effets pléiotropiques pour les cystatines recombinantes ?
Si les perspectives d’application apparaissent prometteuses pour les cystatines recombinantes en protection des plantes, plusieurs études récentes ont soulevé en revanche des questions
quant à leurs éventuels effets d’interférence métabolique (ou effets pléiotropiques) in planta.
Déjà en 2001, une équipe mexicaine rapportait des effets pléiotropiques liés à l’expression d’une
cystatine chez le tabac (Gutierrez-Campos et al., 2001). Ces lignées exprimant la cystatine
du riz OCI manifestaient des caractéristiques avantageuses au niveau morphologique, tels un taux de croissance accéléré, une biomasse accrue, une floraison précoce et un nombre supérieur de fleurs et de graines. De manière similaire, Van der Vyver et al. observaient, au niveau physiologique, un maintien du taux de photosynthèse et un meilleur taux de récupération général après un stress de froid chez des lignées transgéniques exprimant OCI, expliqués par
l’intégrité maintenue de l’appareil photosynthétique (Van der Vyver et al.,2003) et un contenu
protéique accru dans les feuilles (Prins et al., 2008). Au niveau métabolique, l’expression
de la cystatine d’Arabidopsis AtCYS1 dans des cellules de tabac transgénique causait une
inhibition du processus de MCP initié par un agent pathogène ou un stress oxydatif (Belenghi
et al., 2003). Récemment, des plants de tabac exprimant OCI démontraient une protection des enzymes impliquées dans la réponse aux stress oxydatifs, telles la superoxyde dismutase
et des peroxydases, dans des plantes soumises à des stress abiotiques (Demirevska et al.,
2010). Ces effets pléiotropiques associés aux cystatines, s’ils apparaissent d’intérêt potentiel en
pratique, reflètent en revanche des lacunes majeures dans nos connaissances sur les mécanismes protéolytiques mis en place dans la plante hôte.
Le cas de la pomme de terre, Solanum tuberosum
Dans cette perspective, l’objet du présent mémoire était de cerner plus à fond les effets pléio-tropiques des cystatines recombinantes en utilisant comme modèle d’étude la pomme de terre,
Solanum tuberosum. Les lignées transgéniques étudiées étaient dérivées de transformations
gé-nétiques avec les gènes des cystatines OCI (du riz) et CCII (du maïs) exprimés sous le contrôle du promoteur CaMV 35S, une séquence qui dirige une expression constitutive du transgène dans l’ensemble des tissus de la plante, incluant le tubercule.
Le tubercule, un organe de réserve au contenu variable
Le tubercule, un organe de réserve destiné à la consommation humaine, peut être entreposé jusqu’à 12 mois avant d’être consommé. Il est toutefois sensible aux maladies pendant son dé-veloppement et son entreposage post-récolte. Malgré l’importance historique de cette culture, les mécanismes de tubérisation, de dormance et de germination du tubercule ne sont pas encore tout à fait élucidés. La formation du tubercule se divise en plusieurs étapes, au cours desquelles des macromolécules majeures s’accumulent pour constituer les réserves essentielles à la germination future et pour assurer une protection contre les maladies et les stress pendant la période de dormance. Le tubercule parvenu à maturité contient de l’amidon, son sucre le
plus important, des protéines, des lipides et une gamme variée de composés mineurs (Suttle,
2007).
Au moment de la récolte, le tubercule de pomme de terre est chargé de macromolécules, mais son vieillissement en cours d’entreposage entraîne leur dégradation au profit des germes en croissance. La dormance du tubercule est un processus complexe qui implique plusieurs
états physiologiques contrôlés par des signaux hormonaux et environnementaux (Sonnewald
and Sonnewald, 2014). La levée de la dormance mène à l’initiation de la germination, où les tubercules deviennent des organes sources pour supporter la croissance des germes et compléter le cycle de reproduction végétative de la plante. Au cours de ce processus, l’amidon et les protéines sont dégradés afin de libérer des sucres solubles et des acides aminés. Ces réactions cataboliques sont généralement accompagnées, au cours de l’entreposage, d’une perte
de poids du tubercule et d’un ramollissement de sa chair (Sonnewald, 2001). En pratique,
un retard dans la germination du tubercule représente un avantage économique pour les industriels qui désirent offrir un tubercule riche en nutriments, avec une apparence saine et une surface exempte de germes.
Le contrôle de la dormance en contexte industriel
Les pratiques industrielles actuelles destinées à assurer une bonne qualité des tubercules impliquent un entreposage à des températures froides de l’ordre de 2 à 4°C et l’application d’inhibiteurs de germination chimiques. L’entreposage des tubercules au froid retarde leur
germination, mais engendre une hausse du taux de sucres solubles due à la dégradation de
l’amidon de réserve (Coffin et al., 1987), une caractéristique indésirable pour des tubercules
destinés à la transformation alimentaire. Généralement, l’industrie combine une température d’entreposage optimale et une application d’inhibiteurs de germination chimiques pour le
contrôle de la dormance (Aksenova et al., 2013). De nombreux facteurs influencent l’état de
dormance des tubercules et les solutions de traitements chimiques aident les producteurs à minimiser les pertes économiques. Davantage de connaissances sont toutefois essentielles afin de développer de nouvelles stratégies moins dommageables pour la santé humaine et la santé
environnementale (Kleinkopf et al.,2003).
Régulation de la dormance et balance protéases:inhibiteurs
Il est maintenant bien établi que les hormones contrôlent la levée de dormance du tubercule. L’acide abscissique, en particulier, est associé au maintien de la dormance alors que les
gibbé-rellines et les cytokinines favorisent sa levée (Campbell et al.,2010;Carrera et al.,2000;Suttle
and Hultstrand, 1994). Au fur et à mesure que la dormance se prolonge, la teneur en acide abscissique diminue en parallèle au déclin de l’expression des IPs, toutes classes confondues (Callis,1995;Campbell et al.,2008;Schaller,2004).
Un inhibiteur de protéases clé dans le tubercule est la multicystatine de pomme de terre, ou
potato multicystatin (PMC), un inhibiteur de protéases à cystéine composé de huit domaines
inhibiteurs distincts (Waldron et al.,1993;Walsh and Strickland,1993). Cette protéine joue un
rôle important dans la déposition des protéines de réserve pendant la tubérisation en régulant à la baisse l’activité protéolytique du tubercule expliquée majoritairement par des protéases
de type cystéine (Michaud et al., 1994; Weeda et al., 2009). La multicystatine contribue,
en particulier, à l’accumulation de la patatine, tel que suggéré par une corrélation positive
dans l’accumulation de ces deux protéines majeures dans le tubercule en croissance (Weeda
et al.,2009). À l’opposé, la levée de la dormance et l’initiation de la germination dépendent de signaux hormonaux qui enclenchent l’activité protéolytique pour le catabolisme des protéines, au détriment de la multicystatine qui se retrouve dégradée en acides aminés transférés aux
nouveaux germes (Michaud et al.,1994;Weeda et al.,2010).
Sur un plan biochimique, c’est la balance des teneurs en inhibiteurs de protéases et en pro-téases qui déterminerait la cadence du catabolisme des réserves protéiques, et du même fait
l’établissement du processus germinatif dans les organes de réserve (Benchabane et al.,2010).
Dans le tubercule de pomme de terre, il a été établi que la teneur en multicystatine, un régu-lateur des protéases endogènes, diminuait de façon linéaire lors de la mobilisation des réserves
protéiques, le tout associé à une activité protéolytique totale accrue (Weeda et al.,2010). Le
catabolisme des protéines de réserve, qui sont à la fois des protéines de défense importantes et des régulateurs du développement, rend alors le tubercule plus vulnérable aux maladies d’entreposage et favorise l’émergence de germes, source de problèmes économiques pour les
agriculteurs. Une attention particulière a été portée à cette question dans le cadre du présent mémoire, considérant l’effet répresseur des cystatines recombinantes rapporté antérieurement
sur la germination des semences d’Arabidopsis (Hwang et al.,2009) et par conséquent l’effet
répresseur possible des cystatines OCI et CCII sur le processus de débourrement des germes.
Hypothèses et objectifs du projet
En résumé, les sections précédentes ont fait état de l’importance des cystatines au sein de plu-sieurs processus physiologiques chez les plantes. Depuis la mise en évidence de leur implication dans la défense, des gènes de cystatines ont été clonés chez diverses espèces végétales, avec le plus souvent pour objectif de développer des lignées transgéniques résistantes à des orga-nismes nuisibles tels les insectes, les champignons, les bactéries et les virus. Les recherches en transgénèse, combinées à l’apparition de nouveaux outils analytiques, ont permis en parallèle d’étudier les plantes transgéniques au-delà de leurs interactions avec les organismes nuisibles ciblés. Dans le cas des inhibiteurs de protéases, les recherches se sont également consacrées à l’étude de la plante transgénique elle-même qui, dans une perspective de culture et de commercialisation, doit être métaboliquement stable pour sa production à grande échelle.
Étant donné l’implication des protéases de type cystéine dans divers processus physiologiques chez les plantes, il nous apparaissait alors réaliste de proposer que l’expression d’une cystatine recombinante engendrait des perturbations métaboliques dans la plante modifiée.
Des recherches antérieures avaient démontré que l’expression constitutive d’une cystatine végétale chez le tabac causait des effets d’interférence métabolique se traduisant par des
répercussions sur la morphologie de la plante (Gutierrez-Campos et al.,2001). Une tolérance
accrue à des stress abiotiques comme le froid, la salinité ou la sécheresse était aussi observée, vraisemblablement explicable par un effet d’inhibition sur des protéases à cystéine et/ou sur
l’induction de différents gènesin planta. Il était déjà connu que des protéases de type cystéine
sont impliquées au sein de mécanismes de défense endogènes dans les parties végétatives de la plante en réponse aux stress biotiques, de même que dans les organes de réserve pour un contrôle adéquat du processus germinatif, d’où la pertinence d’étudier en détail l’impact possible des cystatines recombinantes exprimées sur la physiologie et le métabolisme de la plante hôte.
Depuis 20 ans, notre équipe de recherche se consacre à la transgénèse de la pomme de terre
avec des gènes de cystatines végétales (Benchekroun et al.,1995). À l’origine, des lignées de
pommes de terre de variété blanche, v. Kennebec, ont été transformées avec la cystatine du riz OCI dans une perspective de phytoprotection. Ces lignées ont été caractérisées et testées pour leur effet protecteur contre le doryphore de la pomme de terre, principal ravageur de cette
culture au Québec (Bouchard et al.,2003a,b;Cloutier et al.,1999,2000). Des études ont aussi
(Simon, 2001). D’autres lignées transgéniques ont été générées avec CCII puis caractérisées pour leur phénotype et leur métabolisme afin de documenter davantage l’établissement d’effets
pléiotropiques éventuels in planta (Pépin,2005;Vaillancourt,2005).
En bref, les lignées de pommes de terre exprimant OCI ou CCII ont montré dans certains cas un phénotype altéré, un retard de floraison et/ou une morphologie anormale du tubercule (Vaillancourt,2005), des résultats qui s’apparentent à ceux rapportés dans la littérature avec d’autres espèces. Certaines de nos lignées transgéniques ont aussi révélé une différence com-parativement aux plantes témoins, à savoir l’expression constitutive de la multicystatine de pomme de terre (PMC) dans les feuilles. Cette protéine est normalement induite suite à un
stress biotique comme l’herbivorie ou par un traitement à l’acide jasmonique (Bolter, 1993;
Girard et al., 2007). Considérant les rôles endogènes de la multicystatine et son accumula-tion importante dans le cytosol des cellules foliaires, il nous apparaissait pertinent d’étudier l’effet de l’expression des cystatines de céréales OCI et CCII sur le protéome des feuilles et d’en évaluer l’impact sur les mécanismes de défense normalement inductibles en conditions de stress.
Le tubercule, dans lequel les réserves protéiques se dégradent au cours de l’entreposage, est également régulé par la multicystatine, qui joue un rôle de protection en limitant le cata-bolisme protéique de type cystéine. Concentrée sous la pelure, cette protéine sensible à la
digestion tryptique (Nissen et al.,2009) est reconnue comme étant un régulateur du contenu
en patatine (Weeda et al., 2009), principale protéine de réserve du tubercule. Puisque les
lignées transgéniques générées au laboratoire exprimaient une cystatine de céréale de manière constitutive et ce, dans tous les tissus de la plante, il nous apparaissait intéressant de véri-fier si cet « ajout » au protéome du tubercule pouvait renforcer l’effet anti-germinatif de la multicystatine en maintenant le ratio cystatines/protéases plus élevé et en ralentissant le
ca-tabolisme protéique. Une étude antérieure réalisée chez la plante modèleArabidopsis thaliana
laissait suggérer un tel effet des cystatines recombinantes dans les semences. La surexpression de la cystatine AtCYS6 avait causé, dans ce cas, un délai dans la germination des graines,
observation corrélée avec une diminution de l’activité protéolytique de type cystéine (Hwang
et al.,2009).
Sur l’ensemble de ces observations, nous avons émis l’hypothèse principale suivante quant aux effets des cystatines CCII et OCI exprimées dans nos lignées transgéniques de pommes de
terre :L’expression constitutive d’une cystatine de céréale chez la pomme de terre
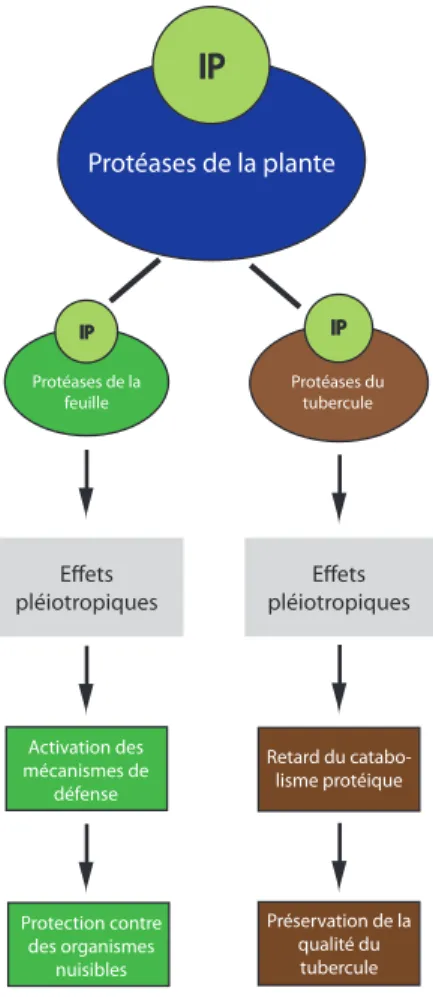
engendre une activation des protéines de stress dans les feuilles et un retard de germination du tubercule (Figure 0.4). Subordonnée à cette hypothèse, nous avons émis
l’hypothèse secondaire suivante, à l’effet que : L’activation du protéome de stress par
une cystatine recombinante a un effet protecteur antifongique dans les parties aériennes de la plante.
Protéases de la plante IP Effets pléiotropiques Activation des mécanismes de défense Retard du catabo-lisme protéique Protéases du tubercule IP Protéases de la feuille IP Effets pléiotropiques Protection contre des organismes nuisibles Préservation de la qualité du tubercule
Figure 0.4 – Schématisation de l’hypothèse de travail
Les objectifs de recherche suivants ont été poursuivis pour tester ces deux hypothèses :
Pour les feuilles :
1. Caractériser, chez des lignées de pommes de terre transgéniques exprimant CCII, un ensemble d’indicateurs de croissance et métaboliques de base, en complément à des
tests antérieurs réalisés sur ces lignées dans notre laboratoire (Vaillancourt,2005).Cette
étape devait permettre de sélectionner des lignées transgéniques semblables, de phénotype normal, pour la poursuite des travaux.
2. Évaluer l’impact de CCII sur le protéome de défense des feuilles dans les lignées
trans-géniques. Cette approche, qui permet l’étude des protéines du tissu étudié dans leur
ensemble, devait permettre d’identifier les variations statistiquement significatives dans les lignées transgéniques comparativement à un témoin non-transgénique.
3. Évaluer le degré de résistance des lignées CCII au champignon phytopathogène
nécro-trophe B. cinerea et documenter le mécanisme effecteur de la cystatine. Cet objectif
permettrait de vérifier si la cystatine recombinante pouvait conférer à la plante une meilleure résistance au champignon. Ces expériences avaient aussi pour but de détermi-ner si la cystatine agissait par une interaction directe avec le pathogène ou, à l’inverse, par des effets indirects liés aux caractéristiques de la plante modifiée.
Pour le tubercule :
4. Évaluer l’impact de l’expression des cystatines OCI et CCII sur le degré de préservation des macromolécules majeures du tubercule en période d’entreposage.
5. Évaluer l’impact d’OCI sur la germination suite à la levée de la dormance.
6. Évaluer le profil du développement des plantes OCI en émergence suite à l’entreposage du tubercule.
Les objectifs de recherche 1 à 3 ont été réalisés sur les feuilles de lignées CCII (Chapitre
1) ; les résultats sont présentés sous la forme d’un article scientifique publié dans le jounal
BMC Plant Biology en 2012 (Munger et al.,2012). Les objectifs 4 à 6 ont été réalisés sur des
tubercules exprimant OCI et/ou CCII (Chapitre 2) ; les résultats sont également présentés
Chapitre 1
Effets bénéfiques inattendus d’une
cystatine de céréale dans des lignées
transgéniques de pomme de terre,
Solanum tuberosum
1.1 Résumé
La découverte du potentiel des cystatines en phytoprotection a permis de développer des va-riétés végétales transgéniques résistantes aux insectes nuisibles. La création de ces « lignées à cystatines » a depuis été rapidement proposée par plusieurs, alors que d’autres auteurs rapportaient des effets d’interférence métabolique dans la plante suite à l’expression de cys-tatines. Nous avons tenté ici de comprendre ces effets pléiotropiques à l’aide de lignées de pommes de terre exprimant la cystatine du maïs (CCII) de façon constitutive dans l’ensemble des tissus de la plante. Nous avons d’abord évalué l’impact de cette transformation génétique sur le protéome foliaire de la plante modifiée afin d’obtenir un portrait global des effets de la protéine recombinante sur son métabolisme. Le protéome foliaire de deux lignées transgé-niques exprimant CCII a été résolu par électrophorèse bi-dimensionnelle (2-DE) et comparé à celui de la lignée mère K utilisée comme parent pour la transformation. Environ 700 pro-téines ont été détectées sur les gels, parmi lesquelles 23 variaient significativement dans les lignées CCII, incluant 14 protéines néo-synthétisées. L’identification de ces protéines régulées par spectrométrie de masse a révélé des protéines reliées aux stress biotiques et abiotiques, notamment des peroxydases, des inhibiteurs de protéases et des protéines reliées à la pa-thogenèse. Suggérant une activation constitutive des mécanismes de défense, des bio-essais
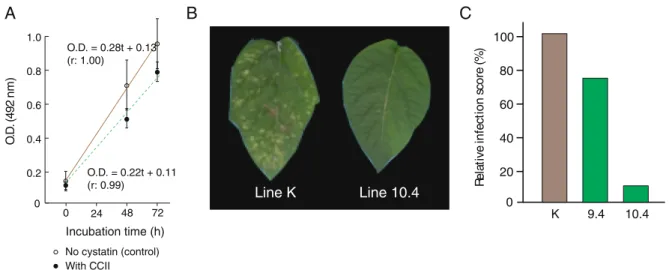
ont été menés avec le champignon phytopathogène Botrytis cinerea, agent responsable de la
la cystatine recombinante dans les lignées transgéniques. Après 12 jours d’infection, l’agent pathogène n’avait infecté que 10% des surfaces foliaires pour une lignée CCII relativement au témoin K, considéré infecté à 100%. L’effet protecteur de CCII était expliqué par un effet indirect de la cystatine lié à l’activation des protéines de défense dans les lignées CCII, et non par un effet inhibiteur sur les protéases extracellulaires du champignon. Les protéases
secré-tées de B. cinerea, qui sont utilisées pour faciliter l’infection, n’ont démontré aucune affinité
pour CCII, tel que démontré par des essais protéolytiques in vitro. Ces résultats suggèrent
dans l’ensemble un effet protecteur de la cystatine CCII via l’induction de mécanismes de défense par un processus encore méconnu. Il est clair, à la lumière de ces résultats, que les cystatines recombinantes représentent des modulateurs des réponses de stress potentiellement utiles pour créer des variétés végétales transgéniques tolérantes à différents stress biotiques et abiotiques.
Beneficial ’unintended effects’ of a cereal cystatin in transgenic
lines of potato, Solanum tuberosum
Munger A, Coenen K, Cantin L, Goulet C, Vaillancourt LP, Goulet MC, Tweddell R, Sainsbury F and Michaud D.
Centre de recherche en horticulture, Département de phytologie, Université Laval, Pavillon des Services, 2440 boul, Hochelaga, Québec QC, G1V 0A6, Canada.
1.2 Abstract
Background: Studies reported unintended pleiotropic effects for a number of pesticidal
pro-teins ectopically expressed in transgenic crops, but the nature and significance of such effects
in planta remain poorly understood. Here we assessed the effects of corn cystatin II (CCII), a
potent inhibitor of C1A cysteine (Cys) proteases considered for insect and pathogen control, on the leaf proteome and pathogen resistance status of potato lines constitutively expressing this protein.
Results: The leaf proteome of lines accumulating CCII at different levels was resolved by
2-dimensional gel electrophoresis and compared with the leaf proteome of a control (parental) line. Out of ca. 700 proteins monitored on 2-D gels, 23 were significantly up- or downregulated in CCII-expressing leaves, including 14 proteins detected de novo or up-regulated by more than five-fold compared to the control. Most up-regulated proteins were abiotic or biotic stress-responsive proteins, including different secretory peroxidases, wound inducible protease inhibitors and pathogenesis-related proteins. Accordingly, infection of leaf tissues by the fungal
necrotroph Botryris cinerea was prevented in CCII-expressing plants, despite a null impact
of CCII on growth of this pathogen and the absence of extracellular Cys protease targets for the inhibitor.
Conclusions: These data point to the onset of pleiotropic effects altering the leaf proteome
in transgenic plants expressing recombinant protease inhibitors. They also show the potential
of these proteins as ectopic modulators of stress responses in planta, useful to engineer biotic
or abiotic stress tolerance in crop plants of economic significance.
1.3 Background
Several studies have described the potential of recombinant protease inhibitors as potent an-tidigestive compounds to protect crop plants from herbivory or pathogenic infection [1,2]. For instance, cysteine (Cys) protease inhibitors of the cystatin protein superfamily were propo-sed as protective agents against various herbivorous arthropods and root-parasitic nematodes [3,4]. Cystatins and most other protease inhibitors in plants are competitive protein inhibitors
acting as pseudosubstrates to enter the active site of proteases [5]. Following inhibition, the target proteases can no longer cleave peptide bonds, which results in a detrimental inhibition of protein digestive functions in herbivorous pests and accounts for the resistance of several transgenic plant lines expressing recombinant inhibitors.
The heterologous expression of protease inhibitors in plants, however, has raised a number of questions about the possible occurrence of unintended metabolic interference –or pleiotropic effects– on endogenous proteolysis, which could eventually alter important cellular functions [2]. Proteases are ubiquitous metabolic effectors involved in the regulation of numerous cellu-lar processes, ranging from housekeeping functions like protein turnover and the elimination of misfolded polypeptides to the processing of polypeptide pre- and pro-regions on maturing protein backbones [6,7]. While studies have reported negligible phenotypic effects for protease inhibitors in transgenic plants based on the assessment of macroscopic indicators such as growth rate, stem diameter or leaf number [8-10], several reports suggest the onset of more subtle effects at the metabolic level. For instance, plant and mammalian serine (Ser) protease inhibitors ectopically expressed in potato were shown to significantly impact protein levels in leaves, positively or negatively [10,11]. Similarly, recombinant cystatins expressed in
Arabi-dopsis or tobacco were shown to induce a range of phenotype alterationsin planta, including
a delayed development of floral organs [12], a modified physiological behaviour under low temperature or light regimes [13], an altered protein content in leaves 13,14], and a strong repression of the pathogen inducible hypersensitive response [15].
These findings, along with studies reporting the tolerance of protease inhibitor-expressing plants to abiotic stress cues such as drought, salinity and low temperatures [13,16-18], point to the occurrence of endogenous protease targets for the recombinant inhibitors, directly or indirectly involved in stress-related processes. The so-called pleiotropic effects of recombinant protease inhibitors, which are often considered as unintended metabolic effects in the modified
plant, might simply reflect a lack of knowledge on stress-related proteolysis in planta and in
fact represent a source of potentially useful traits for crop improvement [2]. Here we provide experimental evidence for the up-regulation of abiotic and biotic stress-related proteins in leaves of transgenic potato lines engineered to express corn cystatin II (CCII), a potent inhibitor of C1A Cys proteases exhibiting potential for herbivore pest control [19,20]. We also link the pleiotropic effects of CCII expression with the compromised ability of a model
1.4 Results and discussion
Pathogenesis-related proteins are up-regulated in CCII-expressing potato lines
Reverse transcriptase (RT) PCR, immunodetection and surface-enhanced laser desorption io-nization time-of-flight mass spectrometry (SELDI TOF MS) analyses were performed to select
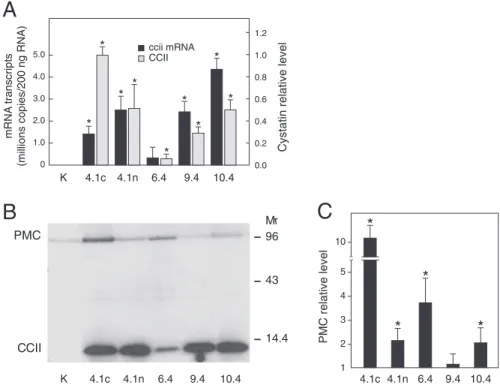
transgenic potato lines expressing CCII at different levels in leaves (Figure 1.1A,B), among
a collection of independent transformants developed earlier in our laboratory [21]. CCII in these lines accumulates in the cytosolic compartment, under the control of the Cauliflower mosaic virus 35S constitutive promoter. Unlike control line K showing no signal, the transge-nic lines showed variable, but easily detectable signals of CCII-encoding mRNA transcripts
(Figure1.1A). Accordingly, CCII was immunodetected as a 12-kDa polypeptide band of
va-riable intensity in the modified lines, in contrast with line K giving no signal (Figure 1.1B).
The amount of CCII in leaf extracts was variable among the lines, as inferred by SELDI TOF MS after capture of the inhibitor on CM-10 biochips for weak cationic exchange
(Fi-gure 1.1A). Unexpectedly, a clear immunoblot signal was detected in several CCII-lines for
the wound inducible homologue of CCII in potato leaves, potato multicystatin (PMC) [22]
(Figure1.1B). This apparent upregulation of the endogenous inhibitor was confirmed by
den-sitometric analysis of the immunoblots revealing significant increases reaching two- to tenfold the signals observed for control line K in four transgenic lines, out of five tested (Student’s
t-test ; P<0.05) (Figure 1.1C).
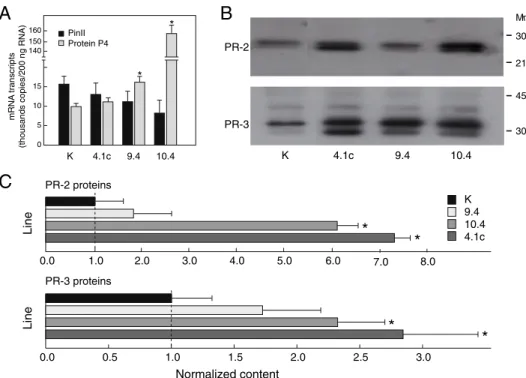
RT PCR assays were conducted with nucleotide primers for mRNA transcripts of protei-nase inhibitor II (Pin-II), a protein marker of wound and jasmonate inducible proteins ; and pathogenesis-related (PR) protein P4, a protein marker of the abiotic stress-/pathogen
indu-cible salicylic acid pathway [23] (Figure 1.2A). Transcript levels for Pin-II were not different
in control and transgenic lines (Student’s t-test ; P=0.10), while transcripts for protein P4
were found at higher levels in several CCII-expressing lines, including line 9.4 and line 10.4 (P<0.05). This observation suggesting an up-regulation of salicylate inducible proteins in the CCII-expressing lines was further supported by immunodetection assays with primary anti-bodies for PR-2 (β-glucanase) and PR-3 (chitinase) proteins, which showed levels of proteins from both families to be significantly higher in lines 4.1c, 9.4 and 10.4 compared to control
line K (Student’s t-tests ; P<0.05) (Figure 1.2).
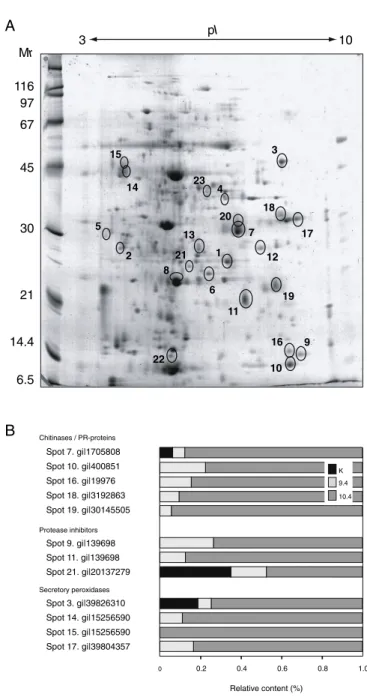
The stress-related proteome is altered in CCII-expressing leaves
A comparative two-dimensional gel electrophoresis (2-DE) proteome analysis was conducted with leaf protein extracts of line 9.4, line 10.4 and control line K to measure the overall impact of CCII on the host plant’s leaf proteome, and to test the hypothesis of a specific up-regulation of salicylate inducible stress-related proteins in plants ectopically expressing