Chemical Conditions in the Boundary Layer Surrounding Phytoplankton Cells Modify Cadmium Bioavailability.
30
0
0
Texte intégral
Figure
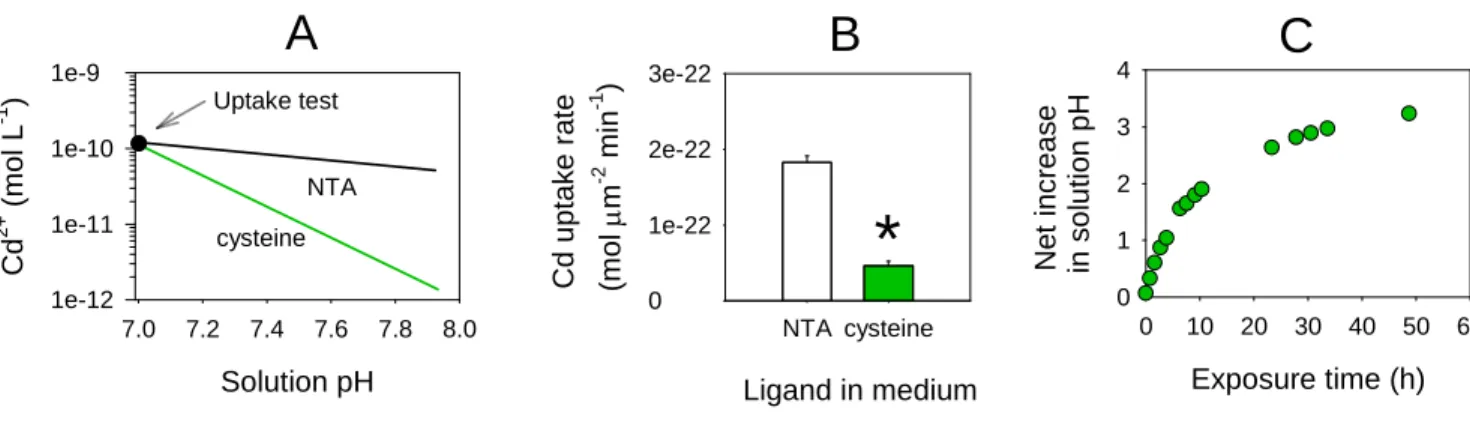
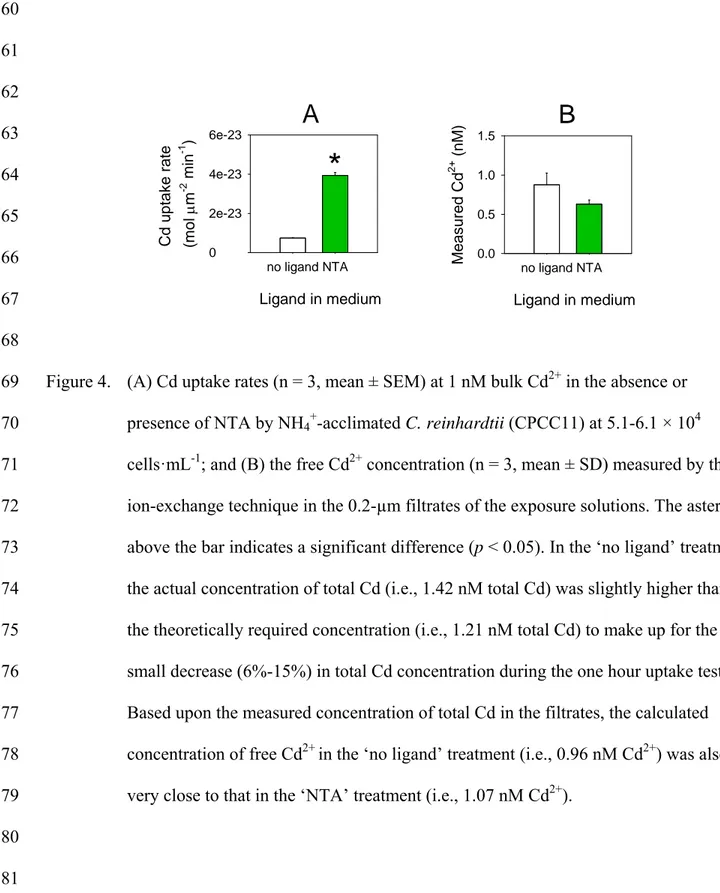
Documents relatifs
