La myopathie facio-scapulo-humérale : un défaut de répression?
Texte intégral
Figure
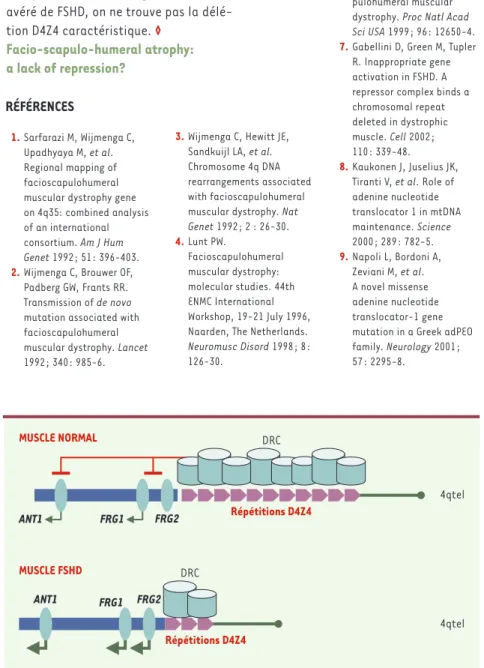
Documents relatifs
Aware of the scientific, technologic, industrial and societal issues in order to analyse the evolution of writing systems and graphical forms during a long period (the Middle Ages)
In Section 2 we show how to construct stable PMLs for so-called generalized Lorentz models and derive the energy estimates for the corresponding time-domain system.. In Section 3
Furthermore, NMDAR antibody encephalitis is classi- cally associated with a polymorphic affective psychosis with catatonic features [7], which is broader than the phe-
12978.. Sch ¨onhardt et al. Title Page Abstract Introduction Conclusions References Tables Figures ◭ ◮ ◭ ◮ Back Close. Full Screen
In this study, we chose not to examine the changes that have occurred with the arrival of PLF tools (to do so, we would have had to conduct interviews and observations before
Par la langue d’Hermès mes pas sont assurés Je côtoie les mystères sur l'arbre des secrets La pythie me transmet le langage des dieux Pour assembler l'épart et construire ce lieu
Yà aussi Hèra la femme de Zeus qui aime pas du tout quand yà son mari qui bisoute des autres Déaisses ou des morts avec des ailes (des morts d’ailes je crois parce que j’ai pas
We report here the bioanalysis of the stay green QTL-2 region based on synteny between rice chromosome 1 and sorghum chromosome 3 and assessment of the allelic diversity of
