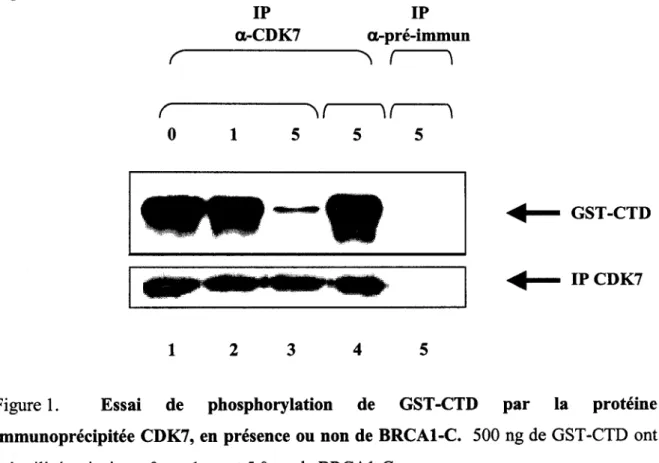
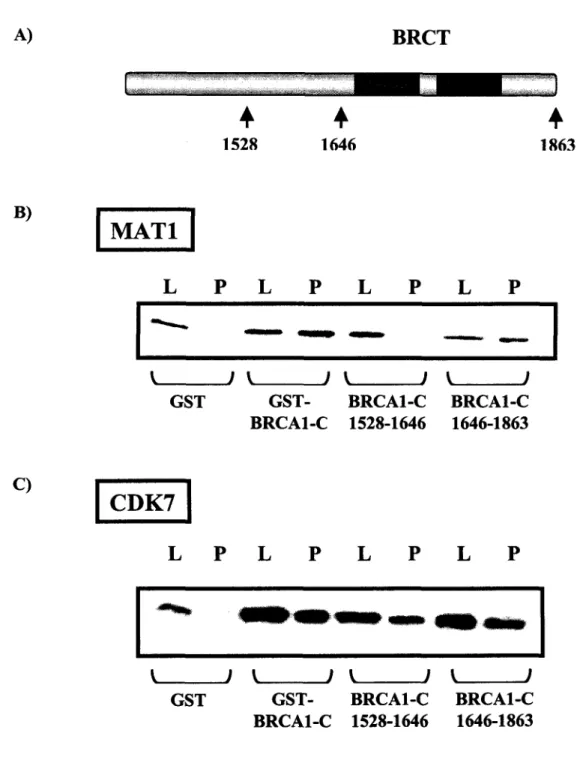
Interactions physiques et fonctionnelles entre la région C-terminale de la protéine BRCA1 et la kinase CDK7
60
0
0
Texte intégral
Figure


Documents relatifs