Calsenilin Is Required for Endocrine Pancreas Development in Zebrafish
V. Stetsyuk,1 B. Peers,2 A. Mavropoulos,3 V. Verbruggen,2 B. Thisse,3 C. Thisse,3 P. Motte,4 B. Duvillié,1 and R.
Scharfmann1
1University Paris-Descartes, Faculty of Medicine; INSERM, Necker Hospital, U845/EMI 363, Paris, France
2Laboratoire de Biologie Moléculaire et de Génie Genetique, Center of Biomedical Integrative Genoproteomics (CBIG), Liege (Sart-Tilman),
Belgium
3Institut de Génétique et de Biologie Moléculaire et Cellulaire, UMR 7104, CNRS/INSERM/ULP, Illkirch, France
4Laboratoire de Biologie Cellulaire Végétale, Cellule d'Appui Technologique en Microscopie (CAT1), Université de Liège, Liège, Belgium Abstract
Calsenilin/DREAM/Kchip3 is a neuronal calcium-binding protein. It is a multifunctional protein, mainly expressed in neural tissues and implicated in regulation of presenilin processing, repression of transcription, and modulation of A-type potassium channels. Here, we performed a search for new genes expressed during
pancreatic development and have studied the spatiotemporal expression pattern and possible role of calsenilin in pancreatic development in zebrafish. We detected calsenilin transcripts in the pancreas from 21 somites to 39 hours postfertilization stages. Using double in situ hybridization, we found that the calsenilin gene was expressed in pancreatic endocrine cells. Loss-of-function experiments with anti-calsenilin morpholinos demonstrated that injected morphants have a significant decrease in the number of pancreatic endocrine cells. Furthermore, the knockdown of calsenilin leads to perturbation in islet morphogenesis, suggesting that calsenilin is required for early islet cell migration. Taken together, our results show that zebrafish calsenilin is involved in endocrine cell differentiation and morphogenesis within the pancreas.
Key words: pancreas; calsenilin; development; zebrafish
INTRODUCTION
The vertebrate pancreas is a mixed exocrine and endocrine gland that plays a crucial role in metabolism. The exocrine pancreas produces and secretes digestive enzymes released into the duodenum through the pancreatic ductal system. The endocrine tissue of the pancreas is organized in individual islets of Langerhans scattered within the exocrine tissue. Islets of Langerhans are composed of four types of differentiated endocrine cells (alpha, beta, delta, and PP cells that produce glucagon, insulin, somatostatin, and pancreatic polypeptide, respectively).
In rodents, the pancreas originates from the dorsal and ventral regions of the foregut endoderm directly posterior to the stomach (Pictet and Rutter, 1972; Slack, 1995). In the past decade, progress has been made in identifying transcription factors and signaling molecules that control pancreatic development (Grapin-Botton and Melton, 2000; Scharfmann, 2000; Kim and Hebrok, 2001; Edlund, 2002). In the mouse, the endodermal region
committed to a pancreatic fate first expresses the homeodomain containing transcription factor Pdx1 (Pancreatic
duodenal homeobox-1). During adulthood, Pdx1 expression becomes largely confined to beta cells where it
activates insulin gene transcription (Ohlsson et al., 1993). Targeted disruption of the Pdx1 gene in mice leads to pancreas agenesis revealing the crucial role of Pdx1 in pancreas development (Ahlgren et al., 1996; Offield et al., 1996). The demonstration of the crucial role of Pdxl in pancreas development was followed by gene targeting of many other genes revealing the functional importance of several transcription factors at various stages of pancreas development. These factors include the Lim homeodomain protein Isl-1 (Ahlgren et al., 1997); the paired box genes Pax4 and Pax6 (Sosa-Pineda et al., 1997; StOnge et al., 1997); the bHLH genes NeuroD and
Neurogenin3 (Naya et al., 1997; Grad-wohl et al., 2000); the NK homeobox genes Nkx2.2, Nkx6.1, Nkx6.2
(Sussel et al., 1998; Sander et al., 2000; Henseleit et al., 2005); and the homeobox gene Hb9 (Harrison et al., 1999; Li et al., 1999). Pancreas development is also tightly controlled by signaling molecules derived from adjacent mesodermal structures. This finding is the case for signals derived from the notochord such as activin and Fgf2 (Kim et al., 1997; Hebrok et al., 1998), from the dorsal aorta such as sphingosine-1-phosphate (Lam-mert et al., 2001; Edsbagge et al., 2005), and from the mesenchyme such as Fgf10 (Bhushan et al., 2001). Despite the increasing knowledge of the genetic cascade occurring during pancreatic development, it can be postulated that additional transcription factors and signaling molecules controlling pancreas development need to be discovered. This point is supported by the fact that, in human, specific forms of abnormal pancreas
development cannot be explained by mutations in genes known to control pancreas development (Mitchell et al., 2004). Moreover, current knowledge on pancreas development is not sufficient to recapitulate beta cell
development from embryonic stem cells (Ra-jagopal et al., 2003; Hansson et al., 2004).
Recently, investigators have described and used new experimental models such as zebrafish (Argenton et al., 1999) and medaka (Assouline et al., 2002) to further dissect pancreas development. In zebrafish, as in rodents, the pancreas emerges from two primordia (Biemar et al., 2001). The dorsal bud first appears at 24 hours postfertilization (hpf). Next, the second bud forms at 34 hpf ventrally and anteriorly to the first one. Finally, the two pancreatic buds merge to form a unique organ at 44 hpf (Field et al., 2003). The dorsal bud differentiates into endocrine cells, whereas the second bud gives rise to the pancreatic ducts, exocrine cells, and some endocrine cells (Field et al., 2003). The regulatory mechanisms controlling pancreas development seem to be conserved among vertebrates. Indeed, the main transcription factors crucial for pancreas development in mammals, such as Pdx-1, Isl1, Nkx2.2, and Pax6.2, are expressed during pancreas development in zebrafish (Biemar et al., 2001), and morpholino-mediated gene knock down in zebrafish of many of these transcription factors, such as pdx-1, has resulted in similar pancreatic phenotypes in mammals and zebrafish (Huang et al., 2001; Yee et al., 2001). Thus, new genes discovered in zebrafish are of potential interest for mammalian studies. In the present study, we focused on the role of Calsenilin/DREAM/ KChIP3 (csen) in pancreas development in zebrafish. Calsenilin is a multifunctional protein mainly expressed in the brain where it binds presenilin and modulates some of its effects. Calsenilin also binds the dynorphin response element and regulates Dynorphin expression and nociception. Finally, Calsenilin binds potassium channels and regulates potassium conductance (Buxbaum, 2004).
Here, we have studied the expression and function of zebrafish calsenilin in pancreas development. We analyzed the spatiotemporal expression pattern of calsenilin in embryonic pancreas. Then, we performed loss-of-function experiments with anti-calsenilin morpholinos and demonstrated that calsenilin is required for endocrine cell differentiation.
RESULTS
Calsenilin Is Expressed in the Pancreas of Zebrafish During Development
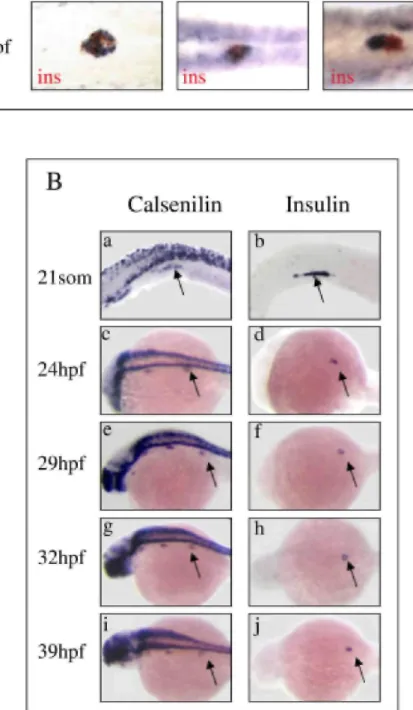
As a first step in searching for new genes expressed in the endocrine pancreas in zebrafish, we further analyzed different clones identified within the course of a large scale in situ hybridization screen whose expression, based on whole-mount in situ hybridization, suggested that they were specific for the endocrine pancreas in zebrafish (Thisse et al., 2004). We cohybridized zebrafish at 24 hpf with probes recognizing the gene to be tested and
insulin. Figure 1A is representative of the type of pattern we observed. In some cases, in situ hybridization
demonstrated costaining between the clone to be tested and insulin. This costaining is, for example, the case for a clone (sequence accession no. BC059672), which displays strong sequence similarities with the mouse Reprimo (TP53-dependent G2 arrest mediator candidate) and for a clone (sequence accession no. BC059629)
corresponding to calsenilin (Fig. 1A, top and middle panels). In other cases, the clone to be tested and insulin were not expressed in the same cell types, but in adjacent cells. This finding is the case for a clone (sequence accession no. BU670721) that corresponds to the gene coding for the regulator of G-protein signaling 4 (Fig. 1A, right panel). In the present work, we focused on the clone that codes for calsenilin.
To characterize more precisely the temporal expression of calsenilin in the pancreas, whole-mount in situ hybridization with digoxigenin (DIG) -labeled probes was performed at stages 18-, 21-somites, and 24, 29, 33, and 39 hpf (Fig. 1B, left panel). At the 18-somite stage, calsenilin transcripts were detected in the neural tube but not in the pancreatic domain (not shown). The first calsenilin transcripts detected in pancreatic endoderm were found at the 21-somite stage (Fig. 1B, panel a). At this stage, the same region of the embryo was positive for
calsenilin and insulin (Fig. 1B, compare panels a and b). Expression of calsenilin was also detected in
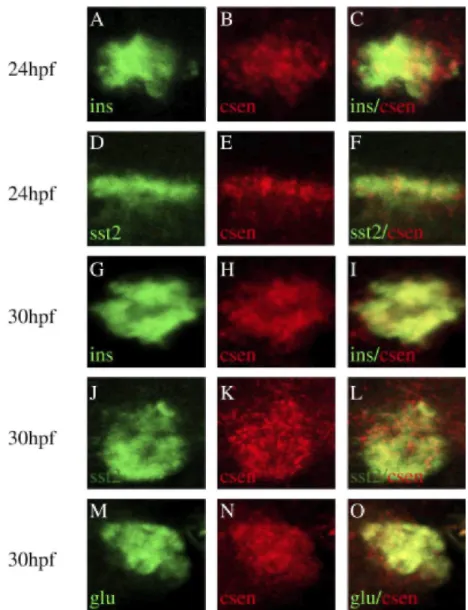
insulin-positive cells at 24 hpf (Fig. 1B, panels c and d). Calsenilin persisted in insulin-insulin-positive cells until at least 39 hpf (Fig. lB).We next carried out fluorescent double in situ hybridization using a calsenilin probe in combination with insulin, glucagon, or somatostatin2 probes. Optical sections of hybridized embryos at 24 hpf and 30 hpf are presented in Figure 2. At 24 hpf, calsenilin colocalized with insulin (Fig. 2A-C) and somatostatin (Fig. 2D-F). At 30 hpf, insulin-, somatostatin2-, and glucagon-positive cells expressed calse nilin (Fig. 2G-L). Taken together, these results indicate that, during pancreas development in zebrafish, calsenilin is expressed in all pancreatic endocrine cell types.
Fig. 1. Expression of calsenilin in the zebrafish pancreas. A: Search for new genes expressed in the pancreas: zebrafish at 24 hours postfertil-ization (hpf) were cohybridized with digoxigenin-labeled antisense probes (in blue) corresponding to the gene to be tested and fluorescein-labeled antisense insulin probe (in red). Calsenilin that was coexpressed with insulin was kept for further analysis. B: Expression of calsenilin during zebrafish development. Whole-mount in situ hybridization was performed with digoxigenin-labeled antisense calsenilin and insulin probes at different stages of development (21 -somites, 24 hpf, 29 hpf, 32 hpf, 39 hpf). Arrows point to insulin-positive regions.
Fig. 2. Coexpression of calsenilin and pancreatic hormones. Double fluorescent in situ hybridization in zebrafish embryos. Zebrafish at 24 hours postfertilization (hpf) and 30 hpf were cohybridized with digoxigenin-labeled antisense calsenilin probe in combination with either DNP-labeled antisense insulin, glucagon, or somatostatin2 probes. Calsenilin was revealed with tyramide-Cy3 substrate (in red) while insulin, glucagon, and somatostatin2 were revealed with tyramide-fluorescein isothio-cyanate (in green). Images on the right column represent superposition of the respective left and middle confocal panels.
Calsenilin Expression Is Downstream of Retinoic Acid and Notch Signaling
In zebrafish, pancreas development is controlled by different pathways such as the retinoic acid (RA) and Notch signaling pathways. We have, therefore, investigated the relationships between calsenilin and these signaling pathways.
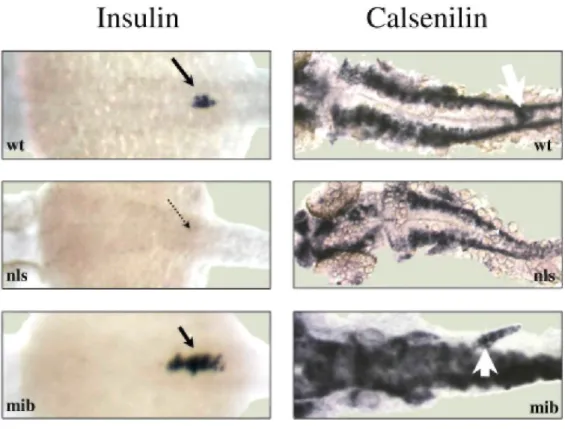
Recent data have shown that retinoic acid signaling is essential for pancreas specification (Stafford and Prince, 2002; Chen et al., 2004; Stafford et al., 2004). A specific mutation in zebrafish, called neckless, inactivates retinaldehyde dehydrogenase type 2, an enzyme involved in retinoic acid biosynthesis. The embryos carrying the
neckless mutation (nls) are characterized by a truncation of the anteroposterior axis, located anterior to the
somites, and by a lack of expression of pancreatic and endocrine markers (Begemann et al., 2001; Stafford and Prince, 2002). We have used nls mutants at 24 hpf as a model of RA loss-of-function. In such mutants, we analyzed the expression pattern of calsenilin and insulin by in situ hybridization. We found that calsenilin was expressed in nonpancreatic tissues in both wild-type and nls embryos, that is, in neural tube (Fig. 3) and central nervous system. On the other hand, calsenilin and insulin transcripts were completely absent from the pancreatic endoderm in nls mutants (Fig. 3).
Recent data also indicate that the Notch signaling pathway controls pancreatic cell development. In mouse and zebrafish, inactivation of genes involved in Delta/Notch-mediated lateral specification, such as Dll-1, RBP-Jk, or
Hes1, leads to an acceleration of endocrine cell differentiation (Apelqvist et al., 1999; Jensen et al., 2000; Hald et
al., 2003; Miyamoto et al., 2003; Esni et al., 2004). In zebrafish, animals carrying homozygous mutations at the
mindbomb locus (mibta52b) lack a ubiquitin ligase required for normal posttranslational processing and trafficking
of Delta, resulting in defective Notch pathway activation (Itoh et al., 2003). Recent data indicate that, in mib mutant embryos, pancreatic endocrine differentiation is perturbed, leading to an increase in the number of
insulin- and somαtostα-fire-expressing cells and a lack of glucagon-expressing cells (Zecchin et al., 2007). To
investigate the effect of the Notch pathway on cαlsenilin expression, we performed in situ hybrization experiments in mib mutants and analyzed insulin and cαlsenilin expression. We detected an up-regulation of
insulin expression in the pancreas and of cαlsenilin in the pancreas as well as in the neural tube (Fig. 3) of the 24
hpf mutants. This result demonstrates that the Notch pathway affects negatively the expression of cαlsenilin. Taken together, these results show that cαlsenilin expression occurs downstream of the retinoic acid and the Notch pathways.
Role of Calsenilin in Pancreatic Endocrine Cell Development
To gain insight into the role of cαlsenilin during pancreas development, we examined the effect of its inactiva-tion. For this purpose, two different antisense morpholinos were used: MO-calsenilin and M03-calsenilin that target the first ATG of the coding sequence and the 5'-untranslated region (UTR) of calsenilin, respectively. As a negative control, we used 5-misMO-calsenilin, a 5-bp mismatch version of the MO-calsenilin morpholino. We tested the ability of the morpholino to block translation using an in vitro translation assay. As shown in Figure 4A, both MO-calsenilin and M03-calsenilin decreased calsenilin translation, when compared with control 5-misMO-calsenilin morpholino. We next injected approximately 2.5 ng of morpholino into one- to two-cell stage embryos. First, we raised injected embryos until 24 hpf and analyzed their morphology. Neither control nor morphant embryos demonstrated gross evidence of developmental delay during the first 24 hpf, as assessed by eye pigmentation and somite formation. Then, the effect on pancreas development was assessed by in situ hybridization to detect the presence or absence of various pancreatic markers.
At 24 hpf, MO-calsenilin morphants had a selective loss of insulin-expressing cells as compared with wild-type or 5-misMO-calsenilin control embryos. The same phenotype was also observed for M03-calsenilin morphants (Fig. 4B). Quantification of the number of insulin-positive cells showed a significant difference between MO-calsenilin-treated embryos and controls. Indeed, MO-calsenilin (n = 21) and M03-calsenilin (n = 9) morphants displayed a reduction of 48.4% (P < 0.0001) and 47% (P < 0.0001) of insulin-positive cells, respectively, as compared with 5-misMO-calsenilin control embryos (Fig. 4B).
We next defined the effect of cαlsenilin inactivation on the development of other pancreatic cell types. We found that, at 24 hpf, the number of somatostatin2-expressing cells was reduced by 35.7% (P < 0.0001) as compared with 5-misMO-calsenilin embryos (Fig. 4C). Expression of pdx1, a marker of pancreatic progenitors, was not significantly affected in morphants as compared with control 5-misMO-calsenilin (Fig. 4C). We finally analyzed the effect of calsenilin inactivation on the expression of isl1 and pax6.2, two genes implicated at early stages of pancreatic endocrine cell development. No significant effect in MO-calsenilin morphants could be observed on the expression of pax6.2, whereas the number of isl1-positive cells in MO-calsenilin morphants decreased by 34.5% (P < 0.01) with respect to wild-type control embryos (Fig. 4C).
In addition to a decrease in the number of insulin- and somatostatin2-expressing cells that developed, morphants also gave rise to defects in pancreatic islet morphogenesis. Indeed, in MO-calsenilin-injected embryos, the
insulin-expressing cells failed to cluster and remained dispersed (Fig. 4B). This effect was also clearly seen
when other pancreatic markers such as isll and pax6.2 (Fig. 4C) were analyzed. The number of embryos with disorganized pancreatic islets visualized by insulin or somatostatin2 expression was next quantified. In MO-calsenilin morphants, 16 of 25 zebrafish had dispersed insulin-expressing cells (64%), a phenotype found in only 1 of 11 of the 5-misMO-calsenilin control embryos (9%). Moreover, 5 of 19 MO-calsenilin morphants had dispersed somatostatin2-expressing cells, whereas this was found in only 1 of 9 of the 5-misMO-calsenilin control embryos. At 40 hpf, development of insulin-positive cells remained perturbed (Fig. 4B, bottom panels), suggesting that loss of calsenilin is effecting endocrine cell specification. Taken together, the dramatic decrease in the number of insulin- and somatostatin -expressing cells in knockdown embryos indicates that calsenilin plays a crucial role in pancreatic endocrine cell development and islet morphogenesis.
DISCUSSION
In the present work, we demonstrate that, in zebrafish, calsenilin is expressed in pancreatic endocrine cells. Loss-of-function experiments using morpholino indicate that calsenilin plays a crucial role in pancreatic endocrine cell development and islet morphogenesis.
We first described the spatiotemporal expression pattern of calsenilin in zebrafish. Calsenilin is mainly
expressed in specific regions of the brain such as telencephalon, diencephalon, and hindbrain. It is also expressed in the spinal chord, in cranial ganglia, and in the pancreas. In the pancreas, expression of calsenilin was not detected at the 18-somite stage but observed at the 21-somite stage and persisted until at least 39 hpf. At these stages, calsenilin was expressed in all pancreatic endocrine cell types: insulin-, glucagon-, and
somatostatin2-expressing cells. Pancreatic endocrine cell development in zebrafish has been previously described in detail. The
first insulin-, somatostatin-, and glucagon-expressing cells appear at 12-somite, 16-somite, and 24 hpf stages, respectively (Argenton et al., 1999; Biemar et al., 2001). Thus, the detection of insulin and somatostatin cells in the developing pancreas appears earlier than calsenilin expression and strongly suggests that calsenilin is not a marker of the early endoderm, but is rather expressed in late differentiation processes of pancreatic endocrine cells in zebrafish. Very recently, Calsenilin expression was examined in mouse pancreatic islets, and it was shown that, in rodent adult pancreas, Calsenilin is expressed in both glucagon- and insulin-positive cells (Jacob-son et al., 2006). Our present findings further support that calsenilin is expressed in the pancreatic endocrine cells and suggests that Calsenilin may have a conserved role in mouse and zebrafish.
Calsenilin belongs to a family of proteins that includes four members: KChIPl, KchIP2, calsenilin/KchIP3/ DREAM, and CALP/KchIP4 (Bux-baum, 2004). In rodents, three major functions have been defined for Calsenilin. First, Calsenilin is involved in apoptosis (Buxbaum et al., 1998). In the brain, calsenilin enhances apoptosis by altering endoplasmic reticulum calcium signaling (Lilliehook et al., 2002). Calsenilin also acts as a calcium-regulated transcriptional repressor and modulates the expression of different genes, such as Dynorphin (Carrion et al., 1999). Finally, Calsenilin interacts with potassium channels and modulates their function (An et al., 2000). The vast majority of these functions have been described in the brain, and rare information concerns other tissues. Very recently, it was shown that Calsenilin is expressed in pancreatic beta cells, where it could be implicated in endocrine cell function. Specifically, it was shown that Calsenilin interacts with the prodinorphin promoter and, using islets from calsenilin-deficient mice, it was demonstrated that calsenilin inhibits
prodynorphin transcription in beta cells (Jacobson et al., 2006). Taken together, such results demonstrate that prodynorphin is a direct target of calsenilin. It was also shown that islets from calsenilin-deficient mice secrete
more glucagon than wild-type islets (Jacobson et al., 2006). A working hypothesis is that calsenilin would control prodynorphin expression in beta cells, which would affect glucagon expression neighboring alpha cells. In the present study, we have investigated the role of calsenilin during pancreas development in zebrafish using morpholinos. Our knockdown experiments demonstrate that calsenilin controls pancreatic endocrine cell development in a positive fashion. Indeed, embryos injected with antisense morpholinos directed against
calsenilin had a dramatic reduction of the number of insulin- and somatostatin2-expressing cells with respect to
control. The mechanism by which calsenilin controls pancreatic endocrine cell development is not yet elucidated, but different hypotheses can be proposed. In rodents, the final number of beta cells that develop is controlled by a balance between proliferation, differentiation, and apoptosis (Finegood et al., 1995), and it can be postulated that the same type of control occurs in zebrafish. One first hypothesis, based on the known roles of
Calsenilin in the brain would be that Calsenilin controls apoptosis in pancreatic endocrine cells. In that case, an
increase in apoptosis should occur in zebrafish embryos injected with antisense morpholinos directed against
calsenilin to explain the observed phenotype. This first hypothesis seems unlikely. Indeed, in rodents, Calsenilin has a pro-apoptotic role (Lilliehook et al., 2002; Jo et al., 2004), and we would have expected a
decrease in apoptosis and, thus, an increase in endocrine cell development in zebrafish that lack calsenilin, if
calsenilin activated apoptosis in the pancreas, as is the case in the brain. A second hypothesis would be that calsenilin controls either pancreatic cell proliferation or differentiation. In this context, it is extremely interesting
to note that recent data indicate that overexpression of Calsenilin enhances gamma-secretase activity (Jo et al., 2005). Such enzymatic activity cleaves within the transmembrane region of the amyloid precursor protein and represents a critical limiting step to generate amyloid beta peptide (Selkoe, 2001). Gamma secre-tase activity is also necessary for the cleavage of Notch, releasing the Notch intracellular domain implicated in cell fate determination in many tissues (Selkoe, 2001). During development, the Notch pathway plays a major role in the control of pancreatic cell proliferation and differentiation. It could, thus, be speculated that calsenilin interacts in the pancreas with this pathway to control beta cell development. Our results also indicate that, in zebrafish embryos injected with anti-sense morpholinos directed against calsenilin, pancreatic endocrine cells are found dispersed in contrast to control zebrafish where endocrine cells associate, forming an islet of Langer-hans.
During zebrafish development, islets form by aggregation of "migrating" endocrine cells, as is the case in mice (Deltour et al., 1991) rather than by clonal growth of individual progenitors. The pancreatic hormone-expressing cells first appear scattered within the prepatterned, pdx-1-posi-tive endodermal epithelium between the 12- and 18-somite stages and associate to form an islet between the 18-somites and 24 hpf stages (Biemar et al., 2001). Our data indicate that, in the absence of calsenilin, the ability of endocrine cells to migrate and form a pancreatic islet is perturbed. Several mutants affecting cell migration in the pancreas of zebrafish have been described. One example is the Minime mutation, which displays a reduced insulin expression and dispersed endocrine cells as in the calsenilin morphant embryos (Kim et al., 2006). Another example is represented by the knypek gene that is involved in cell convergence within the pancreas (Marlow et al., 1998; Biemar et al., 2001). The mechanisms by which calsenilin controls cell migration in the pancreas and the possible interaction with such migratory proteins should be of great interest to define the mechanisms of pancreas morphogenesis.
Recently, mice deficient in Calsenilin expression have been generated (Cheng et al., 2002; Lilliehook et al., 2003). While, such mice were used to analyze in detail the role of calsenilin in pancreatic islet cell function, neither quantitative morphometric analysis of islet cells nor islet morphogenesis was reported (Jacobson et al., 2006). In the present study, we have demonstrated that Calsenilin plays an important role in the terminal differentiation of endocrine cells. A quantitative analysis of islet cells in Csen-/- mice will now be important to define the role of Calsenilin in the development of the rodent pancreas.
Fig. 3. Expression of calsenilin in mutants with affected retinoic acid synthesizing aldehyde dehydrogenase
(RALDH; nls) and Notch (mib) signaling. Whole-mount in situ hybridization with digoxigenin-labeled antisense insulin and
calsenilin probes on wild-type (WT), nls mutant defective in retinoic acid (RA) synthesis and mib mutants defective in the Delta-Notch
pathway. Black and white arrows indicate insulin and calsenilin expression in the pancreas, respectively.
morpholino antisense oligonucleotides directed against calsenilin. B: Whole-mount in situ hybridization on embryos injected with MO-calsenilin, M03-calsenilin or with 5-misMO-calsenilin control antisense oligonucleotides. Zebrafish embryos at 24 hours post-fertilization (hpf) or 40 hpf were hybridized with a digoxigenin-labeled antisense insulin probe. Quantification of insulin-positive cells either in 5-misMO-calsenilin, MO-calsenilin, or M03-calsenilin morphants at stage 24 hpf. In knockdown embryos, the number of insulin-expressing cells was strongly decreased. ***P < 0.0001 C: Whole-mount in situ hybridization with digoxigenin-labeled antisense sst2, pdx1, isl1, and
pax6.2 in MO-calsenilin and control 5-misMO-calsenilin embryos at 24 hpf. Note that in MO-calsenilin morphants, pancreatic endocrine
cells were dispersed and did not associate to form an islet. Quantification of sst2-, is/7-, and Pax6.2-positive cells in wild-type or MO-calsenilin morphants at stage 24 hpf. *P < 0.01 ; ***P < 0.0001
Zebrafish Maintenance and Mutant Lines Used
Zebrafish (Danio rerio) were raised and cared for according to established standard rules and procedures (Westerfield, 1995). Wild-type embryos from the AB strain were used, and the embryos were staged according to Kimmel et al. (1995). Homozygous mutants were obtained by mating heterozygous fishes for the mindbomb,
mibta52b (Haddon et al., 1998), and neckless, nlsi26 (Begemann et al., 2001), alleles, respectively.
Clones and Hybridization Probes
The clones screened are issued from different cDNA libraries. The cDNA clone corresponding to the calsenilin (sequence accession no.BC059629) as well as the clone that displays strong sequence similarities with the mouse
Reprimo (TP53-dependent G2 arrest mediator candidate; sequence accession no. BC059672) derive from a
zebrafish adult retina cDNA library constructed in a lambda ZAPII vector. The clone corresponding to the gene coding for the regulator of G-protein signalling 4 (sequence accession no. BU670721) and has been isolated from a cDNA library made from RNA of 26-somite embryos and cloned into a pSPORTl vector (Thisse et al., 2004).
The calsenilin probe was generated by polymerase chain reaction from the calsenilin cDNA cloned in pBSK(-) with the following primers: Forward, AACAGCTATGACCATGATTAC-3'; Reverse,
5'-GTAAAACGACGGCC-AGT-3'. The amplified fragments contained T7 polymerase site for anti-sense riboprobe synthesis. The other probes used were as follows: pdx-1 and preproinsulin (Milewski et al., 1998),
glucagon (Argenton et al., 1999), somatostatin2 (Devos et al., 2002), isl1 (Korzh et al., 1993), nkx2.2 (Barth and
Wilson, 1995), and pax6.2 (Krauss et al., 1991).
Labeled antisense riboprobes were synthesized by in vitro transcription using either T7, T3, or SP6 polymerases. For in situ hybridization, zebrafish calsenilin, pdx1, paxG.2, nkx2.2, isl1, and somatostatin2 probes were synthesized with the DIG RNA labeling mix (Roche, France). Insulin riboprobes were synthesized either with the DIG RNA labeling mix or with fluorescein RNA labeling mix (Roche). Insulin, somatostatin2, and glucagon probes used in double fluorescent in situ hybridization were synthesized in the presence of DNP-11-UTP ribonucleotides (TSAi Plus system, Perkin Elmer, France) as previously described (Mavropoulos et al., 2005). Whole-Mount In Situ Hybridization on Zebrafish Embryos
Single whole-mount in situ hybridization was carried out according to established procedures (Schulte-Merker et al., 1992) on wild-type and mutant embryos with a 65°C overnight hybridization step. Double whole-mount in situ hybridization was performed as described (Jowett, 2001) at 65°C. Colorimetric revelations were performed with BCIP (Promega) and either NBT (Promega) for DIG-UTP or INT for fluorescein-UTP, to obtain a blue or red staining, respectively. Embryos were mounted in 87% glycerol in phosphate buffered saline and conserved for following analysis.
Fluorescent in situ hybridization was performed as previously described (Mavropoulos et al., 2005). Briefly, embryos were treated with 2% H2O2 during 40 min to inactivate endogenous peroxidase, just before proteinase K
digestion. Hybridization was carried out overnight at 70°C. For immunodetection, preabsorbed horseradish peroxidase (HRP) -coupled an-tidigoxigenin (Roche) or HRP-coupled anti-DNP antibodies (Perkin Elmer) were used followed by extensive washes in TNT buffer. Revelation was performed by incubating embryos in
tyramide-fluorescein isothiocyanate and tyramide-Cy3 prepared at a final dilution of 1/50 in 1x Amplification Reagent (Perkin Elmer; Mavropoulos et al., 2005).
Morpholino Injections in Zebrafish
The morpholino antisense oligonucleotides obtained from Gene Tools were as follows: MO-calsenilin, CCCTTCT-GTCCATCTGCCTGCATG, which targets the first ATG of the coding sequences; M03-calsenilin, GATCCT-CAGATGTTCGCTCTCATTC, which targets the 5'-UTR; and 5-misMO-calsenilin,
CCGTTCTCTCCATGTG-GCTCCATG, which is a 5-bp mismatch version of the MO-calsenilin morpholino. To test the in vitro efficacy of morpholino antisense oligonucleotides directed against calsenilin, mRNA was transcribed from the plasmid construct Calsenilin/ZAP II described above and translated using a TNT T7/ T3-coupled reticulocyte lysate system (Promega) according to the specifications of the manufacturer. Briefly, the DNA template (500 ng), TNT rabbit lysate, reaction buffer, amino acid mixture deficient in methionine (20 µM), methionine 35S (0.8 mCi/ml; GE Healthcare, USA), Rnasin (0.8 U/µl), T3 RNA polymerase, and morpholinos
were combined in a total volume of 25 µl and incubated at 30°C for 90 min. The results of the translation reaction were assessed by sodium dodecyl sul-fate-polyacrylamide gel electrophoresis and autoradiography. For injection, morpholinos were reconstituted in Danieau's buffer (Westerfield, 1995) as 16 mM stock solutions. The stocks were diluted to working concentrations of 1-8 mM in Danieau solution for injection. Rhodamine dextran at 0.5% was added to the samples to check injection efficiency. Approximately 1 nl of working solution of morpholinos was injected into the yolk of one-cell stage wild-type embryos using published procedures (Westerfield, 1995). For quantification, yolk-free embryos were mounted on slides and slightly squashed with cover glass to obtain a monolayer of hormone-positive cells.
Image Acquisition
Whole embryos were photographed by using a dissecting microscope equipped with a Canon digital camera. Digital images were processed using Adobe Photoshop software. Confocal imaging was performed by using a Leica TCS SP2 inverted confocal laser microscope and digitized as previously described (Mavropoulos et al., 2005).
ACKNOWLEDGMENTS
We thank Annie Basmaciogullari for help with in situ hybridization. V. Stetsyuk received support from the Association "l'Aide aux Jeunes Diabétiques." This work was funded by the National Center for Research Resources, a component of the National Institutes of Health.
Grant sponsor: Institut National de la Santé et de la Recherche Médicale, INSERM-JDRF Grant (AIP Cellules souches A03139MS);
Grant sponsor: French National Program of Research on Diabetes;
Grant sponsor: Association Française des Diabétiques; Grant sponsor: Centre National de la Recherche Scientifique;
Grant sponsor: NIH: Grant number: ROI RR15402.
REFERENCES
Ahlgren U, Jonsson J, Edlund H. 1996. The morphogenesis of the pancreatic mesenchyme is uncoupled from that of the pancreatic epithelium in IPFl/PDXl-defi-cient mice. Development 122:1409-1416.
Ahlgren U, Pfaff SL, Jessell TM, Edlund T, Edlund H. 1997. Independent requirement for ISL1 in formation of pancreatic mesenchyme and islet cells. Nature 385: 257-260.
An WF, Bowlby MR, Betty M, Cao J, Ling HP, Mendoza G, Hinson JW, Mattsson KI, Strassle BW, Trimmer JS, Rhodes KJ. 2000. Modulation of A-type potassium channels by a family of calcium sensors. Nature 403:553-556.
Apelqvist A, Li H, Sommer L, Beatus P, Anderson DJ, Honjo T, Hrabe de Angelis M, Lendahl U, Edlund H. 1999. Notch signalling controls pancreatic cell differentiation. Nature 400:877-881.
Argenton F, Zecchin E, Bortolussi M. 1999. Early appearance of pancreatic hormone-expressing cells in the zebrafish embryo. Mech Dev 87:217-221.
Assouline B, Nguyen V, Mahe S, Bourrat F, Scharfmann R. 2002. Development of the pancreas in medaka. Mech Dev 117: 299-303. Barth KA, Wilson SW. 1995. Expression of zebrafish nk2.2 is influenced by sonic hedgehog/vertebrate hedgehog-1 and demarcates a zone of neuronal differentiation in the embryonic forebrain. Development 121:1755-1768.
Begemann G, Schilling TF, Rauch GJ, Gei-sler R, Ingham PW. 2001. The zebrafish neckless mutation reveals a requirement for raldh2 in mesodermal signals that pattern the hindbrain. Development 128: 3081-3094.
Bhushan A, Itoh N, Kato S, Thiery J, Czernichow P, Bellusci S, Scharfmann R. 2001. Fgf10 is essential for maintaining the proliferative capacity of epithelial progenitor cells during early pancreatic organogenesis. Development 128:5109-5117.
Biemar F, Argenton F, Schmidtke R, Ep-perlein S, Peers B, Driever W. 2001. Pancreas development in zebrafish: early dispersed appearance of endocrine hormone expressing cells and their convergence to form the definitive islet. Dev Biol 230:189-203.
Buxbaum JD. 2004. A role for calsenilin and related proteins in multiple aspects of neuronal function. Biochem Biophys Res Commun 322:1140-1144.
Buxbaum JD, Choi EK, Luo Y, Lilliehook C, Crowley AC, Merriam DE, Wasco W. 1998. Calsenilin: a calcium-binding protein that interacts with the presenilins and regulates the levels of a presenilin fragment. Nat Med 4:1177-1181.
Carrion AM, Link WA, Ledo F, Mellstrom B, Naranjo JR. 1999. DREAM is a Ca2 + -regulated transcriptional repressor. Nature 398:80-84. Chen Y, Pan FC, Brandes N, Afelik S, Solter M, Pieler T. 2004. Retinoic acid signaling is essential for pancreas development and promotes endocrine at the expense of exocrine cell differentiation in Xenopus. Dev Biol 271:144-160.
Cheng HY, Pitcher GM, Laviolette SR, Whishaw IQ, Tong KI, Kockeritz LK, Wada T, Joza NA, Crackower M, Gon-calves J, Sarosi I, Woodgett JR, Oliveira-dos-Santos AJ, Ikura M, van der Kooy D, Salter MW, Penninger JM. 2002. DREAM is a critical transcriptional repressor for pain modulation. Cell 108:31-43.
Deltour L, Leduque P, Paldi A, Ripoche M, Dubois P, Jami J. 1991. Polyclonal origin of pancreatic islets in aggregation mouse chimaeras. Development 112:1115-1121.
Devos N, Deflorian G, Biemar F, Bortolussi M, Martial JA, Peers B, Argenton F. 2002. Differential expression of two somatostatin genes during zebrafish embryonic development. Mech Dev 115:133-137.
Edlund H. 2002. Pancreatic organogenesis—developmental mechanisms and implications for therapy. Nat Rev Genet 3:524-532.
Edsbagge J, Johansson JK, Esni F, Luo Y, Radice GL, Semb H. 2005. Vascular function and sphingosine-1-phosphate regulate development of the dorsal pancreatic mesenchyme. Development 132: 1085-1092.
Esni F, Ghosh B, Biankin AV, Lin JW, Albert MA, Yu X, MacDonald RJ, Civin CI, Real FX, Pack MA, Ball DW, Leach SD. 2004. Notch inhibits Ptfl function and acinar cell differentiation in developing mouse and zebrafish pancreas. Development 131:4213-4224.
Field HA, Dong PD, Beis D, Stainier DY. 2003. Formation of the digestive system in zebrafish. II. Pancreas morphogenesis. Dev Biol 261:197-208.
Finegood D, Scaglia L, Bonner-Weir S. 1995. Dynamics of β-cell mass in the growing rat pancreas: estimation with a simple mathematical program. Diabetes 44:249-256.
Gradwohl G, Dierich A, LeMeur M, Guillemot F. 2000. neurogenin3 is required for the development of the four endocrine cell lineages of the pancreas. Proc Natl Acad Sci U S A 97:1607-1611.
Grapin-Botton A, Melton DA. 2000. Endoderm development: from patterning to organogenesis. Trends Genet 16:124-130.
Haddon C, Jiang YJ, Smithers L, Lewis J. 1998. Delta-Notch signalling and the patterning of sensory cell differentiation in the zebrafish ear: evidence from the mind bomb mutant. Development 125: 4637-4644.
Hald J, Hjorth JP, German MS, Madsen OD, Serup P, Jensen J. 2003. Activated Notchl prevents differentiation of pancreatic acinar cells and attenuate endocrine development. Dev Biol 260:426-437.
Hansson M, Tonning A, Frandsen U, Petri A, Rajagopal J, Englund MC, Heller RS, Hakansson J, Fleckner J, Skold HN, Melton D, Semb H, Serup P. 2004. Arti-factual insulin release from differentiated embryonic stem cells. Diabetes 53: 2603-2609.
Harrison KA, Thaler J, Pfaff SL, Gu H, Kehrl JH. 1999. Pancreas dorsal lobe agenesis and abnormal islets of Langer-hans in Hlxb9-deficient mice. Nat Genet 23:71-75.
Hebrok M, Kim S, Melton D. 1998. Noto-chord repression of endodermal sonic hedgehog permits pancreas development. Genes Dev 12:1705-1713.
Henseleit KD, Nelson SB, Kuhlbrodt K, Hennings JC, Ericson J, Sander M. 2005. NKX6 transcription factor activity is required for alpha- and beta-cell development in the pancreas. Development 132: 3139-3149.
Huang H, Vogel SS, Liu N, Melton DA, Lin S. 2001. Analysis of pancreatic development in living transgenic zebrafish embryos. Mol Cell Endocrinol 177:117-124.
Itoh M, Kim CH, Palardy G, Oda T, Jiang YJ, Maust D, Yeo SY, Lorick K, Wright GJ, Ariza-McNaughton L, Weissman AM, Lewis J, Chandrasekharappa SC, Chitnis AB. 2003. Mind bomb is a ubiq-uitin ligase that is essential for efficient activation of Notch signaling by Delta. Dev Cell 4:67-82.
Jacobson DA, Cho J, Landa LR, Tamarina NA, Roe MW, Buxbaum JD, Philipson LH. 2006. The downstream regulatory element antagonistic modulator regulates islet prodynorphin expression. Am J Physiol Endocrinol Metab 18:18.
Jensen J, Pedersen EE, Galante P, Hald J, Heller RS, Ishibashi M, Kageyama R, Guillemot F, Serup P, Madsen OD. 2000. Control of endodermal endocrine development by Hes-1. Nat Genet 24:36-44.
Jo DG, Lee JY, Hong YM, Song S, Mook-Jung I, Koh JY, Jung YK. 2004. Induction of pro-apoptotic calsenilin/DREAM/ KChIP3 in Alzheimer's disease and cultured neurons after amyloid-beta exposure. J Neurochem 88:604-611.
Jo DG, Jang J, Kim BJ, Lundkvist J, Jung YK. 2005. Overexpression of calsenilin enhances gamma-secretase activity. Neurosci Lett 378:59-64.
Jowett T. 2001. Double in situ hybridization techniques in zebrafish. Methods 23: 345-358.
Kim SK, Hebrok M. 2001. Intercellular signals regulating pancreas development and function. Genes Dev 15:111-127.
Kim S, Hebrok M, Melton D. 1997. Noto-chord to endoderm signaling is required for pancreas development. Development 124:4243-4252. Kim HJ, Sumanas S, Palencia-Desai S, Dong Y, Chen JN, Lin S. 2006. Genetic analysis of early endocrine pancreas formation in zebrafish. Mol Endocrinol 20: 194-203.
Kimmel CB, Ballard WW, Kimmel SR, Ullmann B, Schilling TF. 1995. Stages of embryonic development of the zebrafish. Dev Dyn 203:253-310.
Korzh V, Edlund T, Thor S. 1993. Zebrafish primary neurons initiate expression of the LIM homeodomain protein Isl-1 at the end of gastrulation. Development 118: 417-425.
Krauss S, Johansen T, Korzh V, Fjose A. 1991. Expression pattern of zebrafish pax genes suggests a role in early brain regionalization. Nature 353:267-270.
Lammert E, Cleaver O, Melton D. 2001. Induction of pancreatic differentiation by signals from bloodvessels. Science 294: 564-567. Li H, Arber S, Jessell TM, Edlund H. 1999. Selective agenesis of the dorsal pancreas in mice lacking homeobox gene Hlxb9. Nat Genet 23:67-70.
Lilliehook C, Chan S, Choi EK, Zaidi NF, Wasco W, Mattson MP, Buxbaum JD. 2002. Calsenilin enhances apoptosis by altering endoplasmic reticulum calcium signaling. Mol Cell Neurosci 19:552-559.
Lilliehook C, Bozdagi O, Yao J, Gomez-Ramirez M, Zaidi NF, Wasco W, Gandy S, Santucci AC, Haroutunian V, Huntley GW, Buxbaum JD. 2003. Altered Abeta formation and long-term potentiation in a calsenilin knock-out. J Neurosci 23: 9097-9106.
Marlow F, Zwartkruis F, Malicki J, Neu-hauss SC, Abbas L, Weaver M, Driever W, Solnica-Krezel L. 1998. Functional interactions of genes mediating convergent extension, knypek and trilobite, during the partitioning of the eye pri-mordium in zebrafish. Dev Biol 203:382-399. Mavropoulos A, Devos N, Biemar F, Zec-chin E, Argenton F, Edlund H, Motte P, Martial JA, Peers B. 2005. sox4b is a key player of pancreatic alpha cell differentiation in zebrafish. Dev Biol 285:211-223.
Milewski WM, Duguay SJ, Chan SJ, Steiner DF. 1998. Conservation of PDX-1 structure, function, and expression in zebrafish. Endocrinology 139:1440-1449.
Mitchell J, Punthakee Z, Lo B, Bernard C, Chong K, Newman C, Cartier L, Desilets V, Cutz E, Hansen IL, Riley P, Poly-chronakos C. 2004. Neonatal diabetes, with hypoplastic pancreas, intestinal atresia and gall bladder hypoplasia: search for the aetiology of a new autosomal recessive syndrome. Diabetologia 47: 2160-2167.
Miyamoto Y, Maitra A, Ghosh B, Zechner U, Argani P, Iacobuzio-Donahue CA, Sri-uranpong V, Iso T, Meszoely IM, Wolfe MS, Hruban RH, Ball DW, Schmid RM, Leach SD. 2003. Notch mediates TGF alpha-induced changes in epithelial differentiation during pancreatic tumori-genesis. Cancer Cell 3:565-576.
Naya F, Huang H-P, Qiu Y, Mutoh H, De-Mayo F, Leiter A, Tsai M-J. 1997. Diabetes, defective pancreatic morphogenesis, and abnormal enteroendocrine differentiation in BETA2/NeuroD-defi-cient mice. Genes Dev 11:2323-2334.
Offield M, Jetton T, Laborsky P, Ray M, Stein R, Magnuson M, Hogan B, Wright C. 1996. PDX-1 is required for pancreatic outgrowth and differentiation of the rostral duodenum. Development 122:983-995.
Ohlsson H, Karlsson K, Edlund T. 1993. IPF1, a homeodomain-containing trans-activator of the insulin gene. EMBO J 12:4251-4259. Pictet R, Rutter W, editors. 1972. Development of the embryonic pancreas. Baltimore: Williams and Wilkins. p 25-66.
Rajagopal J, Anderson WJ, Kume S, Martinez OI, Melton DA. 2003. Insulin staining of ES cell progeny from insulin uptake. Science 299:363.
Sander M, Sussel L, Conners J, Scheel D, Kalamaras J, Dela Cruz F, Schwitzgebel V, Hayes-Jordan A, German M. 2000. Homeobox gene Nkx6.l lies downstream of Nkx2.2 in the major pathway of beta-cell formation in the pancreas. Development 127:5533-5540.
Scharfmann R. 2000. Control of early development of the pancreas in rodents and humans: implications of signals from the mesenchyme. Diabetologia 43: 1083-1092.
Schulte-Merker S, Ho RK, Herrmann BG, Nusslein-Volhard C. 1992. The protein product of the zebraπsh homologue of the mouse T gene is expressed in nuclei of the germ ring and the notochord of the early embryo. Development 116:1021-1032.
Selkoe DJ. 2001. Presenilin, Notch, and the genesis and treatment of Alzheimer's disease. Proc Natl Acad Sci U S A 98: 11039-11041. Slack JM. 1995. Developmental biology of the pancreas. Development 121:1569-1580.
Sosa-Pineda B, Chowdhury K, Torres M, Oliver G, Gruss P. 1997. The Pax4 gene is essential for differentiation of insulin producing β cells in the mammalian pancreas. Nature 386:399-402.
Stafford D, Prince VE. 2002. Retinoic acid signaling is required for a critical early step in zebrafish pancreatic development. Curr Biol 12:1215-1220.
Stafford D, Hornbruch A, Mueller PR, Prince VE. 2004. A conserved role for retinoid signaling in vertebrate pancreas development. Dev Genes Evol 214:432-441.
StOnge L, Sosa-Pineda B, Chowdhury K, Mansouri A, Gruss P. 1997. Pax6 is required for differentiation of glucagon-producing a-cells in mouse pancreas. Nature 387:406-409.
Sussel L, Kalamaras J, Hartigan-O'Connor D, Meneses J, Pedersen R, Rubinstein J, German M. 1998. Mice lacking the home-odomain transcription factor Nkx2.2 have diabetes due to arrested differentiation of pancreatic β cells. Development 125:2213-2221.
Thisse B, Heyer V, Lux A, Alunni V, De-grave A, Seiliez I, Kirchner J, Parkhill JP, Thisse C. 2004. Spatial and temporal expression of the zebrafish genome by large-scale in situ hybridization screening. Methods Cell Biol 77:505-519.
Westerfield M. 1995. The zebrafish book: guide for the laboratory use of zebrafish (Danio Rerio). 3rd ed. Eugene, OR: University of Oregon Press.
Yee NS, Yusuff S, Pack M. 2001. Zebrafish pdxl morphant displays defects in pancreas development and digestive organ chirality, and potentially identifies a multipotent pancreas progenitor cell. Genesis 30:137-140.
Zecchin E, Filippi A, Biemar F, Tiso N, Pauls S, Ellertsdottir E, Gnugge L, Bor-tolussi M, Driever W, Argenton F. 2007. Distinct delta and jagged genes control sequential segregation of pancreatic cell types from precursor pools in zebrafish. Dev Biol 301:192-204.