MELANIE VEILLEUX-NOLLN
Influence de la saison et de la sévérité des feux récents
sur la régénération de l'épinette noire dans la pessière
noire à mousses du Québec
Mémoire présenté
à la Faculté des études supérieures de l'Université Laval dans le cadre du programme de maîtrise en biologie pour l'obtention du grade de maître es sciences (M. Sc.)
DEPARTEMENT DE BIOLOGIE FACULTÉ DES SCIENCES ET DE GENIE
UNIVERSITÉ LAVAL QUÉBEC
2011
Résumé
La saison et la sévérité des feux récents ont été étudiées au sein de la pessière noire à mousses du Québec afin de déterminer leur influence sur la régénération de l'épinette noire (Picea mariana (Mill.) B.S.P.). Pour ce faire, 13 peuplements brûlés au cours des vingt dernières années au printemps ou en été ont été étudiés. Deux indicateurs écologiques ont été utilisés pour caractériser la sévérité des feux récents, soit l'épaisseur de la matière organique résiduelle et le recouvrement des espèces végétales. Un inventaire exhaustif de la régénération de l'épinette noire, sur une superficie totale de 500 m2 par site, a également été
réalisé dans chacun des sites en prenant soin de prélever toutes les plantules et de noter la nature du lit de germination dans lequel elles s'étaient établies. Que le feu ait lieu au printemps ou en été, le sol de tous les peuplements brûlés est couvert d'une épaisse couche de matière organique résiduelle. La matière organique noircie et les éricacées, indices du passage d'un feu léger, abondent dans tous les sites alors que les mousses et le sol minéral nu, qui indiquent le passage d'un feu sévère, couvrent une surface minime. Les deux indicateurs écologiques utilisés dans cette étude suggèrent que tous les feux étudiés ont été légers. La matière organique noircie étant peu favorable à la germination des graines et à la survie des plantules, la régénération de l'épinette noire est déficiente dans la majorité des sites étudiés. Ainsi, ce travail appuie l'hypothèse énoncée par Girard et collaborateurs (2009) selon laquelle les feux légers contribueraient à l'ouverture de la forêt et, ultimement, à l'expansion de la pessière à lichens au cœur de la forêt boréale fermée du Québec.
Remerciements
La première personne à qui je souhaite adresser mes plus sincères remerciements est bien entendu mon directeur de recherche, Dr. Serge Payette, qui m'a accordé sa confiance dès mes premiers pas dans le monde des études graduées. Grâce à son enthousiasme extraordinaire envers l'écologie forestière et à son expertise scientifique, chaque instant passé à ses côtés a été instructif, tant sur le plan professionnel que personnel. Je tiens aussi à remercier les membres de mon comité d'évaluation, Stéphane Boudreau et Julie Turgeon, qui ont assisté à ma présentation de projet, à mon séminaire et qui ont révisé ce manuscrit.
Merci à Ann Delwaide pour ses judicieux conseils et sa grande disponibilité. Merci également à mon assistante Marianne Ricard, pour sa bonne humeur contagieuse, son positivisme et la rigueur de son travail tant sur le terrain qu'en laboratoire. Merci à Yann Gobeil et Bernard Lasnier, deux assistants de courte durée mais dont la contribution à mon projet a fait toute la différence. Merci aussi à Catherine Dumais de s'être aventurée en forêt avec moi à la recherche de sites d'étude.
La camaraderie et l'entraide qui régnent au laboratoire ont rendu ces deux années de maîtrise très agréables et stimulantes. Merci à mes collègues Mathieu Frégeau, Mélanie Jean, Guillaume de Lafontaine, Catherine Plasse, Joannie Savard, Alice Schaffhauser, Francis St-Amour, Alexandre Truchon-Savard, Corinne Vézeau et Marianne White. Un merci spécial à Pierre-Luc Couillard pour son aide sur le terrain, ses conseils éclairés et son sens de l'humour.
Je souhaite remercier du fond du cœur mes parents, qui croient toujours en moi, m'encouragent et me soutiennent constamment dans la poursuite de mes ambitions. Je remercie également mon copain François Lebel qui sait toujours trouver les mots pour m'encourager. Merci François pour ton écoute, tes conseils et ton énorme patience.
Enfin, je remercie le Ministère des Ressources naturelles et de la Faune du Québec de m'avoir fourni les bases de données nécessaires à mon projet ainsi que le Fonds québécois de la recherche sur la nature et les technologies (FQRNT) pour leur soutien financier.
Table des matières
Résumé i Remerciements ii
Liste des figures iv
Liste des tableaux v Liste des annexes vi Introduction 1 Méthodologie 5
Sélection des sites 5 Dispositif expérimental 6 Variables étudiées 6
Caractéristiques de la forêt brûlée 6
Sévérité du feu 8 Régénération 8 Analyses statistiques 9 Sévérité du feu 9 Régénération 11 Résultats 12 Caractéristiques des sites étudiés 12
Structure de taille des peuplements incendiés 13
Historique des feux 13 Sévérité des feux récents 18
Recouvrement des espèces végétales 18 Relation entre la végétation et l'épaisseur de la matière organique 19
Épaisseur de la matière organique résiduelle 20
Régénération de l'épinette noire 21
Discussion 24 Conclusion 29 Bibliographie 30 Annexes 35
Liste des figures
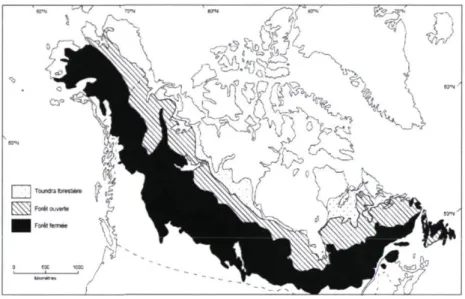
Figure 1. Répartition des trois zones de végétation qui composent la forêt boréale
nord-américaine (Payette 1992) 1 Figure 2. Modèle hypothétique de la régénération de trois catégories différentes de
plantes en fonction de la sévérité du feu : (A) espèces rhizomateuses; (B) espèces à banque de graines; (C) espèces qui dépendent de la dispersion des
graines après feu (Modifiée de Schimmel et Granstrôm 1996) 3 Figure 3. Comparaison du pourcentage de recouvrement (%) moyen de la matière
organique noircie, des éricacées, de la litière, du bois mort, des mousses, des lichens, des sphaignes et du sol minéral selon l'année du feu. Pour un substrat donné, les pourcentages de recouvrement moyen suivis de la même lettre ne sont pas significativement différents, à un seuil de probabilité de 0,05 selon le
test LSD de Fisher 19 Figure 4. Épaisseur (cm) moyenne de la matière organique mesurée sous les mousses, les
lichens, la matière organique noircie et les éricacées. Les épaisseurs mesurées sous les substrats suivis de la même lettre ne sont pas significativement différentes, à un seuil de probabilité de 0,05 selon le test LSD de Fisher 20 Figure 5. Densité (nombre / m ) moyenne des plantules dans les différents lits de
germination, là où au moins une plantule était présente. Données retransformées à l'aide du facteur de correction de Smearing. Les densités chez les substrats suivis de la même lettre ne sont pas significativement différentes, à
un seuil de probabilité de 0,05 selon le test LSD de Fisher 23
Liste des tableaux
Tableau 1. Caractéristiques physiques des sites étudiés 14 Tableau 2. Caractéristiques édaphiques des sites étudiés 16 Tableau 3. TEDF estimé à l'aide de la datation des cernes de croissance et de la
radiodatation des échantillons de charbons 17 Tableau 4. Nombre de plantules d'épinette noire inventoriées par site, sur une superficie
équivalente à 500 m2 22
Tableau 5. Densité à l'hectare (ha) de toutes les tiges formant le peuplement incendié, des tiges issues de graines formant le peuplement et des plantules inventoriées, suivi du rapport de la densité des plantules sur la densité des tiges issues de
Liste des annexes
Annexe 1. Répartition spatiale des treize sites d'étude au Québec, Canada 35 Annexe 2. Distribution de fréquence du nombre d'épinettes noires en fonction du diamètre
à hauteur de poitrine (DHP), accompagnée de la distribution semi-logarithmique du nombre d'épinettes noires en fonction du DHP, avec une
courbe exponentielle négative, pour chacun des sites 36 Annexe 3. Dates 14C des charbons de bois situés à la surface du sol minéral 40
Annexe 4. Pourcentage de recouvrement (%) moyen de la matière organique noircie (M.O. noircie), des éricacées, de la litière, du bois mort, des mousses, des lichens, des sphaignes et du sol minéral dans chacun des sites d'étude. Pour un site donné, les valeurs suivies de la même lettre ne sont pas significativement différentes à
un seuil de probabilité de 0,05 selon le test LSD de Fisher 41 Annexe 5. Corrélation du pourcentage de recouvrement (%) moyen des éricacées en
fonction du pourcentage de recouvrement (%) moyen des mousses 42 Annexe 6. Épaisseur (cm) de la matière organique résiduelle moyenne mesurée dans les
treize sites d'étude. L'épaisseur de la matière organique résiduelle ne diffère pas significativement chez les sites suivis de la même lettre, à un seuil de
probabilité de 0,05 selon le test LSD de Fisher 43 Annexe 7. Distribution de fréquence du nombre de plantules d'épinette noire en fonction
de l'année d'établissement, à partir de l'année du feu, pour tous les sites 44 Annexe 8. Nombre de plantules à l'hectare (ha) croissant dans les différents lits de
germination dans chacun des sites. Pour un site donné, les valeurs suivies de la même lettre ne sont pas significativement différentes à un seuil de probabilité
de 0,05 selon le test LSD de Fisher 46 Annexe 9. Épaisseur de la matière organique (cm) mesurée sous le collet des plantules
dans les différents lits de germination dans chacun des sites. Pour un site donné, les valeurs suivies de la même lettre ne sont pas significativement différentes à
un seuil de 0,05 selon le test LSD de Fisher 46
Introduction
La forêt boréale nord-américaine se divise en trois zones de végétation, dont la différenciation est liée de près à l'influence des feux et du climat (Larsen 1980, Payette
1992). Du sud vers le nord, ces trois zones sont la forêt boréale fermée, la forêt boréale ouverte et la toundra forestière (Figure 1). Elles se distinguent principalement les unes des autres par la densité des peuplements, les espèces d'arbres, la fréquence des feux ainsi que le type de végétation qu'on trouve au sol (Hare 1950, Hare et Ritchie 1972, Payette et al.
1989, Payette 1992).
Figure 1. Répartition des trois zones de végétation qui composent la forêt boréale nord-américaine (Payette 1992).
Au Québec, au cours des 50 dernières années, la superficie occupée par le domaine de la pessière noire à mousses en forêt fermée a diminué de 9 % au profit de la forêt ouverte, qu'on appelle aussi la pessière à lichens (Girard et al. 2008, Girard et al. 2009). Différents facteurs peuvent être responsables d'un tel phénomène. Les perturbations en rafale, comme les feux successifs ou les épidémies de la tordeuse des bourgeons de l'épinette (Choristoneura fumiferana Clemens) suivies d'un feu, réduisent le nombre d'individus matures et la banque de graines viables disponibles pour assurer la régénération de l'épinette noire (Picea mariana (Mill.) B.S.P.) (Black et Bliss 1980, Viereck 1983, Payette et al. 2000, Girard et al. 2008). Les feux légers, qui laissent derrière eux peu de lits de
germination favorables aux graines d'épinettes noires, pourraient aussi être impliqués dans la transformation des pessières à mousses en pessières à lichens (Girard et al. 2009).
Grâce aux cônes semi-sérotineux qu'elle possède, l'épinette noire, qui domine la forêt boréale québécoise, est bien adaptée au feu, l'un des agents de perturbation les plus structurants de cette forêt (Ritchie 1987, Bonan et Shugart 1989, Johnson 1992, Payette 1992). Ces cônes sont spécialement conçus pour s'ouvrir et libérer massivement les graines qu'ils contiennent sous l'action d'une chaleur intense (Wilton 1963, Black et Bliss 1980, Viereck 1983). Or, la libération massive des graines ne peut pas assurer à elle seule le maintien de l'épinette noire dans le paysage forestier. En effet, il est crucial que le site brûlé présente des conditions favorables à la germination des graines et à la survie des plantules, sans quoi la régénération de l'épinette noire risque d'être compromise (Heinselman 1981, Zasada et al. 1983, St-Pierre et al. 1992, Jayen et al. 2006).
La sévérité d'un feu, définie comme l'épaisseur de matière organique consumée par le feu, est un facteur déterminant quant à la qualité des lits de germination disponibles après un incendie (Rowe 1983, Zasada et al. 1983, Thomas et Wein 1985 a, Johnstone et Chapin 2006a). Le maintien de conditions humides dans le lit de germination est primordial à la survie des plantules d'épinette noire car elles sont pourvues d'un court radicule (Ahlgren 1974, Fleming et Mossa 1995). Le sol minéral nu ou recouvert d'une couche de mousses ou de sphaignes, de moins de 2,5 cm d'épaisseur, conserve mieux l'humidité que la matière organique calcinée, ce qui constitue de meilleures conditions pour la germination des graines de l'épinette noire (Heinselman 1981, Zasada et al. 1983, Thomas et Wein 1985a, St-Pierre et al. 1991, Charron et Greene 2002, Greene et al. 2004, Jayen et al. 2006, Johnstone et Chapin 2006a).
Plusieurs auteurs reconnaissent que la sévérité des feux est déterminante quant au type de végétation qui s'établit après un feu (Flinn et Wein 1977, Van Wagner 1983, Viereck 1983, Wein 1983, Schimmel et Granstrôm 1996, Johnstone et Kasischke 2005). À cet effet, Schimmel et Granstrôm (1996) proposent un modèle simple qui exprime le potentiel de régénération de trois catégories de plantes en fonction de la sévérité du feu (Figure 2).
c o '•£ 01 i -•<u c -<u oo •yv cc Sévérité
Figure 2. Modèle hypothétique de la régénération de trois catégories différentes de plantes en fonction de la sévérité du feu : (A) espèces rhizomateuses; (B) espèces à banque de graines; (C) espèces qui dépendent de la dispersion des graines après feu (Modifiée de Schimmel et Granstrôm 1996).
D'après ce modèle, un feu léger, qui brûle la partie aérienne de la végétation sans consumer profondément la matière organique, avantage la régénération des espèces rhizomateuses. Lors du passage d'un feu léger, les parties souterraines de plusieurs espèces végétales demeurent intactes (Flinn et Wein 1977, Larsen 1980, Viereck 1983). Ainsi, les espèces capables de se multiplier végétativement à l'aide d'organes souterrains, comme les rhizomes, prolifèrent après le feu (Mallik et Gimingham 1985, Johnstone et Kasischke 2005, Mallik 1993). C'est le cas notamment des éricacées, dont Vaccinium spp, Rhododendron groenlandicum Oeder et Kalmia angustifolia L. (Flinn et Wein 1977, Schimmel et Granstrôm 1996). Les espèces qui dispersent leurs graines à la suite d'un feu, comme l'épinette noire, se régénèrent mal à la suite d'un feu léger puisque les lits de germination ainsi créés sont inadéquats (Wein 1983, Charron et Greene 2002, Greene et al. 2004, Johnson 1992, Johnstone et Chapin 2006a, Larsen 1980, Viereck 1983, Zasada et al. 1983). À l'inverse, un feu sévère consume la matière organique en profondeur, ce qui entraîne la création de lits de germination favorable à l'épinette noire et aux espèces qui dispersent leurs graines à la suite du feu. Lors du passage d'un feu sévère, les organes souterrains des espèces rhizomateuses sont détruits, ce qui les empêche de recoloniser le site rapidement (Flinn et Wein 1977, Larsen 1980, Heinselman 1981). Seules les espèces végétales dont les organes souterrains pénètrent le sol minéral peuvent s'établir par multiplication végétative immédiatement à la suite d'un feu sévère. C'est le cas, par
exemple, de certaines espèces de mousses telle que Polytrichum sp., qui colonisent rapidement le sol minéral mis à nu par le feu (Flinn et Wein 1977, Heinselman 1981, Viereck 1983, Thomas et al. 1994, Fleming et Mossa 1995, Johnstone et Kasischke 2005). Le feu est souvent plus sévère au pied des arbres dû à l'accumulation de combustibles fins et secs (Johnson 1992, St-Pierre et al. 1991, Miyanishi et Johnson 2002). Conséquemment, les plantules s'établissent en grand nombre au pied des arbres (Miyanishi et Johnson 2002, Jayen et al. 2006).
Avec le temps, la matière organique s'accumule sur le sol minéral (Heinselman 1981, Van Cleve et al. 1981, Wein 1983, Bonan et Shugart 1989). Ainsi, plus une forêt est âgée, plus la matière organique accumulée au sol est épaisse et plus le feu doit consumer davantage de matière organique avant d'atteindre le sol minéral (Rowe et Scotter 1973, Van Wagner 1983, Taylor et al. 1987). Le temps écoulé depuis le dernier feu (TEDF) est donc une variable à considérer dans l'étude de la sévérité des feux.
L'époque pendant laquelle un feu sévit influence la sévérité des feux (Larsen 1980, Heinselman 1981, Weber et Flannigan 1997, Kasischke et Johnstone 2005). Au Québec, en forêt boréale, le sol est couvert de neige des mois de novembre à avril, parfois même de la mi-octobre jusqu'au mois de mai. Il arrive que des feux se produisent au printemps, lorsque les feuilles, les aiguilles et les herbes de la saison précédente sont sèches. Toutefois, comme la fonte des neiges est en cours ou vient tout juste de se produire, le sol est gorgé d'eau, ce qui contraint le feu à consumer la matière organique de surface. Conséquemment, les feux de printemps sont généralement légers (Larsen 1980, Heinselman 1981, Miyanishi et Johnson 2002). En été, cependant, lorsque l'eau contenue dans le sol s'est résorbée et que la matière organique est plus sèche, le feu peut y pénétrer plus profondément.
Certaines années, les conditions climatiques sont particulièrement propices à l'incidence des feux de forêt, lors de périodes sèches et chaudes (Heinselman 1981, Viereck 1983, Johnson 1992). Dans ce cas, on s'attend à ce que les feux soient plus sévères. Au Québec, l'année 2005 fut particulièrement chaude et sèche, ce qui a engendré l'allumage et la propagation de nombreux feux couvrant de grandes surfaces (Bordeleau et al. 2006).
Bien qu'on soupçonne les feux légers d'être impliqués dans la transformation des pessières à mousses en pessières à lichens, on en sait encore très peu à ce sujet (Girard et al. 2008, Girard et al. 2009). L'objectif premier de notre étude est donc d'évaluer la sévérité des feux récents s'étant produits dans la pessière noire à mousses du Québec. Le deuxième objectif est de vérifier l'influence de la sévérité des feux sur la végétation qui colonise les sites incendiés. Enfin, le troisième objectif est d'examiner l'effet de la sévérité des feux sur la régénération de l'épinette noire.
Méthodologie
Sélection des sites
Plusieurs critères ont guidé la recherche des sites d'échantillonnage, soit l'origine du feu, la saison et l'année pendant lesquelles il a été actif, la superficie et l'accessibilité. La recherche de sites d'échantillonnage s'est d'abord effectuée à l'aide de la base de données des feux de forêt du Ministère des Ressources naturelles et de la Faune du Québec. Cette base contient notamment les coordonnées géographiques du point d'allumage de l'incendie, la superficie de la zone de feu, la cause de celui-ci (naturelle ou anthropique), ainsi que l'année et les jours pendant lesquels il a été actif. À l'aide de cartes écoforestières, nous avons vérifié que les feux se soient produits dans des pessières à mousses. Les feux sélectionnés couvrent minimalement une superficie égale à 100 hectares dans le but d'effectuer les inventaires de régénération à plus de 100 mètres des semenciers vivants (Lavoie et Sirois 1998). Comme nous étudions la sévérité des feux et que celle-ci peut être influencée par différentes conditions climatiques, nous n'avons sélectionné que des feux d'origine naturelle. Pour ce qui est de la saison, nous considérons que les feux de printemps sont initiés au mois de mai, alors que la neige vient normalement de fondre en forêt, puis s'éteignent au cours des mois de mai ou de juin. Afin d'éviter que les périodes attribuées aux feux de printemps et d'été se chevauchent, nous déterminons que les feux d'été débutent en juillet et se terminent au plus tard en août, période pendant laquelle le sol est sec. Les sites sélectionnés sont tous accessibles par la route. Or, cela signifie souvent que l'industrie forestière s'y est rendue pour récupérer le bois, ce qui a considérablement réduit 5
le nombre de sites potentiels pour l'étude. Cinq sites de feux datant du printemps 2005, 4 sites datant de l'été 2005 et 4 sites de feux de printemps d'autres années ont ainsi été retenus pour notre étude.
Dispositif expérimental
Un quadrat de 50 mètres par 50 mètres a été positionné aléatoirement dans chaque site. À l'intérieur de celui-ci, 5 bandes de 50 mètres de long par 2 mètres de large, orientées vers le nord, ont été placées à une distance de 10 mètres les unes des autres.
Variables étudiées
La saison, l'année, la sévérité du feu et la régénération de l'épinette noire sont les principales variables à l'étude. Comme la sévérité et la régénération de l'épinette noire peuvent varier en fonction du type de sol, du drainage, de l'âge de la forêt brûlée, de la densité des tiges et de l'historique des feux passés, chacune de ces variables a aussi été prise en compte.
Caractéristiques de la forêt brûlée
Trois échantillons de sol (horizon B) ont été prélevés aléatoirement dans chaque site. Des analyses granulométriques ont été effectuées sur un de ces trois échantillons, selon la méthode des hydromètres de Bouyoucos (McKeague 1978). Trois échantillons de matière organique (horizon FH) ont été récoltés dans chacun des sites, afin de déterminer le pH de la matière organique. La mesure du pH à l'eau a été effectuée sur les trois échantillons. Afin de déterminer la structure de taille et la densité des tiges à l'hectare du peuplement incendié, nous avons mesuré le diamètre à hauteur de poitrine (dhp) des tiges sur une superficie de 500 m2 comprise dans le quadrat d'échantillonnage. Une droite a ensuite été
superposée à la distribution semi-logarithmique du nombre d'épinette en fonction du diamètre afin de vérifier si les distributions de taille obtenues correspondent au modèle démographique de l'exponentielle négative, qui postule un taux de mortalité constant au fil du temps (Hett et Loucks 1976).
Les 10 plus gros arbres de la forêt incendiée se trouvant à l'intérieur du quadrat d'échantillonnage ont été abattus pour y prélever une rondelle au niveau de la souche. En laboratoire, chaque rondelle a été finement sablée pour s'assurer que les cernes de croissance soient bien visibles. Les rondelles ont ensuite été datées à l'aide du logiciel COFECHA, version 6.06, afin de déterminer l'année à laquelle chaque épinette abattue s'est implantée sur le site (Holmes 1983).
Des charbons de bois se trouvant à l'interface du sol minéral et de l'humus ont été prélevés aux extrémités de chaque bande. En laboratoire, ces charbons ont été lavés à l'eau distillée puis datés au 14C selon la méthode AMS (Accelerator Mass Spectrometry) au laboratoire
Keck Carbon Cycle AMS Facility (KCCAMS) de l'Université de Californie à Irvine. L'étalonnage des dates au radiocarbone a été fait en considérant un niveau de probabilité de 2 écarts-types à l'aide de la version 5.0.2 du programme CALIB (Stuiver et Reimer 1993) et de la base de données IntCal04.14c (Reimer et al. 2004).
Le temps écoulé depuis le dernier feu (TEDF) a été estimé à partir des trois indicateurs suivants : les cicatrices de feu observées sur les rondelles prélevées, l'âge des individus formant le peuplement incendié et la radiodatation des échantillons de charbons. Les cicatrices de feu indiquent précisément le TEDF. Chez les sites où aucune cicatrice de feu n'a été observée, l'âge des 10 individus abattus par site a été comparé. Chez les sites dont l'âge des 10 individus est très semblable, nous avons utilisé l'âge de la plus vieille épinette pour estimer le TEDF minimal. Au total, 16 échantillons de charbons ont été datés au radiocarbone, répartis inéquitablement chez 7 sites sur 13. La radiodatation des échantillons de charbon sert à estimer le TEDF chez les sites où aucune cicatrice de feu n'a été observée et où l'âge des individus est très variable ou se rapproche de la limite de longévité de l'épinette noire, soit environ 250 ans. Dans ce cas, pour un site donné, la date étalonnée la plus récente et accompagnée de la plus grande probabilité a été retenue pour estimer le TEDF maximal. Comme nous sommes en 2010 et que les dates obtenues de la radiodatation sont étalonnées en fonction du taux de carbone atmosphérique enregistré en
Sévérité du feu
La sévérité du feu a été évaluée à l'aide de deux indicateurs, soit le recouvrement des espèces végétales et l'épaisseur de la matière organique résiduelle. Le pourcentage de recouvrement de la végétation établie depuis le passage du feu a été mesuré à l'intérieur d'un quadrat de 1 m2, à tous les cinq mètres le long des bandes d'échantillonnage, ce qui
équivaut à une superficie de 50 m par site. Le recouvrement de chaque substrat, soit la matière organique noircie, les mousses, les arbustes, les sphaignes, les herbacées, les lichens, la litière, le sol minéral, le bois mort et les roches, a ainsi été évalué. Dans tous les sites, l'évaluation du pourcentage de recouvrement a été effectuée par le même observateur. À la suite de l'évaluation du pourcentage de recouvrement, l'épaisseur de la matière organique résiduelle a été mesurée sous les substrats les plus répandus du site. Par exemple, dans un site où le recouvrement des mousses et de la matière organique noircie dominent, l'épaisseur de la matière organique résiduelle a été mesurée sous ces deux substrats, à tous les 10 mètres le long des bandes d'échantillonnage, pour un total de 25 mesures d'épaisseur par substrat par site. Pour un site donné, l'épaisseur de la matière organique résiduelle a ensuite été pondérée par le pourcentage de recouvrement de chacun des substrats sous lesquels les mesures ont été prises.
Régénération
La superficie couverte par chacune des bandes, soit 100 m2, a été scrutée dans le but de
prélever toutes les plantules d'épinette noire s'y étant établies depuis le passage du feu. Sur une superficie totale de 500 m2 par site, toutes les plantules ont ainsi été identifiées et
mesurées. Ces mesures nous ont permis de calculer la densité des plantules à l'hectare, dans le but de la comparer à celle des tiges issues de graines formant le peuplement. L'épaisseur de la matière organique se trouvant sous le collet des plantules a été mesurée et le lit de germination noté. En laboratoire, l'âge des plantules a ensuite été déterminé à l'aide de deux méthodes, dans le but de connaître le patron d'établissement des plantules dans chaque site. Toutes les plantules ont d'abord été datées à l'aide des cicatrices de bourgeons. Afin de s'assurer de l'exactitude de la datation, 10 % des plantules de chaque site ont ensuite été datées à l'aide des cernes de croissance. Chez les sites de feu datant de 1999 et
2005, la datation des plantules via les cernes de croissance a confirmé les dates obtenues à l'aide des cicatrices de bourgeons. Chez le site de 1991 cependant, comme les plantules étaient plus âgées, la datation à l'aide des cernes de croissance s'est avérée plus juste que la datation via les cicatrices de bourgeons. Toutes les plantules établies à la suite du feu de
1991 ont alors été datées à l'aide des cernes de croissance. Analyses statistiques
Les analyses ont été réalisées à l'aide du logiciel SAS version 9.2 (SAS Institute Inc. 2009). Comme le plan d'échantillonnage n'est pas balancé, c'est-à-dire qu'on a un nombre inégal de feux de printemps et d'été ainsi qu'un nombre inégal de feux datant de 1991, 1999 et 2005, une correction de Kenward-Roger a été appliquée aux différents modèles statistiques élaborés.
Sévérité du feu
Recouvrement des espèces végétales
Deux analyses de la variance en procédure mixte ont été réalisées afin de déterminer quels facteurs influencent le recouvrement des espèces végétales qui colonisent le site à la suite du feu. Les analyses ont été réalisées sur la moyenne du recouvrement des substrats par bande et par site. Pour répondre aux postulats de l'ANOVA, les données ont subi une transformation racine carrée. La corrélation entre les différents substrats d'une même bande a été modélisée à l'aide d'une matrice non structurée (Littel et al. 2006, Jacqmin-Gadda et al. 2007). La première analyse tient compte de l'effet du site, du substrat et de l'interaction site*substrat, alors que la seconde considère l'effet du substrat, de la saison, de l'année et des interactions substrat * saison et substrat * année. Les résultats ont été retransformés en se basant sur le postulat de normalité (Baskerville 1972).
Comme les mousses indiquent le passage d'un feu sévère, et qu'à l'inverse les éricacées indiquent le passage d'un feu léger, une corrélation entre le pourcentage de recouvrement des mousses et des éricacées par site a été effectuée.
Épaisseur de la matière organique résiduelle
Afin de vérifier l'influence de la sévérité des feux sur le type de végétation qui colonise ensuite les sites, une analyse de la variance en procédure mixte a été effectuée sur les mesures d'épaisseur de la matière organique résiduelle prises sous les substrats les plus répandus chez les 13 sites. L'analyse réalisée considère le substrat (matière organique noircie, mousses, arbustes et lichens) en effet fixe, puis le site, la bande et la station d'échantillonnage en effet aléatoire (Littel et al. 2006, Jacqmin-Gadda et al. 2007). À noter que les mesures d'épaisseur de la matière organique ont préalablement été transformées à l'aide de la racine carrée. De plus, étant donné que les types de végétation sous lesquels les mesures d'épaisseur ont été prises diffèrent entre les sites, et donc d'une année ou d'une saison à l'autre, l'effet de la saison et de l'année n'a pu être vérifié. Les résultats ont été retransformés en se basant sur le postulat de normalité (Baskerville 1972).
L'épaisseur de la matière organique résiduelle moyenne par site a ensuite été calculée, en pondérant l'épaisseur de la matière organique moyenne par bande en fonction du pourcentage de recouvrement des substrats. Deux analyses de la variance ont ensuite été réalisées sur ces données. Pour répondre aux postulats de l'ANOVA, la transformation logarithmique a été appliquée aux données (Littel et al 2006, Jacqmin-Gadda et al. 2007). La première analyse de la variance considère le site en effet fixe dans le but de comparer l'épaisseur de la matière organique résiduelle moyenne mesurée dans les différents sites. La seconde analyse vise à distinguer l'influence de la saison et de l'année sur l'épaisseur de la matière organique résiduelle. Pour ce faire, nous avons construit un modèle mixte qui considère la saison et l'année en effet fixe, et le site en effet aléatoire. Les résultats ont été retransformés en se basant sur le postulat de normalité (Baskerville 1972).
Afin de déterminer quelles caractéristiques physiques des sites incendiés influencent l'épaisseur de la matière organique moyenne mesurée dans chacun des sites, une régression avec sélection de pas à pas a été effectuée. Les caractéristiques physiques des sites incendiés incluses dans le modèle sont le nombre de tiges à l'hectare du peuplement incendié, la pente, la texture du sol, le drainage et le TEDF. L'effet de la saison et de l'année a également été testé. Le seuil d'entrée des variables dans le modèle est de 0,20, alors que le seuil de sortie est de 0,05 (Draper et Smith 1998).
Régénération
Nombre de plantules inventoriées
Une analyse de la variance visant à comparer le nombre de plantules établies dans les sites a été effectuée. Afin de respecter les postulats de l'ANOVA, les données ont subi une transformation de rang et la corrélation entre les différents substrats d'une même bande a été modélisée à l'aide d'une matrice non structurée. Le site, le lit de germination et l'interaction entre ces deux facteurs ont été considérés en effet fixe. Une seconde analyse fut réalisée en considérant l'année et la saison en effet fixe (Littel et al. 2006, Jacqmin-Gadda et al. 2007).
Plusieurs autres variables peuvent influencer le nombre de plantules inventoriées dans un site, ce qui nous a menés à procéder à une analyse multivariée sur le nombre de plantules d'épinette noire. Une régression avec sélection de pas à pas a été effectuée, où le seuil d'entrée des variables dans le modèle est égal à 0,20 et le seuil de sortie à 0,05 (Draper et Smith 1998). Les variables indépendantes incluses dans l'analyse sont la saison et l'année du feu, le nombre de tiges à l'hectare du peuplement incendié, la pente, la texture du sol, le drainage, le TEDF et le recouvrement des différents lits de germination.
Densité des plantules
Comme les lits de germination ne sont pas également représentés dans les sites après un incendie, nous nous sommes intéressés à la densité des semis selon le type de lit de germination. L'objectif de cette analyse est de vérifier si certains substrats sont plus propices que d'autres à la germination et à la survie des plantules. Comme la base des données de la densité des plantules contient plus de 50 % de valeurs égales à zéro, nous avons construit un modèle linéaire mixte généralisé (Littel et al. 2006). La corrélation entre les substrats d'une même bande dans un site a été modélisée à l'aide d'une structure de corrélation non structurée. Les données de densité ont subi une transformation logarithmique afin d'obtenir la convergence du modèle. Comme cette analyse nécessite une variable catégorielle binaire, nous avons divisé les données en deux catégories, soit présence ou absence de plantule. Les données de la densité des plantules égales à zéro ont été placées dans la catégorie absence et les autres dans la catégorie présence. La première
analyse de la variance a été effectuée sur les données de présence, afin de connaître la probabilité de trouver au moins une plantule dans chacun des différents substrats. La seconde analyse de la variance a été réalisée afin de connaître l'effet du substrat sur la densité des semis, là où il y avait des plantules. Une matrice de corrélation de type empirique a été utilisée dans ce modèle, en considérant une matrice de départ d'indépendance, de symétrie composée puis de symétrie composée hétérogène (Liang et Zeger 1986). Les résultats ont été retransformés en se basant sur la méthode de Smearing (Duan 1983).
Epaisseur de la matière organique sous le collet des plantules
L'épaisseur de la matière organique influence la germination et la survie des plantules (St-Pierre et al. 1991, Miyanishi et Johnson 2002). L'analyse de la variance effectuée en procédure mixte vise à vérifier si l'épaisseur de la matière organique mesurée sous le collet des plantules varie en fonction du lit de germination, du site et de l'interaction entre ces deux facteurs. Pour procéder à cette analyse tout en tenant compte de l'hétérogénéité de la variance, nous avons ajouté au modèle des variances hétérogènes par site. De plus, nous considérons qu'il existe un effet général de la bande sur l'épaisseur de la matière organique mesurée sous le collet des plantules et que cet effet peut différer d'un substrat à l'autre. Conséquemment, une ordonnée à l'origine et un effet de substrat aléatoires, appliqués à la bande dans le site, ont été ajoutés au modèle (Littel et al. 2006). Comme le sol minéral est dépourvu de matière organique, il n'a pas été considéré dans cette analyse. Il en est de même pour le bois mort.
Résultats
Caractéristiques des sites étudiés
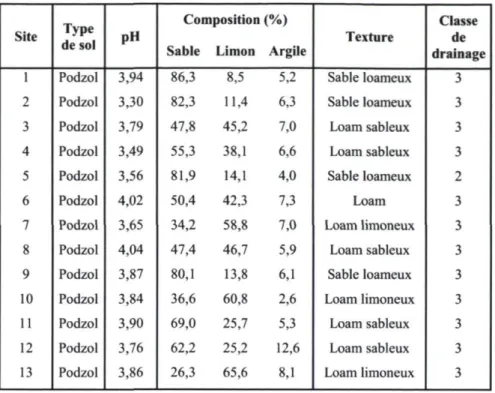
Les treize pessières à mousses incendiées incluses dans l'étude se situent entre 47 et 50 degrés de latitude Nord et entre 70 et 76 degrés de longitude Ouest (Annexe 1). La superficie des feux étudiés varie de 212 hectares à plus de 18 000 hectares (Tableau 1). L'altitude des sites d'échantillonnage passe de 290 à 745 mètres. La pente moyenne se situe
sous 10 degrés et la majorité des sites se trouve à mi-versant. Les sols de chaque site sont des podzols bien drainés (classe 2 ou 3), développés sur des dépôts dont la texture varie de loam limoneux à sable loameux (Tableau 2). Le pH de la matière organique varie de 3,3 à 4,0.
Structure de taille des peuplements incendiés
On observe deux types de distribution de taille chez les peuplements incendiés étudiés (Annexe 2). Trois peuplements (sites 5, 9 et 12) montrent une distribution de taille en forme de cloche plus ou moins complète, caractéristique d'une jeune forêt établie à la suite d'un feu. La distribution de taille des dix autres peuplements, plutôt en forme de J inversée, montre qu'il s'agit d'une forêt plus âgée, où les individus établis par graine à la suite de la perturbation d'origine sont morts ou côtoient les individus établis récemment par multiplication végétative. Le coefficient de détermination (R2), obtenu de la superposition
d'une droite à la distribution semi-logarithmique du nombre d'épinette en fonction du diamètre, varie de 0,5215 à 0,9088. À l'exception des sites 5 et 13, dont le R2 est
respectivement de 0,6428 et 0,5215, la distribution du nombre d'épinette en fonction des classes de DHP correspond bien au modèle démographique de l'exponentielle négative. Historique des feux
L'âge du plus vieil individu de chaque site varie de 77 ans à 256 ans (Tableau 3). Dans trois sites (5, 6 et 7), des cicatrices de feu ont été observées sur les rondelles puis datées. Dans ce cas, on considère que les cicatrices de feu indiquent l'année exacte du feu à l'origine du peuplement étudié et donnent une valeur juste du TEDF.
Dans tous les sites étudiés, les charbons de bois ont été prélevés au contact du sol minéral. Aucun horizon de charbon n'a été observé dans la matière organique, ce qui nous permet de croire que les treize peuplements incendiés se sont établis à la suite de feux sévères. Aucun charbon n'a été daté dans les sites 5, 9 et 12 qui présentent une structure de taille caractéristique d'une jeune forêt. Il en est de même pour les sites 8 et 11 dont l'âge des individus est d'environ 100 ans. Aucun charbon n'a également été daté dans le site 1 car son historique des feux est déjà connu grâce à une étude antérieure menée dans le Parc des
Grands Jardins de Charlevoix (Payette et Delwaide 2003). Chez les sept sites restants, un total de 16 charbons a été daté au 14C (Annexe 3).
Le TEDF chez les sites d'étude varie de 80 ans à 420 ans (Tableau 3). Tel que mentionné précédemment, le TEDF exact des sites 5, 6 et 7 a été obtenu via les cicatrices de feu. Comme l'âge de la plus vieille épinette du site 3 se rapproche de la limite de longévité de l'épinette noire et que l'âge des individus abattus y est très variable, la datation au 14C a
servi à estimer le TEDF. Chez les neuf autres sites, les individus abattus avaient sensiblement le même âge, alors le TEDF a été estimé à partir de l'âge de la plus vieille épinette.
•o 'S 3 •o C/J o 09 U 3 .S" 'pa >> C ir. O
ï
—' ■c — s U «u 3 .O" c .O" ■_> c C c -z = = = g = c c C 2 — ■_> - ^ ^ û C i s - - TZ n r3 rz CS 2 — yj t C £ û C i £ C â C t £ C e oa o C l •J > -J '-> o o ■-> ■-> o u ^ u t > > > 1 > > > > > > > > e Ou SX a. o o "-J 1 5 1 2 « ( T , « « 1 1 « « 5 1 e e ■£ o •Ii Z X O c O 5 ÙO •Ii Z C z c^ ^ ? i/j- !^ z Z a. z Z C c Ô a. ,_, u «A '«U c r i m r» o OC s C « y , o «r, T t T f r- — c «U 2, t u ■o 3 i ^ * » o i «r, r- r i r j OC t n r i r» O r i oc o *- E o T «/-, m r j Q\ o «/-> — O s o — Os 'S r^ r> « r j «r-, T t Cl t T t T t r i t i^~, c r , < _ o «>, o T t r i O s s C O ; t n r^ f~ t n oc sq s _, O X x T t «JB X T t OO oo «y-, s C oô r i p" T t '-3 g, o T «>. T t w> ■n «>, m « > i ce t n F» Ôs 5c r i t n «Vj _ X X b b b b b «-rj b f i b T t b bs T t b b "X s C K O s C T t T t " i r-', r i f^«_ 3 r f X o i ' b — X — O-' P b r i X «X -*- / " v r r , r i o i ' «A, n T t r i T t P £n (^, X r"-l 'Se O r - « r i t n b F« r^ b «bs P t n ^ t T t r j ÔO e ^** T t T t t n t n ■«t T t T t t n P X' r ^ T t r j JN o b o r» !_ ■W X X T t X b X b în o c^ r^ r» t - r- ( " ■ r - r^ X t ~ r» b r~ . 2 ' 3 != eu a. 3 a -= o o A O r r . OC r r , A oc r-n o c r ) o r i DC O s O s r j oc f » r n t n - t t n t n T t r i r i o s C s C O s s C T t s C T t O s «Os (N y ;x ■y. ■x. - y j ■x. ■J-. Efl Efl ■ r .
C e C tx C- a. CJ. c_ C C C C e E E E E E E E E E - y - o • o • z j V i o o o o o o o o o 'S c c c c c = c c c •ÛJ -S •2 ■S «x ' C ■ c ' C •n ■n 'C 'C 'C 'fi a . c c a. £ =- c C c-«V „
O s o O s «y-, «/-, 1 / - , >/~, <r, iy^, u-. u - i i r .
e o* o os O O o o o O O O O O c o- O s o O O o o c O S O S O < —— r i M r i r i r i r i r i r i f S «u r i t»l T t «v, s C r - oc o s O = r i m «X 3 SS Xi H 15
Tableau 2. Caractéristiques édaphiques des sites étudiés.
Site Type
de sol pH
Composition (%)
Sable Limon Argile Texture
Classe de drainage
1 Podzol 3,94 86,3 8,5 5,2 Sable loameux 3
2 Podzol 3,30 82,3 11,4 6,3 Sable loameux 3
3 Podzol 3,79 47,8 45,2 7,0 Loam sableux 3
4 Podzol 3,49 55,3 38,1 6,6 Loam sableux 3
5 Podzol 3,56 81,9 14,1 4,0 Sable loameux 2
6 Podzol 4,02 50,4 42,3 7,3 Loam 3
7 Podzol 3,65 34,2 58,8 7,0 Loam limoneux 3
8 Podzol 4,04 47,4 46,7 5,9 Loam sableux 3
9 Podzol 3,87 80,1 13,8 6,1 Sable loameux 3
10 Podzol 3,84 36,6 60,8 2,6 Loam limoneux 3
11 Podzol 3,90 69,0 25,7 5,3 Loam sableux 3
12 Podzol 3,76 62,2 25,2 12,6 Loam sableux 3
13 Podzol 3,86 26,3 65,6 8,1 Loam limoneux 3
■y. C o | a -e Cfl c o rt ■S o ' O ça CJ -a e o T J O TJ CJ x i CJ CJ c S cfl CJ T J Cfl CJ CJ U Cfl CJ T 3 C O TJ CJ T J CJ T ) -rt -CJ S T -Cfl CJ u-Û ri = « — 4 ,
S i
s s O o -— ^^ » O oc s o O s — oc OS i
C i r i r i a -- . T ~ O s oc m o O s — oc T tS i
~ Tt "" _ — E « u TED F maxim u (année s « O r i O r i T t o r» 1 1 E u o t 1 o O s «y-, 1 « O r i oc ce B O ■ f i s<U U es oc O oc O o «J3 £ — O T t oc =0 o «J3 a « r~ e> 0 > 1 1 r-« 1 1 o 1 « «o U . û C f l e o — o ' o — '— «U CL B P s « B "■C S o cs » f . E CJ o — cs ■o S«ÎV BL 2 « « o r i c s C s C 1 1 E CJ o 1 1 a Os i n 1 i o r i oc o «u M s '•S 9t <•< oi D «yn «n « y - , «y-. ' . r , u - , -H £- ■ -H « y ; -ri S O-r i -H O t n r~ l -ri o t n 1 1 -H O «y-, t « -H «y-, oo ( J S ,-s erne s d ce TED F minimu i (année s O r i O r i O « y , r i r o _ * * O s o o o o oc o O s o o oc O T t u B I j B W flj Cfl ■3 .2 _ ogg
s -* î J
.2 u l & l oc t n s C Os -_ — O t n T t oc r i « y , oc r-■C . . «tu c ■S II « w «V > W — « r i T t t N t ^ oo s C o T t oc c r , « y , oc r- m es ■S II « . . «tu c w «V > W — — r i ~~ -H ^ - * — 1 T t oc ' ' r-— « «U S û ec «u «V r i r r , - f yn vo r^ oc CX O r l c r , Cfl 3 CJ T -o -a cfl CJ CJ Cfl O -a c O rt T-< T 3 rt CL 3 B CJ ■ — -C o u rt X CJ u . Q t u H 17Sévérité des feux récents
Recouvrement des espèces végétales
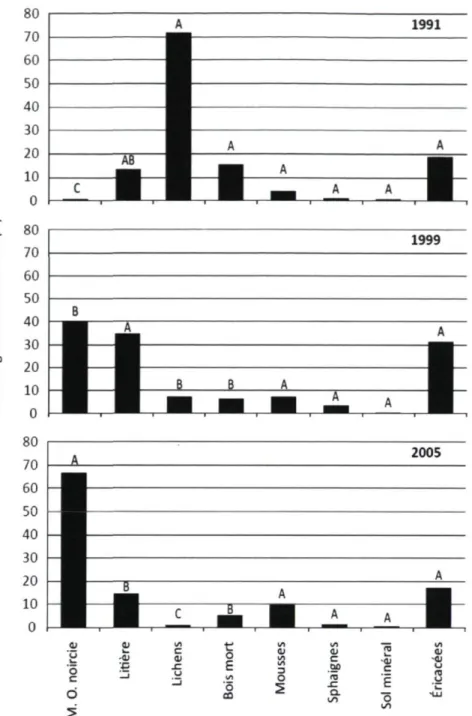
La première analyse de la variance montre que le site (p < 0,0001), le substrat (p < 0,0001) et l'interaction site*substrat (p < 0,0001) ont une influence significative sur les données de recouvrement. L'étude de l'interaction site*substrat permet de connaître le recouvrement des espèces végétales de chacun des sites (Annexe 4). La seconde analyse de variance révèle que le substrat (p = 0,0072), l'année (p = 0,0048) et l'interaction substrat*année (p = 0.0042) ont une influence significative sur le recouvrement des espèces végétales. La saison et l'interaction substrat*saison n'ont pas d'effet significatif sur le recouvrement. L'interaction substrat*année est le reflet de la colonisation graduelle de la matière organique noircie par les végétaux. Peu importe l'année du feu, les mousses, les sphaignes et le sol minéral sont très peu répandus (Figure 3). La matière organique noircie occupe une grande surface chez les sites de feux datant de 2005 et 1999. En ce qui concerne le site 1, où le feu s'est produit en 1991, ce sont plutôt les lichens qui dominent en termes de recouvrement.
La corrélation entre le pourcentage de recouvrement des mousses et des éricacées n'est pas significative (p = 0,1803) (Annexe 5). Toutefois, il est à noter que la corrélation n'a été testée que sur 13 points correspondant aux 13 sites étudiés, ce qui est peu pour ce genre d'analyse.
o c d 5 .tu '•E c «D - C tr o E yT, ! o «u c .SP «TJ - C Q . «yi •eu c o o
Figure 3. Comparaison du pourcentage de recouvrement (%) moyen de la matière organique noircie, des éricacées, de la litière, du bois mort, des mousses, des lichens, des sphaignes et du sol minéral selon l'année du feu. Pour un substrat donné, les pourcentages de recouvrement moyen suivis de la même lettre ne sont pas significativement différents, à un seuil de probabilité de 0,05 selon le test LSD de Fisher.
Relation entre la végétation et l'épaisseur de la matière organique
L'épaisseur de la matière organique résiduelle influence le type de végétation qui colonise le site après un incendie (p < 0,0001). La matière organique résiduelle est moins épaisse 19
sous les mousses (moyenne = 9,72 cm) que sous tous les autres substrats, alors que la plus grande épaisseur se trouve sous les éricacées (moyenne = 19,17 cm). L'épaisseur de la matière organique résiduelle sous les lichens et la matière organique noircie ne diffèrent pas significativement entre elles, mais elles sont significativement différentes de celles sous les mousses et les éricacées (Figure 4).
nj c c u «n /o A. «/t
g S
il
M S? m O Q. UJ 25 20 15 10 0Mousses Lichens M. O. noircie Ericacées
Figure 4. Épaisseur (cm) moyenne de la matière organique mesurée sous les mousses, les lichens, la matière organique noircie et les éricacées. Les épaisseurs mesurées sous les substrats suivis de la même lettre ne sont pas significativement différentes, à un seuil de probabilité de 0.05 selon le test LSD de Fisher.
Épaisseur de la matière organique résiduelle
Le site (p < 0,0001) a un effet significatif sur l'épaisseur de la matière organique résiduelle. L'épaisseur de la matière organique résiduelle moyenne des sites varie de 10,05 cm à 23,24 cm (Annexe 6). L'année influence aussi cette variable de façon significative (p = 0,0197). En poussant l'analyse plus loin, on constate que les comparaisons entre 1991 et 1999 (p = 0,0068) puis entre 1991 et 2005 (p = 0,0124) sont significatives alors que la comparaison entre 1999 et 2005 ne l'est pas (p = 0,4133). La saison du feu n'a pas d'effet significatif sur l'épaisseur de la matière organique résiduelle (p = 0,8650).
Selon l'analyse de sélection pas à pas, la seule caractéristique de la forêt brûlée qui influence de façon significative l'épaisseur de matière organique moyenne pondérée est la texture du sol. La matière organique résiduelle est plus mince chez les sites dont les dépôts sont sableux et plus épaisse chez ceux dont les dépôts sont loameux.
Régénération de l'épinette noire
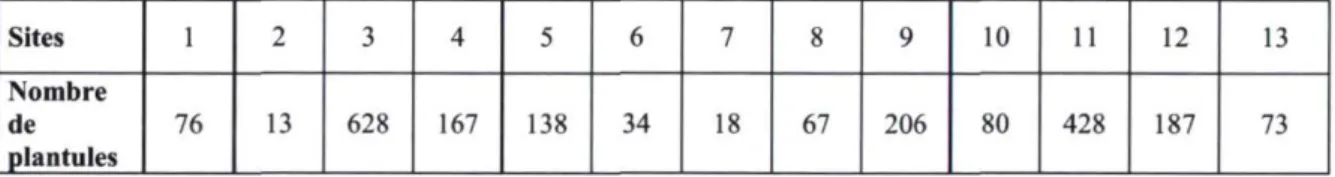
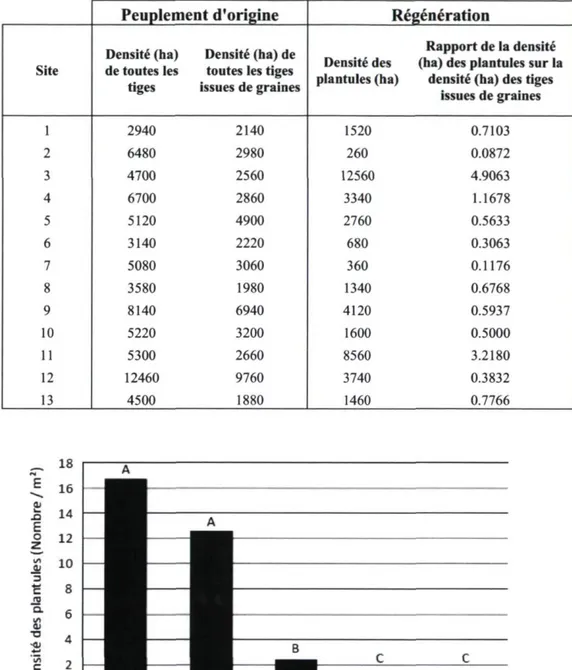
Le nombre de plantules inventoriées par site, sur une superficie constante de 500 m2, est
très variable, soit de 13 à 628 plantules (Tableau 4). En général, la densité des plantules à l'hectare est inférieure à la densité des tiges issues de graines qui forment le peuplement incendié (Tableau 5). Le rapport de la densité des plantules (ha) sur la densité des tiges (ha) de la forêt brûlée est inférieur à 80 % chez 10 sites sur 13 sites.
Le patron d'établissement des plantules à la suite du passage du feu est très semblable chez les sites de même âge après feu (Annexe 7). Au site 1 (1991), les plantules se sont établies pendant une période de 8 ans. Chez les sites 2, 3 et 4 (1999), les plantules se sont établies pendant une période de 6 ans. Quant aux feux de 2005, les plantules se sont établies pendant les quatre premières années suivant le feu, c'est-à-dire jusqu'en 2008. Dans tous les cas, la datation des plantules montre que l'établissement massif de la régénération était terminé au moment de nos inventaires à l'été 2009.
Le site (p < 0,0001), le lit de germination (p < 0,0001) et l'interaction site*lit (p < 0,0001) ont un effet significatif sur le nombre de plantules inventoriées. L'interaction entre ces deux facteurs nous permet de dresser un portrait des lits de germination dans lesquels se trouvaient les plantules inventoriées pour chaque site (Annexe 8). La matière organique fait partie des lits de germination dans lesquels on trouve le plus de plantules d'épinette dans 12 sites sur 13 sites, parfois à égalité avec les sphaignes qui sont le lit de germination de la plupart des plantules chez 5 sites. Le sol minéral et les mousses sont les lits de germination de la majorité des plantules dans seulement 2 sites sur 13. Ni la saison ni l'année du feu n'ont d'effet significatif sur le nombre de plantules inventoriées.
Selon l'analyse de sélection de pas à pas, la seule caractéristique de la forêt brûlée qui influence de façon significative le nombre de plantule est le recouvrement en sphaignes (p = 0,0002). Plus les sphaignes sont abondantes dans un site donné, plus il y a de plantules. On trouve des plantules dans 94 % des cas où l'on a de la matière organique noircie. Dans 46 % des cas où il y a des mousses au sol, on y trouve des plantules. Chez le sol minéral et les sphaignes, cette probabilité égale respectivement 35 % et 43 %. La plus faible probabilité d'occurrence de plantules s'observe dans le bois mort, avec un mince 18 % des
chances d'y observer au moins une plantule. Lorsqu'on trouve des plantules, le type de substrat a un effet significatif sur leur densité (p < 0,0001). Les sphaignes et le sol minéral sont les substrats dans lesquels on trouve la plus forte densité de plantules, suivis par les mousses (Figure 5). À l'inverse, la matière organique noircie et le bois mort sont les substrats où la densité de plantules est la plus faible.
Le site (p < 0,0001), le lit de germination (p < 0,0001) et l'interaction site*lit (p = 0,0017) ont un effet significatif sur l'épaisseur de la matière organique mesurée sous le collet des plantules. L'interaction site*lit permet de connaître l'épaisseur de la matière organique sous le collet des plantules selon le lit de germination dans chaque site (Annexe 9). Il n'existe aucune différence d'épaisseur de la matière organique sous le collet des plantules en fonction du lit de germination dans 5 sites sur 13 sites. Dans les 8 sites restants, l'épaisseur de la matière organique mesurée sous le collet des plantules varie de 1,8 cm sous les mousses au site 1 à 37,0 cm sous les sphaignes au site 4. Ni la saison ni l'année n'ont d'effet significatif sur l'épaisseur de la matière organique mesurée sous le collet des plantules.
Tableau 4. Nombre de plantules d'épinette noire inventoriées par site, sur une superficie équivalente à 500 m2. Sites 1 2 3 4 5 6 7 8 9 10 11 12 13 Nombre de 76 13 628 167 138 34 18 67 206 80 428 187 73 plantules 22
Tableau 5. Densité à l'hectare (ha) de toutes les tiges formant le peuplement incendié, des tiges issues de graines formant le peuplement et des plantules inventoriées, suivi du rapport de la densité des plantules sur la densité des tiges issues de graines.
Peuplement d'origine Régénération
Site
Densité (ha) de toutes les
tiges
Densité (ha) de toutes les tiges issues de graines
Densité des plantules (ha)
Rapport de la densité (ha) des plantules sur la
densité (ha) des tiges issues de graines 1 2940 2140 1520 0.7103 2 6480 2980 260 0.0872 3 4700 2560 12560 4.9063 4 6700 2860 3340 1.1678 5 5120 4900 2760 0.5633 6 3140 2220 680 0.3063 7 5080 3060 360 0.1176 8 3580 1980 1340 0.6768 9 8140 6940 4120 0.5937 10 5220 3200 1600 0.5000 11 5300 2660 8560 3.2180 12 12460 9760 3740 0.3832 13 4500 1880 1460 0.7766 .a E o Z a. ■a •tu «/i c 3
Sphaignes Sol minéral Mousses Bois mort M. O. noircie
Figure 5. Densité (nombre / m2) moyenne des plantules dans les différents lits de germination,
là où au moins une plantule était présente. Données retransformées à l'aide du facteur de correction de Smearing. Les densités chez les substrats suivis de la même lettre ne sont pas significativement différentes, à un seuil de probabilité de 0,05 selon le test LSD de Fisher.
Discussion
Plusieurs études soulignent l'importance de la combustion de la matière organique afin d'assurer la régénération après feu des espèces conifériennes de la forêt boréale comme le pin gris (Pinus banksiana Lamb.) et l'épinette noire (Thomas et Wein 1985b, St-Pierre et al. 1991, Duchesne et Sirois 1995, Miyanishi et Johnson 2002, Jayen etal. 2006, Johnstone et Chapin 2006a, Kemball et al. 2006). Johnstone et Chapin (2006a) montrent qu'à la suite d'un incendie la régénération se limite pratiquement aux microsites où l'épaisseur de la matière organique a été réduite à moins de 2,5 cm. L'épaisseur de la matière organique mesurée sous le collet des plantules indique que les plantules ont germé sur des microsites où la matière organique était beaucoup plus épaisse, à défaut probablement d'avoir accès à des microsites où la matière organique avait été considérablement réduite par le feu. D'ailleurs, l'épaisseur de la matière organique résiduelle moyenne minimale mesurée dans les 13 sites d'étude est de 10 cm et plus, ce qui laisse croire que les feux étudiés ont été superficiels. Pourtant, un seul horizon de charbons a été observé dans tous les sites et ce, au contact du sol minéral, ce qui indique que les feux à l'origine des peuplements étudiés étaient probablement sévères en réduisant considérablement l'épaisseur de la couche organique. Le TEDF minimal, égal à une centaine d'années ou plus dans l'ensemble des sites, révélerait alors que l'épaisse couche de matière organique mesurée dans tous les sites se serait accumulée rapidement au cours du dernier siècle. Pour le moment, cependant, aucune étude n'a montré que la matière organique s'est accumulée davantage au cours du dernier siècle que dans le passé.
Comme l'ont signalé Johnstone et Kasischke (2005), les communautés végétales qui s'établissent dans les sites légèrement brûlés sont généralement dominées par les arbustes et les graminées, alors que les mousses acrocarpes, qui colonisent rapidement le sol minéral, abondent dans les sites sévèrement brûlés (Schimmel et Granstrôm 1996). Dans les sites étudiés, le sol minéral nu et les mousses recouvrent, en moyenne, moins de 1 % de la surface. Lutz (1956) rapporte que le sol minéral occupe 35 % à 50 % de la surface affectée par un feu sévère en Alaska. Par ailleurs, la matière organique noircie et les éricacées abondent dans les sites étudiés. Les éricacées les plus communes sont Kalmia angustifolia, Rhododendron groenlandicum et Vaccinium angustifolium Aiton. La grande
abondance de ces espèces dans les sites étudiés suggère ainsi que le feu n'a consumé que la partie superficielle de la matière organique. Les données relatives à l'épaisseur de la matière organique résiduelle couverte d'éricacées abondent dans le même sens.
Les deux indices de la sévérité utilisés dans cette étude, soit l'épaisseur de la matière organique résiduelle et le recouvrement des espèces végétales, montrent que l'ensemble des feux étudiés ont été légers, sans égard à la saison pendant laquelle ils se sont produits. Pourtant, tel que démontré par Kasischke et Johnstone (2005), les feux d'été auraient dû être davantage sévères que ceux de printemps. Ici, ni les feux d'été ni les feux de printemps n'ont été sévères, affectant de manière significative la régénération de l'épinette noire. La quasi-absence de lits de germination favorables aux graines d'épinette noire dans les sites d'étude, c'est-à-dire le sol minéral nu et les minces couches de mousses et de sphaignes, compromet la régénération de l'épinette noire. On reconnaît que la matière organique carbonisée est un lit de germination défavorable à cette espèce, puisque l'humidité y est rapidement perdue par evaporation ou par drainage (Thomas et Wein 1985a, Duchesne et Sirois 1995, Charron et Greene 2002, Greene et al. 2004, Kemball et al. 2006). Or, il s'agit du substrat le plus répandu dans les sites d'étude et du lit de germination dans lequel a germé le plus grand nombre de plantules. Cela s'explique par l'abondance de la matière organique noircie et par le fait que dans les microsites humides, peu importe l'épaisseur de la matière organique, les conditions d'humidité sont favorables à la germination des graines d'épinette noire même sur les substrats organiques (Johnstone et Chapin 2006a). Toutefois, la survie des plantules qui ont germé sur la matière organique noircie risque d'être réduite. En effet, Kemball et al. (2006) ont montré que la survie des plantules établies dans les microsites légèrement brûlés diminue au fil du temps, alors que les meilleures chances de survie s'observent dans les lits de germination sévèrement brûlés. De plus, la densité des plantules calculée pour les différents lits de germination suggère que la matière organique n'est pas un lit de germination favorable à l'épinette noire. En effet, pour une surface donnée, le nombre de plantules qui ont germé dans ce substrat est nettement inférieur au nombre de plantules trouvées dans les sphaignes et le sol minéral, deux substrats peu représentés dans les sites étudiés mais reconnus pour être des lits de
germination propices à l'épinette noire (Zasada et al. 1983, Thomas et Wein 1985b, Ohlson et Zackrisson 1992, Groot et Adams 1994, Greene etal. 2004).
Au moment des inventaires de régénération, à l'été 2009, l'établissement massif de l'épinette noire était terminé dans l'ensemble des sites. Les patrons d'établissement des plantules appuient les conclusions d'autres études selon lesquelles le taux de germination de l'épinette noire est faible au cours de l'année même du feu, est maximal la première ou la deuxième année suivant le feu puis diminue considérablement par après (St-Pierre et al. 1992, Lavoie et Sirois 1998, Charron et Greene 2002, Greene et al. 2004). St-Pierre et al. (1992) suggèrent que l'absence de plantule d'épinette noire établie l'année du feu pourrait être due aux conditions difficiles qu'on trouve dans les cendres et les surfaces noircies, c'est-à-dire une chaleur intense et une forte évapotranspiration. Les plantules d'épinette noire, qui sont particulièrement sensibles à la dessiccation, ne survivraient donc pas dans ces conditions extrêmes. À ce sujet, l'analyse de sélection de pas à pas, qui a révélé que la seule caractéristique de la forêt brûlée qui influence de façon significative le nombre de plantule est le recouvrement en sphaignes, corroborent l'idée selon laquelle l'humidité constitue un facteur limitant à la régénération de l'épinette noire.
Selon Jayen et al. (2006), le nombre d'arbres semenciers est la variable la plus importante permettant de prédire le nombre de plantules nécessaire pour assurer le maintien du peuplement. Dans les peuplements de plus de 75 ans, Johnstone et Chapin (2006b) ont observé que la surface terri ère des espèces présentes avant l'incendie était corrélée à la densité de la régénération à la suite du feu. À la lumière de ces études qui suggèrent que la composition des peuplements matures devrait être la même après feu, le rapport de la densité des plantules à celle des tiges matures issues de graines devrait être égal ou supérieur à 100 %, compte tenu de la mortalité qui risque de réduire la densité des plantules au cours des années à venir. Dans 10 des 13 sites étudiés, cependant, ce rapport est inférieur à 80 %. Dans 4 de ces 10 sites, le rapport est égal ou inférieur à 50 %. Les trois sites qui affichent un rapport supérieur à 100 % se caractérisent par quelques parcelles de sphaignes densément peuplées de plantules. Chez ces trois sites, on prévoit un taux de mortalité élevé des plantules dû à leur trop forte densité dans les parcelles de sphaignes. (Annexes 4 et 8).
Comme la phase d'établissement de la régénération est terminée, les plantules d'épinette noire établies au moment de nos inventaires représentent l'ensemble des individus de la cohorte colonisatrice du prochain peuplement. Il a été démontré que la densité initiale des plantules peut induire des changements à long terme dans la composition et la structure des peuplements forestiers boréaux (Schimmel et Granstrôm 1996, Arseneault 2001, Johnstone et Chapin 2006a). Lecomte et al. (2006) ont montré que les peuplements forestiers qui tirent leur origine de feux légers demeurent ouverts tout au long de la succession écologique alors que ceux qui s'établissent à la suite de feux sévères évolue normalement vers un peuplement dense.
Les feux, les chablis, les épidémies d'insectes et la coupe forestière sont des perturbations qui influencent la succession écologique des écosystèmes boréaux comme la pessière à mousses (Heinselman 1981, Johnson 1992, Payette 1992). Grâce aux adaptations que possède l'épinette noire, la forêt brûlée devrait bien se régénérer et former un peuplement dense semblable au précédent (Heinselman 1981, Payette 1992). Lavoie et Sirois (1998) ont proposé une hypothèse pour expliquer la faible régénération de la pessière à mousses à la suite d'un incendie. Us soutiennent que lors des saisons de grands feux, comme ce fut le cas en 2005 au Québec, les conditions climatiques sont propices à la propagation de feux d'une grande intensité qui provoquent la destruction de la banque de graines viables contenues dans les cônes. La petite quantité de graines viables dispersées à la suite de l'incendie compromettrait alors la régénération de l'épinette noire. Sachant que plusieurs des feux étudiés se sont produits en 2005, cette hypothèse aurait pu expliquer la régénération déficiente observée. Toutefois, la forte densité des plantules inventoriées dans les microsites humides, comme le couvert de sphaignes ou les dépressions, laisse supposer que les graines dispersées à la suite du feu étaient nombreuses et viables. En termes de recouvrement, la dominance de la matière organique noircie qui est peu propice à la germination des graines d'épinette noire demeure l'hypothèse la plus plausible pour expliquer la faible régénération de cette espèce dans les sites d'étude.
Lorsque la pessière à mousses peine à se régénérer, il arrive qu'elle se transforme en pessière à lichens plutôt que de revenir à son état initial (Payette et al. 2000, Payette et Delwaide 2003). Au Québec, au cours des 50 dernières années, près de 9 % de la surface
occupée par la pessière à mousses s'est convertie en pessière à lichens (Girard 2008). Girard et al. (2009) ont identifié trois causes principales responsables de la transformation des pessières à mousses en pessières à lichens, soit l'incidence d'une épidémie de la tordeuse des bougeons de l'épinette (Choristoneura fumiferana Clem.) suivie par un feu, les feux successifs et le passage de feux légers. Notre étude, qui visait à décrire la sévérité des feux récents s'étant produits dans la pessière à mousses du Québec, appuie l'hypothèse selon laquelle les feux légers peuvent entraîner la transformation de la pessière à mousses en pessière à lichens. La majorité des sites étudiés semblent avoir subi le passage de feux légers, qui ont laissé derrière eux une épaisse couche de matière organique résiduelle et une surface carbonisée peu favorables à la germination des graines d'épinette noire. Tel qu'observé dans le site 1 qui date de 1991, au fil du temps, les lichens devraient recouvrir la matière organique noircie chez les autres peuplements étudiés. La régénération déficiente observée laisse entrevoir l'ouverture éventuelle des peuplements et, ultimement, la transformation des pessières à mousses étudiées en pessières à lichens.
Conclusion
La pessière noire à mousses du Québec est adaptée pour se maintenir dans le paysage pendant plusieurs siècles sous l'influence de perturbations secondaires (Bonan et Shugart 1989, Pollock et Payette 2010). Toutefois, à la suite d'une perturbation majeure comme le feu, l'épinette noire doit se régénérer suffisamment pour assurer le maintien de la forêt fermée, sans quoi le peuplement risque de s'ouvrir et de se transformer en pessière à lichens (Payette et al. 2000, Girard et al. 2009).
Dans la majorité des sites étudiés, l'épinette noire peine à se régénérer à la suite du passage d'un feu peu sévère et la densité des plantules est nettement inférieure à la densité des tiges formant le peuplement brûlé. Conséquemment, l'ouverture des peuplements est à prévoir et le passage de feux légers semble en être la cause. En effet, l'épaisse couche de matière organique résiduelle carbonisée qui résulte des feux légers est défavorable à la germination des graines et à la survie des plantules, ce qui entraîne une faible régénération de l'épinette noire. Les causes menant au passage de feux légers au cours des dernières décennies demeurent inconnues. L'augmentation des précipitations, l'accumulation accélérée de la matière organique sur le sol forestier et/ou l'intervention humaine pour éteindre les feux pourraient bien être impliqués dans le passage des feux légers mais, pour l'instant, aucune de ces hypothèses n'a été testée.
Compte tenu de la faible régénération observée après feu dans l'ensemble des sites d'étude, il serait souhaitable que le sol soit scarifié lors des opérations de récupération du bois par l'industrie forestière. La scarification, qui entraîne la création de lits de germination favorables à l'épinette noire, suivie par la plantation, pourraient pallier la faible régénération naturelle observée. On favoriserait ainsi le retour vers un peuplement fermé et le maintien de la pessière noire à mousses dans le paysage forestier du Québec.
Bibliographie
Ahlgren, CE. 1974. Effects of fires on temperate forests: North central United States. Dans Kozlowski, T.T. Fire and ecosystems. Academic Press, New York, U.S.A., 542 pages.
Arseneault, D. 2001. Impact of fire behavior on postfire forest development in a homogeneous boreal landscape. Canadian Journal of Forest Research 31:
1367-1374.
Baskerville, G.L. 1972. Use of logarithmic regression in estimation of plant biomass. Canadian Journal Forest Research 2: 49-53.
Black, R.A. et L.C. Bliss. 1980. Reproductive ecology of Picea mariana (Mill.) BSP., at tree line near Inuvik, Northwest Territories, Canada. Ecological Monographs 50: 331-354.
Bonan, G.B. et H.H. Shugart. 1989. Environmental factors and ecological processes in boreal forests. Annual Review of Ecology and Systematics 20: 1-28.
Bordeleau, C , J. Fortin, G. Gagnon, L. Innes, C. Lachance, L. Marchand, L. Morneau, D. Paré, M. Prémont et S. Simard. 2006. Insectes, maladies et feux dans les forêts québécoises en 2005. Ministère des Ressources naturelles et de la Faune, Direction de la protection des forêts, Québec, Canada, 74 pages.
Charron, I. et D.F. Greene. 2002. Post-fire seedbeds and tree establishment in the southern mixedwood boreal forest. Canadian Journal of Forest Research 32: 1607-1615. Draper, N. et H. Smith. 1998. Applied regression analysis. 3ieme édition, John Wiley and
Sons, New-York, U.S.A., 706 pages.
Duan, N. 1983. Smearing estimate: a nonparametric retransformation method. Journal of the American Statistical Association 78: 605-610.
Duchesne, S. et L. Sirois. 1995. Phase initiale de régénération après feu des populations conifériennes subarctiques. Canadian Journal of Forest Research 25: 307-318. Fleming, R.L. et D.S. Mossa. 1995. Direct seeding of black spruce in northwestern Ontario:
Temporal changes in seedbed coverage and receptivity. The Forestry Chronicle 70: 219-227.
Flinn, M.A. et R.W. Wein. 1977. Depth of underground plant organs and theoretical survival during fire. Canadian Journal of Botany 55: 2550-2554.
Girard, F., S. Payette et R. Gagnon. 2008. Rapid expansion of lichen woodlands within the closed-crown boreal forest zone over the last 50 years caused by stand disturbances in eastern Canada. Journal of Biogeography 35: 529-537.