HAL Id: hal-02818372
https://hal.inrae.fr/hal-02818372
Submitted on 6 Jun 2020
HAL is a multi-disciplinary open access
archive for the deposit and dissemination of sci-entific research documents, whether they are pub-lished or not. The documents may come from teaching and research institutions in France or abroad, or from public or private research centers.
L’archive ouverte pluridisciplinaire HAL, est destinée au dépôt et à la diffusion de documents scientifiques de niveau recherche, publiés ou non, émanant des établissements d’enseignement et de recherche français ou étrangers, des laboratoires publics ou privés.
Dynamique du sapin pectiné (Abies alba Mill.) sur le
Mont Ventoux et effet d’un gradient climatique sur le
succès reproducteur femelle
Laurent Catrice
To cite this version:
Laurent Catrice. Dynamique du sapin pectiné (Abies alba Mill.) sur le Mont Ventoux et effet d’un gradient climatique sur le succès reproducteur femelle. [Stage] Université Paris Sud - Paris 11 (UP11), Paris, FRA. 2007, 45 p. �hal-02818372�
M2 Mention Ecologie, spécialité EBE
Biologie de la Conservation - Recherche
Année 2006-2007
Soutenance les 18 et 19 juin 2007
Laurent CATRICE
Dynamique du sapin pectiné (Abies
alba. Mill.) sur le mont Ventoux et effet
d’un gradient climatique sur le succès
reproducteur femelle
Laboratoire d’accueil : INRA Centre de recherche d’Avignon
Unité de Recherche sur les Forêts Méditerranéennes
Avant propos_______________________________________________
Introduction générale________________________________________
Présentation du sujet d’étude__________________________________
1) Le mont Ventoux
a) Situation du mont Ventoux b) Historique
2) Le sapin pectiné
a) Ecologie b) Morphologie c) Reproduction
3) Morphologie des graines
a) Graines rudimentaires b) Graines plates
c) Graines vides & molles d) Graines parasitées e) Graines pleines
4) Influence de la structure du peuplement et de l’effet de l’altitude sur
la production en cônes et graines
a) Impact de la densité b) Impact du statut de l’arbre c) Effets de l’altitude
Matériels et méthodes________________________________________
I)
Jeu de données
1) Obtention des données génétiques
a) Marqueurs microsatellites chloroplastiques b) Extraction et amplification de l’ADN c) Génotypage et lecture des gels
2) Obtention des données sur les graines
a) Echantillonage
b) Radiographie des graines
c) Données phénologiques et géographiques
II)
Analyses statistiques
1) Analyses sur la diversité génétique
a) Mise en forme des données
b) Diversité et différenciation génétique c) Structuration de la diversité génétique
2) Analyses sur la production de graines
a) Structure du jeu de données b) Modèles utilisés
c) Effet maternel & environnement
d) Effet des différents facteurs sur les variables e) Optimum altitudinal
Résultats___________________________________________________
4
5
8
8 8 8 8 9 9 9 10 10 11 11 12 12 12 12 13 1314
14
14 14 14 15 15 15 16 1617
17 17 17 17 18 18 19 19 20 2021
I)
Facteurs et données utilisés dans l’analyse
II)
Diversité génétique
1) Structure génétique intra-population
2) Structure génétique inter-population
3) Comparaison des structures génétiques et géographiques
III)
Production des graines
1) Effet maternel
2) Effet des différents facteurs sur les variables
3) Effet de l’altitude & optimum
Discussion__________________________________________________
1) Dynamique de la recolonisation du mont Ventoux par le sapin pectiné
2) Dynamique actuelle et future du sapin pectiné
3) Effet maternel sur le succès reproducteur femelle
4) Effet des facteurs du milieu sur la production de graines
a) Nombre de graines
b) Taux de fécondation & pollinisation c) Dépression de consanguinité
d) Graines parasitées e) Taux de reproduction
Conclusion & perspectives____________________________________
1) Dynamique du sapin pectiné et conservation
2) Amélioration pour la poursuite de l’étude
Remerciements______________________________________________
Bibliographie_______________________________________________
Annexes____________________________________________________
21
22
22 23 2324
24 24 2628
28 28 29 30 30 31 31 32 3233
33 3335
36
39
Avant propos
L’homme a toujours eu un impact sur son environnement, mais depuis l’avènement de l’ère industrielle, celui-ci a pris des proportions telles que c’est l’ensemble de notre globe qui en est perturbé. Cette prise de conscience relativement récente a donné une nouvelle dimension à la biologie de la conservation (convention sur la diversité génétique, sommet de la Terre de Rio, 1992).
Après la conservation ex-situ, l’approche in-situ s’est considérablement développée pour tenter de répondre au défi que représente le maintien de la biodiversité à l’échelle d’une population, d’un peuplement, d’un écosystème ou d’un paysage complexe, mosaïque de ces derniers. La biologie de la conservation a notamment développé des outils de gestion faisant appels tant à la génétique des populations qu’à l’écologie, et utilise des concepts et des notions désormais fondamentaux, comme la taille efficace, la dépression de consanguinité, la capacité de reproduction, le potentiel de dispersion, le potentiel adaptatif ou la valeur sélective.
De nombreuses études sont actuellement en cours afin de mieux comprendre la dynamique forestière tant au niveau des processus d’extinction et de recolonisation que de l’évolution des successions végétales. La compréhension des facteurs écologiques et génétiques sous-jacents pourrait nous permettre d’aborder la gestion forestière d’une part dans un contexte de développement durable à court terme, et d’autre part, dans la perspective d’un changement climatique global qui semble désormais inéluctable pour une majorité de scientifiques. Dans ce contexte, le mont Ventoux fait l’objet, de par son histoire, d’importantes recherches et constitue un site d’étude intensive (ISS) du réseau d’excellence européen EvolTree.
L’INRA d’Avignon contribue largement à ces travaux, et le projet ECOGER (ECOlogie pour la Gestion des Ecosystèmes et de leurs Ressources) lancé en 2005, dans lequel s’inscrit mon stage, s’est donné pour objectif d’étudier les bases d’une gestion durable des forêts mélangées à travers l’écophysiologie, la croissance et la démogénétique des espèces constitutives de cet écosystème.
Introduction générale
Dans le cadre des changements climatiques annoncés (les modèles prévoient selon les scénarios une augmentation de 1,4 °C à 5,8 °C d’ici 2100, IPCC 2007), une des préoccupations majeures de notre société est de savoir comment vont évoluer les écosystèmes et si les espèces qui les constituent seront capables de se maintenir. Si les essences du Mont Ventoux font toutes l’objet d’un suivi, le sapin pectiné présente un intérêt supplémentaire du fait qu’il se situe à la limite sud de son aire de répartition et qu’il est donc particulièrement sensible aux modifications de son environnement. L’étude de sa dynamique générale au niveau du Mont Ventoux peut ainsi nous fournir des informations sur l’expansion ou le recul de son aire de répartition et de la vitesse du phénomène à l’échelle fine de l’individu et de la population.
En outre, nous avons choisi d’étudier le sapin pectiné le long d’un gradient altitudinal. En se basant sur l’hypothèse que la température est le facteur qui varie le plus en fonction de l’altitude (0,8 °C par 100 m), nous espérons pouvoir, par comparaison thermique, extrapoler les résultats obtenus sur un gradient latitudinal et ainsi prévoir l’évolution du sapin pectiné à une échelle géographique beaucoup plus vaste.
Bien entendu, la prédiction de l’évolution de l’aire de répartition du sapin n’a de sens que si l’on est en mesure de pouvoir agir en conséquence. Pour l’heure, le sapin pectiné est en régression sur l’ensemble de sa limite sud et on parle déjà du problème de sa conservation. A ce propos, le projet ANR vulnérabilité a été lancé en 2007 au sein de l’URFM dans le but d’étudier plus précisément le dépérissement du sapin.
La compréhension de la structure des populations et de leur histoire, et du déclin ou de l’expansion des espèces a été considérablement accrue avec l’appréhension de la structure génétique des populations. Il a été clairement démontré en biologie de la conservation que la diversité génétique était directement reliée au potentiel adaptatif d’une espèce (Frankham & al, 2002). L’observation de cette diversité génétique s’est développée en particulier par la mise au point de la technique de l’électrophorèse en biologie moléculaire, d’abord avec des marqueurs enzymatiques puis avec des marqueurs moléculaires. Le panel de marqueurs à notre disposition offre la possibilité de voir le polymorphisme de gènes ou séquences d’ADN
soumis à la sélection naturelle ou considérés comme neutre, d’origine maternelle, paternelle ou nucléaire.
Cette base génétique, combinée aux facteurs environnementaux, permet aux individus de se développer et de contribuer de manière plus ou moins importante à la pérennisation de l’espèce. Le succès reproducteur, composant de la fitness, est couramment utilisé pour évaluer la contribution d’un individu ou d’une population à la génération suivante.
Dans un premier temps, nous avons réalisé une étude de la diversité génétique des semenciers le long du gradient altitudinal afin de pouvoir comprendre leur structure spatiale, leur histoire (origines) et d’évaluer leur potentiel adaptatif. Nous avons utilisé des marqueurs microsatellites chloroplastiques dont la transmission est paternelle chez les gymnospermes et donc chez le sapin pectiné. Du fait que le sapin pectiné est dans une phase récente de recolonisation du mont Ventoux à partir de quelques semenciers « reliques », on s’attend à avoir des effets de fondation relativement marqués, avec une baisse de la diversité génétique chez les individus situés en marge de la population. Nous avons cherché également à savoir s’il existait une structuration de la population autour d’un optimum altitudinal avec une diversité moindre chez les individus situés aux marges.
Dans un second temps, nous avons étudié une composante du succès reproducteur et nous avons ainsi regardé la production en graines de chaque semencier le long du transect. On s’attend à ce que les différences observées soient expliquées principalement par la variabilité inter individus (Kärkkäinen & al., 1999 ; Mosseler & al., 2004), et dans une moindre proportion par les facteurs environnementaux, notamment la structure du peuplement. En mettant en valeur les facteurs susceptibles d’influencer la quantité et les qualités morphologiques ou génétiques des graines, on espère pouvoir proposer aux forestiers des stratégies de développement permettant de favoriser la meilleure production de graines possible chez les peuplements actuels et futurs.
Au sein de l’URFM, d’autres personnes étudient actuellement d’autres composantes de la dynamique forestière, à savoir : la variabilité spatio-temporelle de la reproduction (régime de reproduction et fertilité) (thèse en cours de Gwendal Restoux), la potentialité de la migration des semis (thèse en cours d’Annabelle Amm), etc.
L’un des objectifs à terme est de pouvoir intégrer l’ensemble des processus de la dynamique forestière : génétiques, démographiques, biologiques et les perturbations (par le feu notamment), dans un modèle individu centré, spatialisé, sur la plateforme de simulation CAPSIS.
Présentation du sujet d’étude
1) Le Mont Ventoux
a) Situation du mont Ventoux
Le mont Ventoux culmine à 1 909 m et se situe au sud ouest des Préalpes méridionales (44°10’N, 5°15’E), dans le département du Vaucluse (84) (Terre et vie). Issu de la tectogenèse pyrénéo-provençale, puis modifié par la tectogenèse alpine, il s’étend sur 24 km d’est en ouest et sur 15 km du nord au sud. Tandis que son versant sud présente une inclinaison modérée, son versant nord est lui beaucoup plus abrupt avec des pentes pouvant aller jusqu’à plus de 50 °. Le climat enregistré par la station météorologique du mont Serein (1 400 m, versant nord) est celui de l’étage montagnard médio-européen avec des précipitations annuelles de 1 270 mm et une température moyenne annuelle de 6,6 °C. L’influence méditerranéenne se ressent avec l’observation régulière d’étés subsecs.
b) Historique
Le site du mont Ventoux constitue actuellement un lieu privilégié pour étudier la dynamique forestière de différentes espèces et des successions végétales (Sagnard, 2001). Jusqu’au milieu du XIXe siècle, le mont Ventoux, tout comme la plupart des massifs forestiers français, a subi un défrichement massif au profit de l’industrie du bois et du pâturage. La reconstitution des peuplements forestiers n’a commencé qu’à la suite de la déprise rurale, sous l’impulsion du programme de Restauration des Terrains de Montagne (RTM) en 1861. A partir du milieu du XIXe siècle, des essences exotiques et naturellement présentes ont donc été implantées et on peut ainsi répertorier des peuplements de pin noir d’Autriche, pin sylvestre, pin maritime, pin à crochet, pin cembro, cèdre de l’Atlas, mélèze et épicéa pour les principaux. Aujourd’hui, ces peuplements forestiers ont atteint leur maturité et on observe une évolution plus « naturelle » avec la recolonisation, au niveau de l’étage montagnard du versant nord, des pinèdes pionnières par le sapin pectiné à partir de quelques semenciers reliques, et la reconstitution de hêtraies-sapinières de la série médio-européenne. Ces dernières sont par ailleurs classées en réserve Man And Biosphere (MAB) et sont incluses dans le programme Natura 2000.
2) Le sapin pectiné
a) Ecologie
Le sapin pectiné (Abies alba) est une espèce médio européenne sciaphile et exigeante en lumière, surtout lors de ses premiers stades de développement. On le trouve essentiellement dans l’étage montagnard de 1 100 m à 1 500 m, en peuplement pur, en association avec le hêtre à sa marge inférieure ou avec l’épicéa puis le pin à crochet à sa marge supérieure. Sur le mont Ventoux, on peut l’observer de 900 m à 1 700 m. Il est tolérant quant à la nature du sol avec cependant une légère préférence pour les sols basiques.
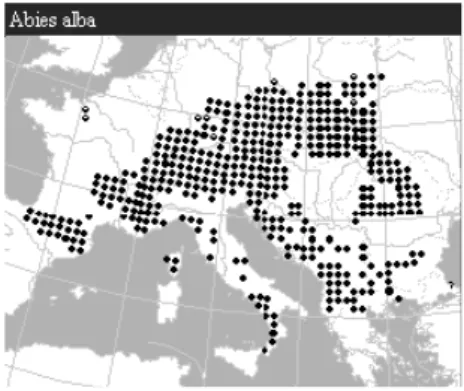
Actuellement, l’aire de répartition du sapin pectiné recouvre une grande partie de l’Europe, s’étendant de l’Allemagne jusqu’au sud de l’Italie et des Pyrénées jusqu’à l’est de la Roumanie. Cette aire est cependant en recul depuis quelques années.
Figure 3 : Aire de répartition d’Abies alba, Atlas Flora Europeae, 1998
b) Morphologie
Le sapin pectiné est un conifère de la famille des Pinaceae et du genre Abies. D’assez grande taille, cet arbre peut atteindre 40 m à 50 m de hauteur, avec une circonférence à une hauteur de 1,30 m de 260 cm à l’âge adulte. Il vit en moyenne 200 ans.
Figure 4 : Morphologie d’Abies alba
La structure du houppier est pyramidale puis devient tabulaire chez les individus âgés. Les branches sont disposées horizontalement en verticille sur un tronc droit présentant une écorce grisâtre pour les jeunes individus et gris foncé à brun rouge par la suite. Les aiguilles sont pluriannuelles et insérées de manière pectinée (en peigne, d’où son nom) directement sur le rameau et présentent deux bandes blanchâtres de stomates sur la face inférieure.
c) Reproduction
Le sapin pectiné est une espèce monoïque, c'est-à-dire que les inflorescences femelles et mâles sont présentes simultanément sur un même individu. Le sapin est une espèce anémophile (dispersion du pollen par le vent). L’allofécondation est majoritaire (94,6 % Schroeder & al (1994) et 85,4 % au niveau du mont Ventoux Fady & al (1999)), malgré
l’absence de système d’auto- incompatibilité. Figure 5 : Reproduction des conifères, encyclopédie
Encarta
Les individus matures (à partir de 30 ans environ) vont développer sur les rameaux de l’année n-1 des chatons mâles au niveau des branches basses, de couleur jaune puis brun/rouge à maturité, et des cônes femelles sur les branches les plus hautes. La dispersion du pollen s’effectue de mi-avril jusqu’à fin mai mais les fleurs femelles sont réceptives durant seulement quelques jours (maximum une semaine) à la même période. La pollinisation, la fécondation et la maturation des cônes se font dans la même année et les graines se dispersent après la désarticulation du cône, de septembre à janvier. Le mode de dispersion des graines est anémochore (par le vent), et les graines sont d’ailleurs pourvues d’une aile.
3) Morphologie des graines
a) graines rudimentaires
Les cônes femelles sont de forme allongée et sont composés d’écailles disposées en spirales autour d’un axe rigide appelé rachis. A la base de chacune des écailles se situent deux graines munies d’une aile. Il a été démontré sur de nombreuses espèces de conifères, dont le sapin pectiné, que les graines des écailles situées au niveau des régions distales étaient non fertiles du fait des contraintes physiques liées à leur position au sein du cône (Arista & al., 1996 ; Owens & al., 1998, 1994). Il est fort probable que ces écailles constituent un investissement de structure et donc une perte pour la reproduction. Ces graines peuvent se distinguer des autres par leur taille réduite et leurs ailes atrophiées. Par la suite, ce type de graine sera qualifié de rudimentaire.
b) graines plates
Les autres graines, situées donc dans la partie centrale du cône, présentent toutes les structures nécessaires à la reproduction. A maturité, les écailles des cônes vont s’écarter afin de permettre la fécondation par le pollen durant quelques jours seulement (Owens & al., 1994). A ce stade, on peut d’ores et déjà constater qu’une partie des graines potentielles a avorté, principalement à cause du gel (Owens & al., 1998). La pollinisation représente quant à elle une étape difficile chez le sapin pectiné et beaucoup d’autres gymnospermes : le mode de pollinisation anémophile donne une grande importance aux conditions météorologiques (Krouchi & al., 2004) et à la structure du peuplement (dense ou pas) durant les quelques jours de réceptivité des cônes femelles. Il a été constaté que des pluies, ou simplement du brouillard, pendant cette période suffisaient pour diminuer de manière significative le taux de fécondation (Mosseler & al., 2000). Le manque de pollen, ou la pollinisation par un pollen non viable, entraîne le non développement de la graine ou un avortement précoce de l’embryon et la graine reste aplatie, uniquement entourée par un tégument rigide (Owens & al., 1994, 1990 ; Kärkkäinen & al., 1999). Ces graines seront qualifiées de plates.
c) graines vides & molles
Les graines ayant reçu un pollen viable, et qui ont donc été fécondées vont commencer à développer les structures nécessaires à la graine (cotylédons, embryon, etc.). Cependant, une partie d’entre elles vont avorter à cause des gelées tardives entre autres (Mosseler & al., 2000). Le sapin pectiné, ne dispose pas de gènes d’auto-incompatibilité et peut dès lors s’autoféconder. La dépression de consanguinité qui peut en résulter par une augmentation de l’homozygotie, notamment celle des gènes létaux ou délétères, provoque l’avortement d’une partie des graines (Owens & al., 1994, 1990 ; Mosseler & al., 2000 ; Willson & al., 1984). Celles-ci ont une forme ronde normale, mais sont vides (Kärkkäinen & al., 1999). Certaines dont l’avortement a été plus tardif vont être vides avec un résidu de structures nourricières et d’embryon en leurs centres (Owens & al., 1998, 1990). Les premières seront dénommées graines vides et les secondes graines molles. Mosseler (2000) a montré que des graines non pollinisées pouvaient elles aussi développer un pseudo embryon qui avorte par la suite, ce qui pourrait apporter une confusion entre les graines plates et les graines vides. L’estimation de leur proportion étant faible parmi les graines vides, je maintiendrai la classification des graines présentée jusqu’alors.
d) graines parasitées
Les graines du sapin pectiné subissent des pertes dues au parasitisme. Il en existe de nombreux, mais le plus important observé sur le site du mont Ventoux est Megastigmus
suspectus Borr. (Hymenoptera, Torymidae). L’insecte s’introduit au moment de la réceptivité
des cônes femelles et durant le développement de l’embryon (Owens & al., 1998). La larve pondue au sein de la graine va se nourrir des structures internes de celle-ci pour se développer. La cavité de la graine ne contient dès lors que la larve.
e) graines pleines
Les graines qui ont pu être fécondées et développer un embryon avec toutes les structures nécessaires sont donc considérées comme viables. Elles seront qualifiées de pleines par la suite. La viabilité des graines dépend donc de l’ensemble des facteurs régissant la pollinisation, le parasitisme et le bon développement des graines.
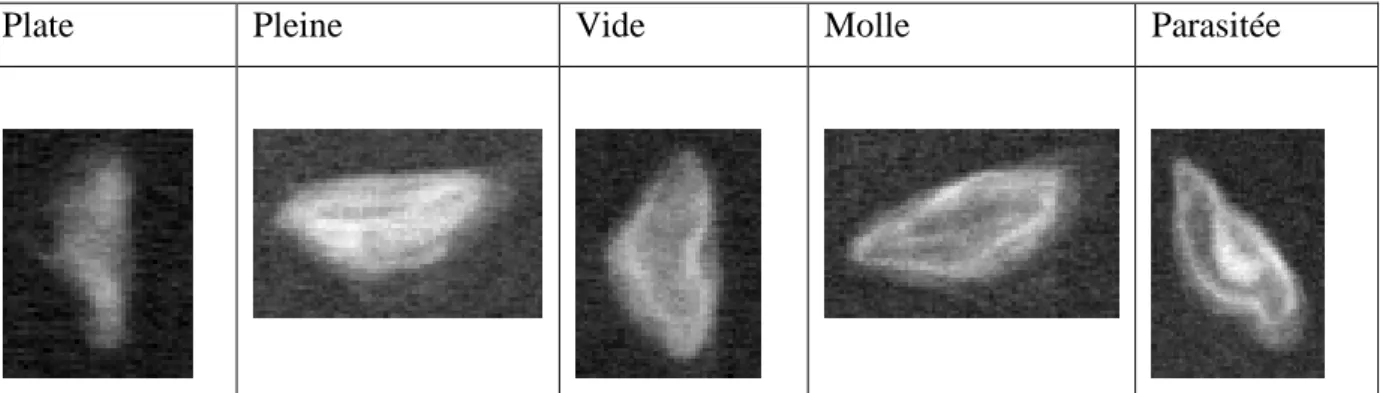
Plate Pleine Vide Molle Parasitée
Tableau 1 : Distinction des différents types de graines à la radiographie.
4) Influence de la structure du peuplement et de l’altitude sur la production en
cônes et en graines
a) Impact de la densité
La densité d’un peuplement est importante car elle joue sur plusieurs facteurs essentiels que sont l’accès à la lumière (Difazio & al., 1998), l’importance et la composition du nuage pollinique et l’impact du parasitisme.
A forte densité, les individus reproducteurs sont en forte concurrence pour l’accès à la lumière qui est un facteur très limitant pour l’acquisition de ressources. La conséquence est que les cônes femelles sont produits en moindre quantité mais disposent en revanche de
ressources plus importantes. Les graines seront plus nombreuses au sein de chaque cône et subiront moins de pertes en proportion du nombre total de graines (Arista & al., 1996). Le nuage pollinique sera d’autant plus diversifié qu’il y aura d’arbres reproducteurs dans le peuplement et les risques d’autofécondation d’autant plus faibles (Arista & al., 1996). Le parasitisme est important (rapport Invagraine).
A faible densité, les arbres reproducteurs ne sont pas limités par la ressource en lumière et de nombreux cônes sont produits jusqu’à des branches relativement basses. Le nombre de graines est cependant moins important au sein de chacun des cônes. L’étape la plus difficile pour la reproduction est la pollinisation. Le manque de contribution en allo-pollen (issu des autres individus) crée un nombre important de graines non fécondées et augmente le taux des graines plates (Arista & al., 1996). De plus, l’autofécondation plus importante va augmenter par conséquence l’expression de gènes délétères et de fait le taux des graines vides et molles.
Le parasitisme semble avoir un impact plus modéré sur les arbres isolés ou à faible densité, même s’il reste important. C’est à moyenne densité que le parasitisme est le plus faible (rapport Invagraine).
La prédation est plus importante (Difazio & al., 1998) mais ne sera pas considérée dans cette étude.
b) Impact du statut de l’arbre
Le statut de l’arbre a une influence également sur la disponibilité en lumière. On comprend aisément que les individus dominants auront un accès privilégié et produiront des cônes femelles avec un taux de graines viables supérieur aux arbres dominés. La contrepartie est que les arbres dominants, plus hauts vont être plus parasités que les autres. Le facteur visuel semble décisif en l’occurrence.
c) Effets de l’altitude
L’altitude a été peu prise en compte dans les différentes études qui ont été réalisées sur le sapin pectiné, d’où l’intérêt de ce stage ! Il a juste pu être démontré qu’il y avait un décalage dans la maturation des cônes avec un retard chez les individus situés à une altitude plus élevée (Owens & al., 1998), sans doute afin d’éviter les gels plus tardifs.
Matériels & Méthodes
Le gradient altitudinal est compris entre 970 m et 1 524 m sur le versant nord et il a été conçu de manière à ce que l’exposition soit la plus constante possible en tout point du gradient. Afin de respecter cette condition et du fait de la géographie du mont Ventoux, le transect est constitué de deux parties. Le transect bas est situé de 970 m à 1 139 m et le transect haut de 1 241m à 1 524m. Soixante cinq sapins adultes y sont suivis depuis 2006, dont cinquante trois étaient reproducteurs en 2006.
I)
Jeu de données
1) Obtention des données génétiques
a) Marqueurs microsatellites chloroplastiques
Les microsatellites sont des petites séquences d’ADN composées d’une répétition d’un motif de 1 à 6 nucléotides. On retrouve ces séquences dans l’ensemble du génome (nucléaire, mitochondrial et chloroplastique), et de manière relativement fréquente (104 à 105 régions microsatellites par génome). Leur polymorphisme est assez important et se défini par le nombre de répétition qui entraîne des longueurs différentes pour la séquence d’ADN, facilement mises en évidence par la technique de l’électrophorèse.
Le génome chloroplastique a la particularité d’être transmis de manière uniparentale, en l’occurrence par le père chez le sapin pectiné. Ce génome ne recombine pas et on observe ainsi l’existence de lignées chloroplastiques permettant de réaliser un suivi sur l’histoire d’une population. L’autre avantage est que ce génome est très conservé d’une espèce à l’autre et on peut ainsi profiter des recherches déjà réalisées sur d’autres espèces proches.
Nous avons utilisés pour cette étude 2 amorces (PT71936 et PT63718) découvertes par Vandramin en 1996 sur Pinus thunbergii, et 2 amorces (PT30141 et PT30249) décrites par Liepelt en 2001, permettant de révéler 4 marqueurs microsatellites, tous mononucléotidiques.
b) Extraction et amplification de l’ADN
A la mi-mars, des rameaux verts ont été prélevés sur les 63 individus du transect et conservés en chambre froide à - 4°C. Pour chacun, un poids de 50 mg d’aiguille a été placé
dans une plaque d’extraction de 8*12 tubes avec une bille en métal. La plaque a été placée dans un bain d’azote liquide puis dans le broyeur à bille Quiagen afin de réduire en poudre les aiguilles. L’extraction de l’ADN se fait ensuite avec le DNeasy 96 Plant Kit Quiagen (protocole en annexe 1). La quantité d’ADN a ensuite été ramenée à une concentration de 50 ng.µ L-1 pour l’amplification.
L’ADN a été amplifié par PCR (Polymérase Chain Reaction) sur un thermocycleur PTC 100TM (Programmable Thermal Controller) de MJ Research. Cette étape a été effectuée avec 4 paires d’amorces révélant en tout 4 loci polymorphes. Deux de ces amorces (PT71936 et PT30249) sont marquées par des fluorochromes détectés à 700 nm (IRDye700) et les deux autres (PT30141 et PT63718) par des fluorochromes détectés à 800 nm (IRDye800). Les deux amorces révélées par le même fluorochrome sont associées à des fragments d’ADN de taille différente. Ceci nous permet de pouvoir lire les 4 marqueurs sur un même gel et de ne faire qu’une migration par échantillon, d’autant que ces 4 marqueurs sont multiplexés.
Le succès de l’opération a été vérifié en faisant migrer les échantillons sur un gel d’agarose à 1 % à 100 V avec une gamme étalon de concentration allant de 20 à 100 ng.µ L-1.
c) Génotypage et lecture des gels
Le génotypage se réalise avec un séquenceur automatique Li-Cor Long Readir 4200 permettant de lire à 700 nm et 800 nm. La migration des échantillons s’est faîte avec des marqueurs de taille et un individu référence de manière à faciliter la lecture des génotypes. L’analyse des gels a été effectuée avec le logiciel SAGA GT. Le marqueur PT63718 n’a pas donné des résultats satisfaisants, et je n’ai donc conservé que les marqueurs PT30141, PT71936 et PT30249
Ce travail réalisé pour les trois marqueurs, les données ont finalement été regroupées afin de disposer de l’haplotype de chaque individu.
2) Obtention des données sur les graines
a) Echantillonnage
Afin de pouvoir évaluer le succès reproducteur, les cônes de 53 reproducteurs sur les 65 arbres du transect Nord Ventoux ont été prélevés en automne 2006. Après avoir été mesurés, ces derniers ont été décortiqués et l’ensemble de leurs graines ont été dénombrées, pesées et conservées au sein de la graineterie de l’URFM à une température de 5°C.
chacun des arbres. Cependant, pour des raisons de temps, seuls 3 cônes par arbre ont été analysés. Les cônes ont été choisis en fonction de leur taille : un petit, un moyen et un grand. Ceci nous permet d’avoir un échantillon représentatif des cônes de chaque arbre et d’éviter d’avoir un biais lié à la taille du cône dans nos résultats.
Pour l’estimation du nombre de chaque type de graine décrit plus haut, la plupart des études ont recours à la dissection systématique des graines. Nous avons préféré utiliser la radiographie aux rayons X des différents lots de graines pour les discriminer. Cette technique présente des résultats tout à fait satisfaisants.
b) Radiographie des graines
La radiographie se fait avec un Faxitron X-ray Mx-20 et un scanner EZ240. Les graines composant le cône sont disposées sur une plaque puis exposés aux rayon pendant 1 min 44 s, à une intensité de 20 kV. Ces deux paramètres ont été établis d’une part pour conserver la viabilité potentielle des graines et d’autre part de manière à obtenir une qualité de photo suffisante pour permettre de déterminer chaque type de graine. L’enregistrement au scanner s’est fait avec un balayage de 70 ms à une résolution de 2 pb.
Le principe de la radiographie nous permet d’acquérir une image de l’intérieur des graines avec un contraste entre les parties vides et la matière (annexe 2). A partir de la littérature, nous avons choisi de distinguer 5 types de graines : graines plates, graines pleines, graines vides, graines « molles » et graines parasitées. Le décompte de chaque type de graine s’est effectué sur un tirage papier avec pointage systématique de chaque graine. Il a été vérifié que l’image n’était pas biaisée en fonction de la position des graines sur la radiographie.
c) Données phénologiques et géographiques
Les données concernant la phénologie des individus a été obtenues par les diverses équipes de l’URFM et sont disponibles sur une base de données interne à l’INRA. J’ai ainsi pu obtenir des informations concernant : la longueur (lon.co), le poids (pds) et le nombre de graines (grai) de chaque cône, l’altitude (alt) et la position GPS, la circonférence (cir), la hauteur (hau), le statut social (dominant, codominant ou dominé) (sta), la production en chaton mâle 2006 (pm), le nombre de cône femelle produit (ncf), et une estimation de la densité locale en terme de surface terrière pour chaque individu (dens).
II)
Analyses statistiques
1) Analyses sur la diversité génétique
a) Mise en forme des données
J’ai analysé la diversité génétique sur l’ensemble du gradient. Cependant, afin de pouvoir étudier cette diversité au niveau des marges du transect, j’ai d’abord divisé le transect en 7 sous-populations relativement homogènes en nombre d’individu, réparties le long du gradient altitudinal, la population « a » étant la plus basse et la population « g » la plus haute. La division du transect en deux parties, transect bas (sous-populations a, b et c) et transect haut (sous-populations d, e, f et g), a également justifié ce type d’approche afin de voir si la diversité génétique confirme ou infirme le fait qu’il s’agisse d’une seule et même population.
b) Diversité et différenciation génétique
L’analyse de mes données génétiques a été réalisé avec le logiciel GenAlEx6, créé par Peakall & Smouse (1996, 2006), et qui fonctionne en tant que macro sous Excel.
J’ai calculé les indices de diversité de chaque sous populations avec l’indice de Nei :
∑
−
=1 pi²
He pour i allèles.
La différenciation génétique, peut elle se calculer d’après 2 méthodes : la méthode Infinite Allele Model (IAM) considère que deux haplotypes différents sont issus d’un seul événement mutationnel, et la méthode Stepwise Mutation Model (SMM) considère qu’il y a un pas mutationnel pour chaque différence d’une répétition (pour les microsatellites) entre deux haplotypes. J’ai donc comparé la différenciation génétique des 2 modèles, le Rst (IAM) et le
Gst (SMM), avec le logiciel PermutCpSSR. N’ayant pas trouvé de différence significative,
j’ai utilisé la distance génétique de Nei basée sur la méthode IAM.
c) structuration de la diversité génétique
J’ai utilisé une Analyse de Variance Moléculaire (AMOVA) qui permet d’analyser l’origine intra- ou inter-population des différences génétiques observées entre les sous-populations. J’ai étudié d’une part s’il y avait une structuration au sein des 7 sous-sous-populations. Puis, j’ai regardé séparément les différences entre la marge inférieure et la marge supérieure de la population en créant une sous-population correspondant à la marge étudiée (inférieure ou supérieure), que j’ai ensuite comparée au reste du peuplement.
J’ai enfin cherché à savoir si la diversité génétique observée pouvait être liée à la structuration géographique des populations le long du transect. Pour ce faire, j’ai calculé les distances génétiques de Nei et les distances géographiques entre chaque individu ou population, et j’ai regardé leur corrélation avec un test de Mantel (cf. GenAlEx6 appendix 1). Cette approche a été complétée par une analyse de la structure spatiale des distances génétiques de Nei.
2) Analyse sur la production de graines
Les données ont été traitées sous « R » (http://www.r-project.org/) qui est un logiciel gratuit sous le terme de la « Free Software Foundation’s GNU General Public License ».
a) Structuration du jeu de données
Le jeu de données est relativement important puisqu’il comporte 6 catégories de graines et 3 variables liées à chaque cône, 10 variables propres à chaque arbre, et le tout pour 53 individus.
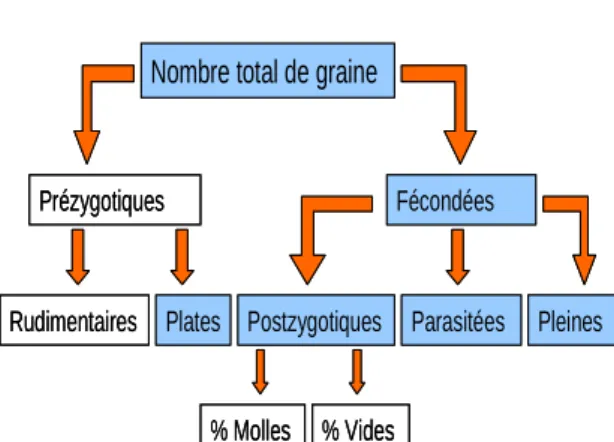
Dans un premier temps, les données sur les graines ont été réarrangées de manière à fournir une information pertinente sur la production de graines et deux nouvelles catégories de graines ont ainsi été définies :
- Graines fécondées, qui incluent toutes les graines ayant pu être fécondées. Cela fournit une estimation de la fécondité.
- Graines postzygotiques, qui regroupent les graines qui ont avorté après la fécondation. Dans l’hypothèse où la cause principale réside dans l’expression de gènes délétères, cela donne une information sur la dépression de consanguinité.
Rudimentaires
Nombre total de graine
Prézygotiques Fécondées
Plates Postzygotiques Parasitées Pleines
% Molles % Vides Rudimentaires
Nombre total de graine
Prézygotiques Fécondées
Plates Postzygotiques Parasitées Pleines
% Molles % Vides
Figure 6 : Répartition des types de graines
J’ai défini au final 6 variables pour décrire la production de graines par cône :
taux de graines plates (plates / totale), taux de graines fécondées (fécondées / totale), taux de pertes postzygotiques
(post-zygotiques / fécondées), taux de graines
pleines (pleines / fécondées), taux de parasitisme (parasitées / fécondées), et nombre de graines produites.
Dans un second temps, une analyse des données par ACP (Analyse en Composantes Principales) en tenant compte des coefficients de corrélation (annexe 3), a permis de mieux cibler les variables explicatives choisies comme facteur. Certains facteurs, apportant une information redondante ou une information non exploitable pour expliquer la variabilité des graines par cône, n’ont pas été pris en compte.
b) Modèles utilisés
Pour l’analyse statistique, j’ai cherché à savoir quels étaient les liens éventuels entre les différentes variables à expliquer (nombre de graines par cône, taux de graines plates, taux de graines fécondées, taux de graines pleines, taux de graines parasitées et taux des pertes postzygotiques) et les facteurs. J’ai utilisé pour cela des modèles linéaires.
Dans le cas du nombre de graine, un modèle linéaire généralisé (GLM) a été utilisé avec la fonction de transformation log (poisson). En revanche dans le cas des variables représentant un taux, j’ai dû utiliser un GLM avec la fonction de transformation Logit. En fonction de la structuration des données entre Y et X, la relation établie a été linéaire ou polynomiale.
c) Effet maternel & environnement
La production des graines est déterminée en premier lieu par l’effet génétique de l’individu (effet mère), puis par son environnement et l’association des deux. Afin de distinguer ces différents effets, j’ai tout d’abord effectué une analyse de variance (Anova) de chaque variable en fonction des individus. La part de variance expliquée par le modèle est celle qui est imputable à l’effet génétique des mères et aux différences inter individuelles dues aux facteurs environnementaux. La part résiduelle représente la variabilité intra individuelle (liée aux différences entre les 3 cônes). J’ai ensuite regardé la variabilité expliquée par les différences inter individuelles en ayant enlever auparavant toute la variabilité expliquée par les facteurs environnementaux. J’obtiens ainsi la variabilité inter individuelle qui n’est pas expliquée par les facteurs environnementaux et qui est donc due aux différences génétiques des individus (effet mère). Il est à noter que j’inclue dans les facteurs environnementaux des paramètres propres à l’arbre, tels que la production en gamètes, la taille du tronc, etc. ; cela est discutable mais j’ai considéré qu’ils étaient en parti régis par les conditions environnementales pour lesquelles peu d’informations étaient disponibles (paramètres physico-chimique du sol, T°C journalière, Humidité de l’air…).
d) Effet des différents facteurs sur les variables
Les facteurs qui ont été mesurés sont propres à chaque individu et sont donc masqués par l’effet mère. Seuls les facteurs mesurés au niveau des cônes peuvent apporter une variabilité qui n’est pas expliquée par l’effet mère. Pour analyser les différents facteurs environnementaux, je n’ai pas pris en compte l’effet mère et j’ai ensuite observé les effets propres de chaque facteur conditionnellement aux autres.
Une anova m’a permis de voir si il y avait un effet significatif des facteurs étudiés et j’ai regardé la significativité de la part de déviance expliquée par le modèle par rapport à la déviance totale avec un test de Khi². Dans le cas où il y avait un effet significatif de X sur Y, j’ai regardé la valeur du coefficient, ainsi que sa significativité avec un test basé par rapport à la loi normale centrée réduite.
Pour finir, j’ai fait une procédure « stepwise » en me basant sur les critères de sélection de modèles, afin de savoir quelle combinaison de facteur explique le mieux chacune des variables.
e) Optimum altitudinal
Une des hypothèses de départ sur l’effet de l’altitude est qu’il existe un optimum écologique à une altitude particulière correspondant aux conditions les plus favorables au développement du sapin pectiné. Au delà et en deçà de cet optimum, le sapin se retrouverait moins performant et notamment au niveau de son succès reproducteur. Dans un premier temps, j’ai cherché à modéliser, lorsque les données s’y prêtaient, la variable qui nous intéresse avec l’altitude en utilisant une fonction présentant un maximum ou un minimum altitudinal afin de pouvoir le définir :
(
altitude optimum)
const tcoefficien x
f( )= ×( − puissance)+
Dans un second temps, j’ai cherché les paramètres (notamment l’optimum) de cette fonction qui minimisent la somme des carrés de la différence entre la valeur réelle de la variable et celle estimée par la fonction. Sous « R », cela est réalisable avec la procédure optim (fonction, paramètres). J’ai ainsi pu obtenir l’altitude minimale ou maximale pour laquelle il y avait un changement de comportement de la variable.
Résultats
I)
Facteurs et données utilisés dans l’analyse
1 grai pds pds.g alt ncf pm cir hau vol dens -1 -1 1 grai pds pds.g alt ncf pm cir hau vol dens -1 -1 1 grai pds pds.g alt ncf pm cir hau vol dens -1 -1 1
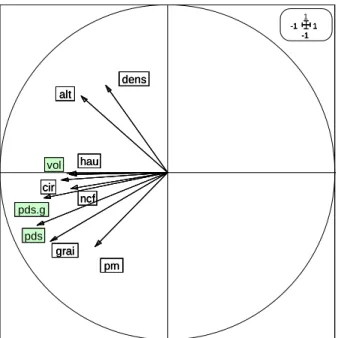
Figure 7 : ACP sur les facteurs disponibles
Le poids pour 1000 graines (pds.g) et le poids des cônes (pds) apportent une information non exploitable pour expliquer la variabilité des graines car leur poids dépend lui-même de la qualité des graines et on ne connaît pas le poids spécifique de chaque type de graine. La variable volume (vol) n’a pas été retenue du fait qu’elle soit très corrélée avec la circonférence et la hauteur, et qu’elle explique moins de processus biologiques.
Les variables et facteurs retenus sont résumés dans le tableau 2
53 individus 3 cônes / individu
8 facteurs
altitude, densité, production chatons mâles, nombre cônes femelles, hauteur, circonférence, statut
Nombre de graines par cône, taux de graines fécondées
5
variables Nombre de graine par cône
% de graines fécondées, % de graines plates, % des pertes postzygotiques des graines, % de graines parasitées, % de graines pleines
Tableau 2 : résumé des variables et des facteurs utilisés dans l’étude.
Les facteurs qui ont été retenus pour l’analyse, sont tout de même assez corrélés entre eux (figure 8), et il reste un risque de confusion entre les effets dus à la hauteur, la circonférence et le nombre de cônes femelles, et entre la production en chatons mâles et le nombre de graines par cône. La densité est corrélée avec l’altitude, mais cela est trompeur car en réalité il y a une forte différence entre les 2 parties du transect : le bas est peu dense et le haut beaucoup plus (figure 9). Au sein de ces 2 parties, la densité n’est pas corrélée avec l’altitude.
grai pré pré.p fec fec.p post post.p plei plei.p para para.p mol mol.p vid vid.p pla pla.p rud rud.p ncf pm cir hau dens -1 1 -1 1 grai pré pré.p fec fec.p post post.p plei plei.p para para.p mol mol.p vid vid.p pla pla.p rud rud.p ncf pm cir hau dens -1 1 -1 1
Figure 8: ACP des facteurs retenus avec les variables Figure 9 : structure de la densité avec l’altitude.
L’individu TB118 a été retiré du jeu de données car ses cônes avaient été entièrement rongés par des chenilles de Diorictrya abietella et n’apportaient aucune information sur la proportion des différents types de graines. Cette initiative a été justifiée durant les analyses statistiques, les distances de Cook et les résidus normalisés montrant que TB118 tirait les données dans son sens de façon trop importante.
II)
Diversité génétique
1) Structure génétique intra-population
Les 3 marqueurs que j’ai utilisés se sont tous avérés très polymorphes, et on trouve 31 haplotypes différents dont 22 propres à un seul individu, pour les 53 au total (tableau 3).
Tableau 3 : Polymorphisme des marqueurs
Dans les 7 sous-populations, la diversité génétique est relativement homogène avec un He
moyen de 0,827 et une variance de 0,046. Il y a cependant une légère différenciation pour la population la plus haute (g) qui a une diversité plus faible que les autres (He = 0,735).
Marqueurs Nombre d’allèles Allèles uniques pour un individu PT71936 14 4 PT30241 8 2 PT30249 5 1 1000 1100 1200 1300 1400 2 0 3 0 4 0 5 0 6 0 alt d e n s
2) Structure génétique inter-population
L’AMOVA nous montre que seulement 2 % (p = 0,079) de la variabilité est expliquée
par la différenciation en 7 sous populations. La diversité génétique entre le transect bas et le transect haut est similaire (He = 0,946 et He = 0,922) et l’AMOVA révèle que toute la
variabilité est alors expliquée par les différences intra-population (p = 0,276).
La marge inférieure (7 individus de 970 m à 1 025 m) n’est pas différente du reste du peuplement et l’AMOVA permet de montrer que toute la variabilité est expliquée par les différences intra-population (p = 0,481). Pour la marge supérieure (de 1 419 m à 1 477 m), les
résultats de l’AMOVA indique que 4 % (p = 0,028) de la variabilité est expliquée par les
différences inter-population.
Les différentes comparaisons des sous-populations ont toujours montré des différences non significatives, ou moindre (1 %, p = 0,262) dans le cas d’une marge supérieure élargie
(de 1 366 m à 1 477 m).
3) Comparaison des structures génétiques et géographiques
Le test de Mantel ne montre pas de corrélation significative entre la distance géographique et la distance génétique de Nei (Z = 0,038, p = 0,373). De même, je n’ai trouvé
aucune corrélation lors de la comparaison entre la marge inférieure du transect (Z = 0,022, p
= 0,280) ou la marge supérieure (Z = 0,022, p = 0,203) avec le reste du peuplement.
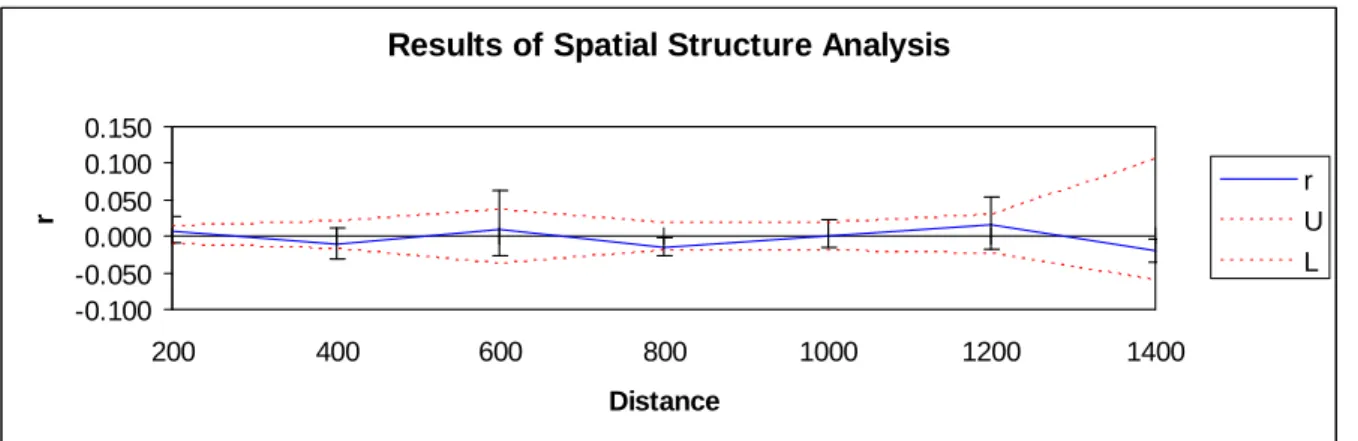
L’analyse de la structure spatiale des distances génétiques sur l’ensemble du gradient (Figure 10) ne montre pas de résultat significatif et confirme ceux du test de Mantel.
Results of Spatial Structure Analysis
-0.100 -0.050 0.000 0.050 0.100 0.150 200 400 600 800 1000 1200 1400 Distance r r U L
Figure 10 : Structuration des distances génétiques en fonction des distances géographiques pour l’ensemble du
III)
Production des graines
1) Effet maternel
La variabilité intra-arbre permet d’expliquer environ les ¾ de la variabilité de la production des graines. Sur la part de variation restante, l’effet mère en explique 80 %, et les facteurs qui ont été introduits dans le modèle seulement 20 % en moyenne (tableau 4).
Variable Variabilité intra
arbre p-value
Répartition de la variabilité
inter arbre p-value
Effet mère 78,2 %
Nombre de graine par
cône 68,7 % 1.10 -171 Facteurs 21,8 % 9.10-97 Effet mère 83,6 % % de graines plates 81 % 1.10-217 Facteurs 16,4 % 2.10 -167 Effet mère 82,9 % % de graines fécondées 77,5 % 4,5.10 -14 Facteurs 17,1 % 6.10 -11 Effet mère 76,9 % % de graines parasitées 83,3 % 1.10-272 Facteurs 23,1 % 4.10 -189 Effet mère 80,8 % % des pertes postzygotiques 76,1 % 2.10 -269 Facteurs 19,2 % 5.10 -169 Effet mère 81,1 % % de graines pleines 69 % 7.10-169 Facteurs 18,9 % 1.10 -112
Tableau 4 : Répartition de la variance des taux de graines
2) Effet des différents facteurs sur les variables
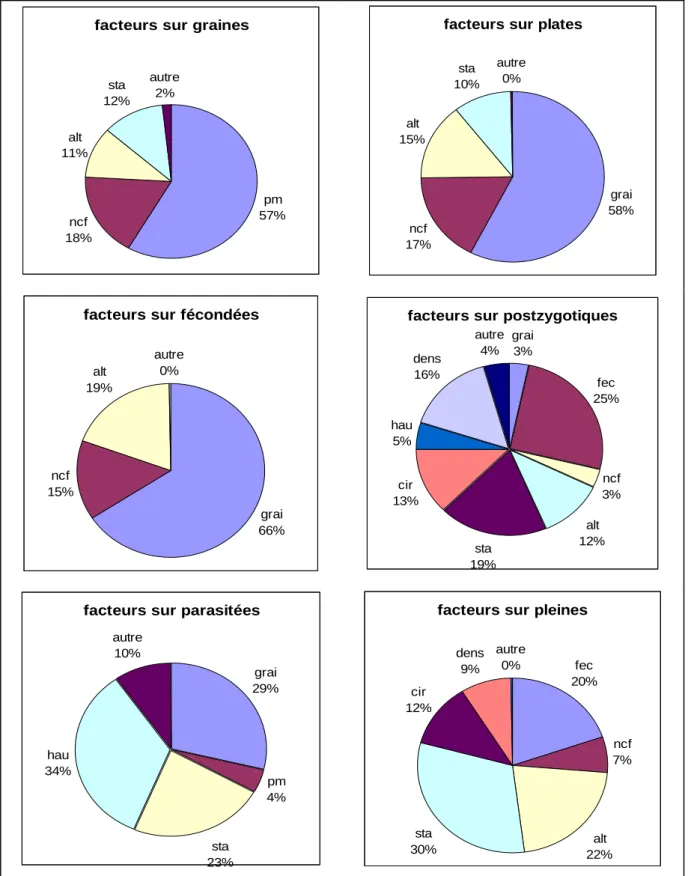
Le nombre de graines par cône est expliqué principalement par le nombre de chatons mâles (57 %) et le nombre de cônes femelles (18 %), c'est-à-dire par des caractéristiques en rapport avec la capacité de reproduction. Le nombre de graines par cône semble ainsi être une caractéristique propre aux gros reproducteurs. Les arbres dominés produisent moins de graines par cône et l’altitude a un impact significatif mais faible.
Le taux de graines plates diminue significativement lorsque le nombre de graines par cône (58 %) augmente et qu’il y a plus de cônes femelles sur l’arbre (17 %). Les arbres dominés ont plus de graines plates (10 %). Lorsque je ne tiens pas compte de l’effet du nombre de
graines par cône, j’explique une part plus importante de la variation par le nombre de cônes femelles (30 %), du statut social (21,7 %) et un effet négatif de la production de pollen (4 %).
facteurs sur graines
ncf 18% alt 11% sta 12% autre 2% pm 57%
facteurs sur plates
grai 58% ncf 17% alt 15% sta 10% autre 0%
facteurs sur fécondées
grai 66% ncf 15% alt 19% autre 0%
facteurs sur postzygotiques
grai 3% fec 25% ncf 3% alt 12% sta 19% cir 13% hau 5% dens 16% autre 4%
facteurs sur parasitées
grai 29% pm 4% sta 23% hau 34% autre 10%
facteurs sur pleines
fec 20% ncf 7% alt 22% sta 30% cir 12% dens 9% autre 0%
Le taux de graines fécondées augmente proportionnellement avec le nombre de graines par cône (66 %), l’altitude (19 %) et le nombre de cône femelles (15 %). Si l’on ne prend pas en compte l’effet des graines, on trouve un effet plus important du nombre de cônes femelles (26 %) et un effet de la production de pollen (22,6 %), alors que l’altitude n’est plus significative.
Le taux de pertes postzygotiques est influencé négativement par de nombreux facteurs : le taux de graines fécondées (25 %), le statut dominant (19 %), la densité locale (16 %), la circonférence (13 %), l’altitude (12 %), le nombre de cônes femelles (7 %), la hauteur (5 %) et le nombre de graines par cône (3 %). J’ai ensuite étudié l’effet des facteurs environnementaux en retirant celui du taux de graines fécondées. Les résultats sont similaires avec une contribution plus importante pour tous les facteurs et en particulier pour le statut dominant (32,8 %) et le nombre de cônes femelles (14,4 %).
La hauteur de l’arbre (34 %) et le statut dominé (23 %) ont un effet positif sur le taux de graines parasitées. Les facteurs ayant un effet négatif sont le nombre de graines par cône (29 %), le statut dominant (23 %), et la production de chatons mâles (4 %).
Le taux de graines pleines est principalement expliqué par le statut de l’arbre avec une augmentation significative chez les individus dominants (30 %) et l’altitude (22 %). Cette valeur est également favorisée par le taux de fécondation (20 %), la circonférence (12 %), la densité locale (9 %) et le nombre de cônes femelles (7 %). En ne tenant pas compte du taux de fécondation, l’effet de l’individu augmente significativement (45 %).
3) Effet de l’altitude & optimum
J’ai ajusté mes données sur un modèle basé sur une fonction de type parabolique. On trouve ici un optimum écologique situé vers 1 190 m pour le taux de graines fécondées et le taux de graines pleines, et on constate un minimum écologique à 1 283 m pour le taux de graines plates et le taux de pertes postzygotiques. L’inversion des courbes aux mêmes optimums pour le taux de graines plates et le taux de graines fécondées, ainsi que le taux de pertes prézygotiques et le taux de graines pleines, est du au mode de calcul de ces derniers et de leur complémentarité vis à vis du nombre total de graines par cône ou du nombre de graines fécondées (tableau 5).
Une seconde approche statistique a permis de préciser les optimums observés. Le jeu de données a été divisé en 2 parties à partir de l’optimum présumé pour chaque variable, puis la
linéarité des variables en fonction de l’altitude sur le bas et sur le haut du transect a été testée. Les coefficients de corrélation ainsi que leur significativité sont résumés dans le tableau 4.
Nb. graines % de plates % fécondées % post % pleines
Courbe ajustée
Optimum 1 340 m 1 190 m 1 185 m 1 284 m 1 283 m
Coef bas 0,055 NS -0,0037 * 0,0027 · -0,0038 *** 0,0035 ***
Coef haut -0,46 ** 0,003 ** -0,0024 ** 0,0024 NS -0,0032 ·
Tableau 5 : Comportement des variables en fonction de l’altitude
Une anova entre la régression linéaire et la régression polynomiale ne montre pas de différence significative. L’impact de l’altitude est modéré : l’augmentation ou la diminution des différents taux de graines est de l’ordre de 0,3% pour 100 m d’altitude. Au niveau du nombre de graines par cône, on observe une augmentation non significative d’environ 5 graines pour 100 m, et une diminution moyenne de l’ordre de 50 graines pour 100 m sur le haut du transect. 1000 1100 1200 1300 1400 1500 -2 -1 0 1 2 alt lo g (p le i. p/( 1 p le i.p )) 1000 1100 1200 1300 1400 1500 -2 -1 0 1 2 alt lo g(p ost .p /( 1 p o st .p )) 1000 1100 1200 1300 1400 1500 0 .0 0.5 1.0 1.5 2.0 2 .5 3 .0 alt lo g(f ec .p /( 1 fe c.p )) 900 1000 1100 1200 1300 1400 1500 -4 -3 -2 -1 0 1 alt lo g(p la .p /(1 - pla .p )) 1000 1100 1200 1300 1400 15 0 2 00 2 50 30 0 35 0 alt g ra i
Discussion
1) Dynamique de la recolonisation du mont Ventoux par le sapin pectiné
Les populations de sapin sur le mont Ventoux sont assez récentes et sont dans une phase de recolonisation depuis un peu plus d’un siècle. La structure génétique nous montre que le transect bas et le transect haut forment bien une seule population. En revanche, il existe une différenciation entre la population la plus haute en altitude et le reste du peuplement. Il pourrait s’agir de 2 sous-populations. L’estimation de l’âge des individus du transect, à partir des cernes (Maxime Cailleret, stage de M2), va de 30 ans à 159 ans. Sur le haut du transect, autour de 1 350 m, on retrouve des individus plus âgés, ce qui nous laisse supposer qu’il pourrait s’agir d’individus « reliques » ayant contribués à la recolonisation du mont Ventoux. Les données génétiques ne vont cependant pas tout à fait dans ce sens puisque la diversité génétique est justement inférieure chez les individus du haut du transect. On pouvait s’attendre à avoir une perte progressive de la diversité du haut vers le bas du transect, le long de l’hypothétique front de colonisation. Or, on ne retrouve pas non plus, de corrélation entre la diversité génétique des individus et leurs distances géographiques.
L’explication la plus probable est que la recolonisation s’est effectuée à partir de plusieurs populations « reliques » de diversité faible, notamment à plus basse altitude, ce qui a permis de maintenir une diversité génétique importante chez les jeunes individus du transect issus du mélange de plusieurs populations « relique ». De nombreux allèles sont d’ailleurs présents uniquement chez les jeunes individus et ne peuvent provenir des individus âgés du haut du transect. Ce résultat confirme l’importance du rôle des méta-populations dans le maintien de la diversité génétique ; il serait cependant intéressant d’effectuer une étude plus large autour du transect afin de vérifier la présence d’autres arbres « reliques ».
2) Dynamique actuelle et future du sapin pectiné
La colonisation du sapin est liée en grande partie au fait qu’il se réimplante dans des espaces où il devait être naturellement présent. On l’observe actuellement à partir de 900 m jusqu’à 1500 m, et son optimum écologique a été établi autour de 1 200 m (Sagnard, 2001). Les résultats que j’ai obtenus sur la production de graines montrent que cet optimum se situe
entre 1 200 m et 1 300 m. Il semble donc y avoir un décalage de l’optimum écologique du sapin vers une altitude plus élevée. Cette notion d’optimum écologique doit tout de même être prise avec modération. Au regard des coefficients de corrélation (tableau 5), il y a bien un changement de comportement des variables en fonction de l’altitude et elles n’évoluent pas de façon strictement monotone, mais l’optimum s’apparente souvent à une phase de stagnation. Le manque de donnée entre 1 132 m et 1 241 m ne devrait pas a priori modifier considérablement l’existence de l’optimum, juste le décaler légèrement. En revanche, la position de ces données manquantes peut accentuer la structuration polynomiale ou linéaire des variables en fonction de l’altitude. L’effet de l’altitude peut donc changer en terme de significativité.
Au niveau des limites du transect, si le nombre de graines est similaire, je trouve en revanche une proportion de graines pleines supérieure au niveau de la marge supérieure par rapport à la marge inférieure. La constatation d’un dépérissement important du sapin principalement sur le bas du transect, causé semble-t-il par des conditions climatiques plus sèches, conforte l’idée que le sapin pectiné qui se trouve en limite d’aire sur le mont Ventoux, est peut-être en recul, et qu’il est poussé à se réfugier à une altitude supérieure. Une étude réalisée sur la croissance du sapin pectiné le long du même gradient par Maxime Cailleret, montre également, d’une part l’existence d’une croissance optimale entre 1 200 m et 1 300 m, et d’autre part que cet optimum s’est aussi décalé vers le haut du transect.
Avant de pouvoir émettre des conclusions sur la dynamique du sapin pectiné au niveau de son aire globale de répartition en tenant compte des changements climatiques annoncés, il faudra regarder de manière plus spécifique les résultats observés sur le mont Ventoux avec des données climatiques (températures, précipitations, etc.).
3) Effet maternel sur le succès reproducteur femelle
Je trouve un effet majeur de la mère, qui explique environ 80 % de la variabilité de la production des graines, ce qui confirme les précédentes études (Kärkkäinen & al., 1999). Il faut préciser que cet effet mère regroupe également l’effet des facteurs qui n’ont pas été pris en compte dans cette étude, comme la quantité de lumière reçue, la composition physico chimique du sol, les conditions climatiques journalières et en particulier l’effet de la densité des individus reproducteurs qui influe de manière très significative, d’une part sur la constitution du nuage pollinique ( Robledo & al., 2004) et donc le taux de graines vides, et
la part de la variabilité due à la mère reste considérable et, au niveau de la gestion forestière, cela confirme la nécessité de maintenir une diversité génétique importante pour espérer avoir des arbres avec une bonne production de graines et donc une dynamique permettant de maintien du peuplement.
Il faut préciser qu’il y a aussi une très forte variabilité de la production de graines entre les individus et surtout entre les années (Krouchi & al., 2004 ; Sagnard, 2001). Outre le fait qu’il y a une alternance entre des années de forte production de graines et des années de faible production (Lazaro & al., 2006), il semblerait que les individus eux-mêmes n’ont pas une production de graines constante chaque année (Krouchi & al., 2004 ; Restoux, thèse en cours). Les résultats obtenus sur la production de graines, en relation avec l’altitude notamment, peuvent donc être en partie dus à l’échantillonnage des individus. Dans ce cas, un suivi sur plusieurs années s’avère nécessaire pour comprendre si les différences inter-individuelles persistent lors du cumul de plusieurs épisodes de reproduction.
4) Effet des facteurs du milieu sur la production de graines
a) Nombre de graines
Les arbres qui produisent beaucoup de graines sont ceux qui produisent également beaucoup de pollen et de cônes femelles. Aucun des autres facteurs n’a d’influence sur ce nombre de graines par cône, notamment la circonférence et la hauteur (Mosseler & al., 2000). Il ressort tout de même que la classe phytosociologique revêt une grande importance, les arbres dominés produisant moins de graines, tout comme ils produisent moins de pollen et moins de cônes femelles que les arbres dominants. A la vue de ces résultats, il me semble que l’on peut distinguer d’une part les arbres qui ont investi dans la reproduction, et d’autre part ceux qui ont investi dans d’autres processus, et ce indépendamment des facteurs externes. Il existe en effet un trade-off au niveau de l’arbre entre l’investissement dans la reproduction et l’investissement dans la croissance que ces résultats pourraient confirmer (Cailleret, stage M2, 2007). Les arbres dominants auraient quant à eux une plus forte probabilité d’investir dans la reproduction que les arbres dominés. Toutefois, lorsque l’on regarde les effets indépendamment, on constate que la hauteur et la circonférence sont les facteurs les plus influents. On peut donc penser que l’effet lié au pollen et à la production des cônes femelles inclut l’effet de la circonférence et de la hauteur. Ainsi, le nombre de graines serait tout de même lié à la hauteur et à la circonférence de l’arbre bien que ces deux facteurs n’aient pas
b) Taux de fécondation & pollinisation
Les taux de graines plates et fécondées sont principalement expliqués par le nombre de graines par cône. Cet effet semble important car il inclut la variabilité expliquée par d’autres facteurs (nombre de cônes femelles, production de pollen, etc). En ne prenant pas en compte l’effet du nombre de graines par cône, je trouve un effet de la production de pollen. Cela peut être interprété comme la contribution non négligeable de l’autofécondation qui augmente la pollinisation et permet de diminuer le taux de graines plates. D’autre part, la forte influence du nombre de graines par cône pourrait masquer l’effet important de la structure du nuage pollinique. Dans nos hypothèses de départ, l’importance du nuage pollinique dépend beaucoup de la densité locale en reproducteur, donnée indisponible pour cette étude. Or à forte densité, le nombre de cônes femelles diminue tandis que le nombre de graines par cône augmente. On peut donc interpréter l’effet du nombre de graines, comme l’effet indirecte de la densité. Le taux de graines plates diminue lorsqu’il y a une forte densité de reproducteurs et que la taille du nuage pollinique est plus importante.
c) Dépression de consanguinité
Les pertes postzygotiques sont représentatives de la dépression de consanguinité. Le fait que la production de pollen de l’individu n’a pas d’influence significative sur ce taux semble indiquer que la dépression de consanguinité n’est pas due majoritairement à de l’autofécondation. De plus, le long du transect, les sapins sont souvent plus hauts que les autres essences qui peuvent diminuer l’autofécondation en limitant l’ascension verticale du pollen (les fleurs mâles se situent sur les branches basses du houppier), ce qui pourrait expliquer l’effet négatif de la densité locale. Ce serait dès lors l’apparentement entre les individus qui serait principalement responsable des pertes postzygotiques. Il semble donc relativement cohérent que l’on trouve comme facteurs explicatifs ceux qui permettent aux arbres d’avoir accès au nuage pollinique : circonférence (fortement corrélée avec la hauteur) et statut phytosociologique dominant.
En revanche, un effet fort de la densité locale en reproducteur (Mosseler & al., 2004), associé au nombre de graine par cône était attendu. Or celui-ci n’a pas d’effet significatif, on observe même un effet négatif du nombre de cônes femelles. Cela remet en cause l’estimation de la densité en reproducteur par le nombre de graines par cône, et il me parait essentiel à l’avenir de mesurer cette dernière directement.
d) Graines parasitées
Le taux de graines parasitées est expliqué en premier par la hauteur de l’arbre. L’influence de la hauteur est liée au mode de déplacement de Megastigmus suspectus. Ce parasite se laisse initialement porter par les vents, puis il s’oriente vers son hôte sur le critère visuel, d’où l’importance de la hauteur. Pour ce qui est de l’influence du nombre de graine en tant qu’approche de la densité en reproducteurs, nos résultats ne vont pas exactement dans le sens de ce que l’on attendait. Je trouve en effet qu’à forte densité le taux de parasitisme est le plus faible alors que ce dernier devrait être à une densité moyenne. L’estimation de la densité par le nombre de graines par cône peut une nouvelle fois être mise en doute. Il est possible aussi que la densité maximale du peuplement du transect, qui est en phase de recolonisation, corresponde à une densité moyenne pour un peuplement mature, auquel cas mes résultats correspondraient à la littérature (rapport Invagraine).
e) Taux de reproduction
Il est intéressant de constater que le taux de reproduction (graines pleines) est peu influencé par la capacité de reproduction de l’individu (production de chatons mâles et de cônes femelles), mais dépend essentiellement, du statut dominant de l’arbre. Le sapin pectiné est très majoritairement allofécondé, ce qui explique que la production de pollen n’influe pas sur la reproduction de l’individu en soit. L’autofécondation étant suivie d’une augmentation des avortements postzygotiques, le nombre de graines pleines ne doit pas être si différents d’un individu ayant moins de graines pollinisées par l’allofécondation et donc moins de pertes postzygotiques. En revanche, le statut dominant peut permettre à l’arbre d’avoir accès plus largement au nuage pollinique et ainsi favoriser sa pollinisation avec une majorité d’allopollen, favorisant ainsi la fécondation de ses graines tout en limitant les risques de dépression de consanguinité.