DÉTERMINANTS ET COÛTS À LONG TERME DE LA REPRODUCTION EN JEUNE ÂGE CHEZ LES MOUFLONS D'AMÉRIQUE MÂLES
par
Yanny Ritchot
Mémoire présenté au Département de biologie en vue de l’obtention du grade de maître ès sciences (M.Sc.)
FACULTÉ DES SCIENCES UNIVERSITÉ DE SHERBROOKE
Le 20 mars 2021
Le jury a accepté le mémoire de Monsieur Yanny Ritchot dans sa version finale.
Membres du jury
Professeure Fanie Pelletier Directrice de recherche Département de biologie Université de Sherbrooke
Professeur Marco Festa-Bianchet Codirecteur de recherche
Département de biologie Université de Sherbrooke
Professeur Marc Bélisle Évaluateur interne Département de biologie Université de Sherbrooke
Professeur Dany Garant Président-rapporteur Département de biologie Université de Sherbrooke
REMERCIEMENTS
J’aimerais premièrement remercier mes directeurs, Pre Fanie Pelletier et Pr Marco Festa-Bianchet. Merci pour tout le temps et l’attention que vous avez mis pour la réalisation ce projet. Merci pour tous ces échanges autant en personne, en vidéo-conférence ou même au téléphone du haut de la montagne sous la tente-prospecteur pour tenter de finaliser notre manuscrit. Votre support et vos conseils m’ont grandement aidé et c’est très apprécié. Merci aussi aux membres de mon comité, Marc Bélisle et Dany Garant, qui ont apporté des suggestions intéressantes au cours de ma maîtrise. Un grand merci également à Pr David Coltman pour ton aide sur la rédaction du manuscrit. Merci à François Rousseu pour tes conseils sur les analyses statistiques et tes pouvoirs magiques pour résoudre les casse-têtes les plus complexes dans R.
Merci aux membres des labos Pelletier et Festa-Bianchet pour vos conseils, suggestions et supports tout au long de ce projet. Merci également à tous les membres des autres laboratoires d’écologie pour votre support moral. Un remerciement spécial à Benjamin, Cédric, alias « Peepo », Roxane, Emmanuelle et Denis avec qui j’ai partagé des moments inoubliables à Ram Mountain.
Finalement, j’aimerais remercier le CRSNG et l’ACA pour leur soutien financier et qui ont rendu ce projet réalisable.
SOMMAIRE
Comme les ressources sont limitées, la théorie des histoires de vie suppose qu’un individu doit faire des compromis dans la façon dont il alloue les ressources acquises. Une allocation à la reproduction diminue les ressources disponibles à la survie ou à la croissance, ce qui signifie que la reproduction a un coût. Cependant, puisque la capacité d’acquisition des ressources varie d’un individu à l’autre au sein d’une population, les coûts de la reproduction seront moins ressentis pour les individus avec une capacité d’acquisition supérieure. Chez les espèces polygynes et longévives, le succès reproducteur des mâles est généralement monopolisé par quelques individus adultes et dominants. Les jeunes subordonnés peuvent donc utiliser des tactiques alternatives pour réussir à s’accoupler. Cela dit, peu d’études se sont intéressées aux déterminants du succès reproducteur en jeune âge. Pourtant, la variabilité dans la participation à la reproduction en jeune âge est un élément clé pour comprendre la force de la sélection sexuelle et les compromis d’allocation potentiels sur différents traits d’histoire de vie. En fait, une grande proportion de la littérature dans ce domaine porte sur les femelles, alors qu’il y a peu d’information sur les coûts de la reproduction chez les mâles chez les espèces où les soins parentaux sont exclusivement prodigués par les femelles. De plus, les connaissances acquises sur les histoires de vie d’un sexe ne peuvent pas être appliquées pour l’autre puisque les déterminants et les stratégies de reproduction vont généralement différer. La paternité doit également être assignée à l’aide de matériel génétique, ce qui limite l’accessibilité aux données et contribue au manque d’études. De plus, l’hétérogénéité interindividuelle au niveau de l’acquisition des ressources accentue ce problème puisqu’elle peut également mener à une corrélation positive entre deux traits à l’échelle populationnelle si la variance en acquisition des ressources est grande, mais que la variance en allocation est faible. Ceci tend à masquer les coûts de la reproduction et justifie pourquoi peu de résultats ont été observés dans les systèmes d’étude où le matériel génétique est disponible. L’objectif principal de mon mémoire était d’approfondir les connaissances sur les coûts de la reproduction chez les mâles des espèces polygynes. Plus précisément, mon projet visait à évaluer les déterminants de la reproduction en jeune âge chez les mâles d’une population sauvage de mouflons d’Amérique (Ovis canadensis) et évaluer leurs
x effets à long terme sur la longévité et le succès reproducteur total. En utilisant 30 ans de données à l’échelle individuelle, j’ai pu montrer qu’une augmentation de la masse corporelle, de la proportion des jeunes de 2-4 ans parmi les mâles adultes et du nombre de femelles par mâle (sexe-ratio d’accouplement) avaient tous un effet positif sur le succès reproducteur à trois ans. À deux ans, seulement 6% des mâles se sont reproduits et aucuns déterminants étudiés n’influençaient la probabilité de se reproduire, ce qui suggère que le succès reproducteur à cet âge est un événement stochastique. À quatre ans, aucun déterminant n’était statistiquement significatif, mais les tendances étaient similaires à celles observées chez les mâles de trois ans. Ces résultats suggèrent que le succès reproducteur en jeune âge dépend fortement de la démographie et des capacités d’un individu à compétitionner. J’ai aussi pu montrer que les individus s’étant reproduits avant l’âge de cinq ans avaient un succès reproducteur à vie plus élevé et une longévité réduite. Ceci suggère un coût à long terme de la reproduction en début de vie. Finalement, mon projet fait partie des rares études qui ont étudié les compromis d’allocation chez les mâles d’une espèce polygyne et renforce l’hypothèse sur la dichotomie dans les déterminants et les stratégies de reproduction entre les sexes.
Mots-clés : Ovis canadensis, compromis d’allocation, polygynie, coûts de la reproduction, tactiques de reproduction, âge à la première reproduction
TABLE DES MATIÈRES
CHAPITRE 1 INTRODUCTION GÉNÉRALE ... 1
1.1 Contexte général ... 1
1.1.1. Théorie des histoires de vie ... 1
1.1.2. Allocation et acquisition des ressources ... 1
1.1.3. Objectif général et objectifs spécifiques ... 4
1.2 Sélection sexuelle ... 5
1.2.1. Anisogamie ... 5
1.2.2. Théorie de la sélection sexuelle ... 7
1.2.3. Sélection intrasexuelle ... 7
1.2.4. Sélection intersexuelle ... 8
1.3 Principe de Bateman et l'investissement parental ... 10
1.4 Stratégies de reproduction et tactiques de reproduction alternatives ... 12
1.5 Investissement dans la reproduction ... 13
1.6 Système d'étude ... 14
1.6.1. Le mouflon d'Amérique ... 14
1.6.2. Tactiques de reproduction du mouflon d’Amérique ... 15
1.6.3. Chasse aux trophées ... 16
1.6.4. Population de Ram Mountain ... 17
CHAPITRE 2 DÉTERMINANTS ET COÛTS À LONG TERME D'UNE REPRODUCTION EN JEUNE ÂGE CHEZ UN MÂLE D'UN MAMMIFÈRE POLYGYNE ET LONGÉVIF ... 21
2.1 Description de l’article et contribution ... 21
2.2 Abstract ... 23
2.3 Significance Statement ... 24
2.4 Introduction ... 25
2.5 Material and Methods ... 29
2.5.1. Study area and population ... 29
xiii
2.5.3. Statistical analyses ... 30
2.6 Results ... 34
2.6.1. Body mass candidate metrics ... 34
2.6.2. Determinants of reproductive success between two and four years of age ... 34
2.6.3. Long-term effects of age at first reproduction on fitness ... 37
2.7 Discussion ... 42 2.8 Acknowledgements ... 47 2.9 Ethical statement ... 47 2.9.1. Funding ... 47 2.9.2. Conflict of interests ... 47 2.9.3. Ethical approval ... 47 2.9.4. Informed consent ... 47
2.9.5. Availability of data and material ... 48
2.9.6. Authors’ Contributions ... 48
2.10 References ... 49
CHAPITRE 3 DISCUSSION ET CONCLUSION ... 57
3.1 Retour sur les résultats ... 57
3.2 Discussion générale ... 57 3.3 Limitations de l’étude ... 60 3.4 Perspectives futures ... 61 3.5 Conclusion ... 63 ANNEXE ... 65 BIBLIOGRAPHIE ... 78
LISTE DES TABLEAUX
Table 2.1 Coefficient estimates of the effects of body mass, age structure, sex ratio, population density and number of lambs sampled for DNA on the reproductive success of bighorn sheep males aged two, three and four years at Ram Mountain, Alberta during the ruts of 1987-2017 ...
36 Table 2.2 Coefficient estimates of the effects of age at first reproduction
(AFR) and body mass at two years (kg) on lifetime reproductive success of bighorn sheep males at Ram Mountain, Alberta during the ruts of 1987-2017 ...
38 Table 2.3 Coefficient estimates of the effects of age at first reproduction
between two and four years and body mass at two years on the longevity of 23 bighorn sheep males at Ram Mountain, Alberta during the ruts of 1987-2017 ...
41 Table S1 Model selection for body mass effects on reproductive success of
bighorn sheep males aged 2-4 at Ram Mountain, Alberta, ruts 1987-2017 ... 65 Table S2 Model selection for determinants of reproductive success of
two-year-old males at Ram Mountain, Alberta, Canada, ruts 1987-2017 68 Table S3 Model selection for determinants of reproductive success of
three-year-old males at Ram Mountain, Alberta, Canada, ruts 1987-2017 70 Table S4 Model selection for determinants of reproductive success of
xvi Table S5 Model selection for lifetime reproductive success as a function of
age at first reproduction for bighorn sheep males at Ram Mountain, Alberta, ruts 1987-2017 ... 73 Table S6 Model selection for lifetime reproductive success as a function of
age at first reproduction between two and four years for bighorn sheep males at Ram Mountain, Alberta, ruts 1987-2017... 73 Table S7 Model selection for longevity as a function of age at first
reproduction and body mass at two years for bighorn sheep males . 74 Table S8 Model selection for longevity as a function of age at first
reproduction between two and four years and body mass at two years ... 74 Table S9 Effects of age at first reproduction and body mass at two years on
the longevity of bighorn sheep males at Ram Mountain, Alberta .... 75 Table S10 Model selection for lifetime reproductive success as a function of
early or late age at first reproduction for bighorn sheep males at Ram Mountain, Alberta, ruts 1987-2017 ... 76
LISTE DES FIGURES
Figure 1.1. Corrélation positive entre les traits d’histoire de vie. ... 3 Figure 1.2. Critères de légalité pour la récolte de mouflons d’Amérique en
Alberta ... 17 Figure 2.1 Age distribution of reproductive success for 141 bighorn rams aged
two years and older at Ram Mountain, Alberta during the ruts of 1987-2017 ...
33 Figure 2.2 Probability to reproduce as a function of body mass, sex ratio, age
structure and population density for young bighorn sheep males at Ram Mountain, Alberta during the ruts of 1987-2017 ... 35 Figure 2.3 Effect of age at first reproduction (AFR) on lifetime reproductive
success (LRS) and longevity for bighorn sheep males at Ram Mountain, Alberta during the ruts of 1987-2017 ... 40
LISTE DES ABRÉVIATIONS
AIC Critère d’information d’Akaike (Akaike Information Criterion)
AICc Critère d’information d’Akaike corrigé (Second-order Akaike Information Criterion)
AFR Âge à la première reproduction (Age at First reproduction) BSR Sexe-ratio d’accouplement (Breeder Sex Ratio)
CI Intervalle de confiance (Confidence Interval)
DNA Acide Désoxyribonucléique (DeoxyriboNucleic Acid)
K Nombre de paramètres
LRS Succès reproducteur total (Lifetime Reproductive Success)
NSERC Conseil de recherches en sciences naturelles et en génie (Natural Sciences and Engineering Research)
SE Erreur type (Standard Error)
CHAPITRE 1
INTRODUCTION GÉNÉRALE
1.1 Contexte général
1.1.1. Théorie des histoires de vie
La théorie des histoires de vie est une branche de la biologie évolutive qui cherche à expliquer comment les individus allouent leurs ressources selon différentes variables, comme la disponibilité de celles-ci, le sexe et l’âge des individus, afin de se reproduire avec succès (Stearns 2000). En nature, une multitude de stratégies de reproduction peut être observée entre les organismes, soit un ensemble de décisions et de règles sous une base génétique sur la façon d’allouer des ressources pour favoriser le succès reproducteur (Gross 1996). Cette diversité suscite des questions depuis des années. Par exemple : pourquoi certaines espèces sont de grandes tailles, alors que d’autres sont petites? Pourquoi le processus d’atteignement de la maturité sexuelle est parfois rapide et parfois lent? Pourquoi certaines espèces ont un jeune par événement de reproduction, alors que d’autres en ont des centaines? Pourquoi la durée de vie est si variable d’une espèce à l’autre (Stearns 2000)? Pour répondre à ces questions, il faut mesurer et comparer les traits d’histoire de vie des organismes, c’est-à-dire tout événement lié à la survie, la reproduction ou la croissance qui a le potentiel d’influencer l’aptitude phénotypique (Hill 1993, Stearns 1977). Comprendre la diversité observée entre les organismes est cruciale pour ensuite comprendre l’importance de la présence et de la quantité de variance génétique (Stearns 1992).
1.1.2. Allocation et acquisition des ressources
En milieu naturel, l’énergie disponible à allouer à la reproduction, la croissance ou la survie est limitée par les ressources disponibles et acquises par un individu. Ceci impose un compromis
2 dans l’allocation des ressources entre différents traits (Stearns 1992). Par exemple, une étude sur les écureuils roux (Tamiasciurus hudsonicus) a montré que les femelles ayant acquis plus de ressources avaient un âge de première reproduction plus précoce. Ceci augmentait leur aptitude phénotypique, malgré le coût engendré sur la durée de vie (Descamps et al., 2006). Cet exemple montre un coût de la reproduction issu d’un compromis d’allocation entre reproduction et survie, où les femelles qui allouaient plus tôt à la reproduction avaient moins de ressources à allouer au maintien de la condition corporelle, ce qui se traduisait par une mortalité précoce.
L’âge auquel un individu commence à se reproduire n’est pas la seule composante du succès reproducteur. En effet, d’autres traits d'histoire de vie, tels que la taille des portées, sont souvent étroitement liés à l'aptitude phénotypique d'un individu (Clutton-Brock et al. 1982, Harper 1977, Warner 1984). Les coûts liés à la reproduction ont été explorés sous plusieurs angles chez les femelles de différents taxons chez les animaux et les végétaux (Jorgenson et al. 1997, Roberts et al. 1995, Silk et al. 2003, Yanagi et Miyatake 2003). D’ailleurs, Lemaître et al. (2015) ont montré que 85.3% des études sur les compromis d’allocation entre les traits de début et fin de vie portaient sur les femelles.
La littérature scientifique contient beaucoup moins d’information sur les coûts de la reproduction chez les mâles, surtout chez les espèces polygynes où les soins parentaux sont principalement prodigués par les femelles (Schubert et al. 2009). Effectivement, chez ces espèces, le lien de paternité doit être identifié à l’aide d’outils moléculaires, ce qui limite les recherches au niveau logistique et monétaire (Hamel et al. 2010). Toutefois, quelques études ont porté sur les coûts de la reproduction chez les mâles des espèces polygynes. D'ailleurs, Hamel et al. (2010) ont recensé un total de 12 populations de mammifères où les coûts de la reproduction des mâles faisaient l’objet d’étude, dont seulement sept sur les grands mammifères. Plus récemment, quelques études se sont ajoutées, par exemple sur l’orignal (Alces alces, Markussen et al. 2019) et sur l’éléphant de mer du Sud (Mirounga leonina, Lloyd et al. 2020). L’hétérogénéité dans l’acquisition des ressources est un autre facteur qui explique pourquoi peu de coûts de la reproduction ont été rapportés dans la littérature (Hamel et al. 2010, van
3 Noordwijk et de Jong 1986). En effet, bien qu’au niveau individuel une relation négative entre la reproduction et la survie et/ou reproduction future soit probable, cette relation au niveau populationnel peut être masquée (van Noordwijk et de Jong 1986) : ces coûts seront apparents seulement si la variation en acquisition est plus faible que la variation en allocation (Figure 1.1, Hamel et al. 2010). Des manipulations expérimentales en allocation peuvent permettre d’étudier ce phénomène, mais cette technique est difficile à appliquer chez les mammifères et encore plus chez les mâles (Hamel et al. 2010). Toutefois, Stevenson et Bancroft (1995) ont montré qu'il y avait un coût à la reproduction sur la survie chez le mouton Soay (Ovis aries) mâle en castrant certains individus pour éviter qu'ils participent au rut.
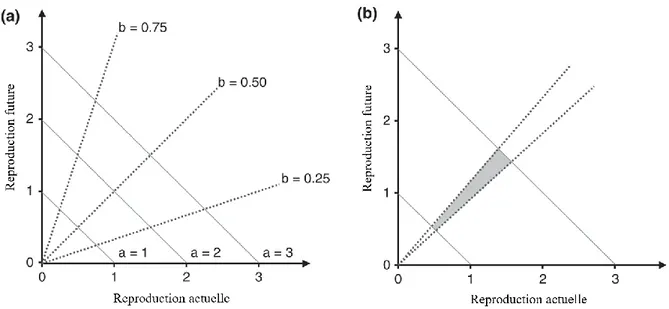
Figure 1.1. Corrélation positive entre les traits d’histoire de vie.
Les lignes pointillées du panneau a) illustrent différentes façons dont les individus allouer leurs ressources envers deux traits d’histoire de vie, ici
reproduction actuelle et future, alors que les lignes pleines montrent qu’il y a de la variance en acquisition entre les individus d’une population. Le panneau b) illustre la corrélation positive qui peut se former à l’échelle populationnelle
4 entre deux traits (zone colorée) malgré la relation négative inévitable à l’échelle individuelle. Cette dernière se produit si la variance en acquisition des
ressources dans une population est grande et la variation en allocation est faible. Figure traduite de Hamel et al. (2010) initialement créée par van Noordwijk et de Jong (1986).
De plus, l’information connue sur les coûts de la reproduction chez les femelles ne s’applique pas nécessairement aux mâles d’une même espèce, puisque les déterminants et les stratégies de reproduction vont généralement différer (Hoffman et al. 2008). La raison principale est que le succès reproducteur des mâles est généralement limité par l’accès aux femelles, alors que celui des femelles est souvent limité par les ressources (Emlen et Oring 1977). De plus, les mâles chez les espèces qui pratiquent la promiscuité ou la polygynie ont un potentiel reproducteur plus élevé que chez les espèces monogames, ce qui incite les mâles à prendre plus de risques pour s’accoupler (Tidière et al. 2015). Ceci mène également à une sélection pour une masse corporelle supérieure et au développement d’armements ou d’ornements. L’expression de ces traits est coûteuse à long terme et expliquerait pourquoi les mâles meurent plus jeunes que les femelles (Tidière et al. 2015). Il est donc critique d'augmenter nos connaissances sur les pressions de sélection agissant sur les mâles étant donné le manque de d’études sur le sujet et la dichotomie marquée dans les déterminants et les stratégies de reproduction entre les sexes chez les espèces polygynes.
1.1.3. Objectif général et objectifs spécifiques
L’objectif principal de ce projet était d’approfondir les connaissances sur les coûts de la reproduction pour les mâles d’une espèce polygyne, le mouflon d’Amérique (Ovis canadensis), pour laquelle des données détaillées sur l’âge de première reproduction et le succès reproducteur sont disponibles. De manière plus spécifique, ce projet avait comme buts :
1. D’explorer les différents déterminants phénotypiques et démographiques qui pourraient favoriser une reproduction en jeune âge chez les béliers.
5 2. D’évaluer si le succès reproducteur en jeune âge a un impact sur la longévité et le succès reproducteur total.
Établir les différents déterminants qui favorisent la reproduction en jeune âge permettra de mieux comprendre le maintien d’une certaine variabilité génétique dans un système d’accouplement majoritairement monopolisé par quelques individus. Une relation entre le succès reproducteur en jeune âge et la longévité suggérerait également un lien positif entre l’effort de reproduction en jeune âge et le succès. Dans les populations d’espèces polygynes, une hiérarchie sociale est généralement observée et les quelques mâles dominants obtiennent la majorité des paternités annuelles (Coltman et al. 2002, Hogg et Forbes 1997, McElligott et al. 2001). De plus, évaluer les causes et conséquences d’une allocation à la reproduction tôt dans la vie permettra d’approfondir nos connaissances sur les compromis d’allocation entre les traits de début et de fin de vie et ainsi mieux comprendre la senescence. Les résultats issus de mes travaux ouvriront également des pistes de recherches sur les différences dans les déterminants et les stratégies de reproduction entre les sexes, ce qui devrait accroître notre compréhension de l’évolution des histoires de vie et de la sélection sexuelle.
1.2 Sélection sexuelle
1.2.1. Anisogamie
Chez la plupart des espèces multicellulaires, deux tailles de gamètes distinctes sont observées selon le sexe, ce qu'on appelle l'anisogamie. L'origine de ce phénomène est encore sujette à débat, mais son importance dans l'évolution et les rôles sexuels est tout autrement bien acceptée (Bell 1978, Charlesworth 1978, Kalmus et Smith 1960, Parker et al. 1972). Bien qu'il n'y ait pas de théorie entièrement corroborée, quelques hypothèses sont relativement bien acceptées (Parker et al. 1972).
D'abord, Kalmus et Smith (1960) ont proposé comme hypothèse la sélection perturbatrice à partir de l'isogamie. Celle-ci suggère l'apparition d'une population homozygote pour un gène
6 récessif produisant des petits gamètes qui a vu apparaître un gène dominant pour la production de gros gamètes. Bien qu’un gros gamète ait des réserves énergétiques plus importantes, la fécondation avec un autre gros gamète serait presqu’impossible puisque leur taille diminue le mouvement et les rend presque statiques. Un équilibre entre petits et gros gamètes est donc nécessaire pour la survie de la population.
Une seconde hypothèse, proposée par Parker et al. (1972), explique l'apparition de l'anisogamie en considérant les forces sélectives majeures qui influencent la taille des gamètes. Parmi ces forces, il y a le taux de production de gamètes que peut avoir un parent par unité de temps et la probabilité qu’un zygote atteigne la maturité sexuelle et se reproduise à son tour. Ces forces se contraignent puisqu’une tend à limiter la taille des gamètes et l’autre cherche à l’augmenter pour emmagasiner un maximum de ressources. Il est donc nécessaire pour une espèce d'avoir les deux types de gamètes.
L'anisogamie serait possiblement à la base de l'origine des sexes (Charlesworth 1978). De plus, la masse d’un gamète produit correspond directement à l'énergie dépensée pour le produire, ce qui implique que les macrogamètes (ovocytes) demandent une allocation supérieure aux microgamètes (spermatozoïdes, Parker 2011). En outre, comme la productivité de spermatozoïdes est très élevée, seule une faible partie pourra se fusionner à un ovocyte pour former un zygote, initiant les premières traces d'une compétition intrasexuelle. L'anisogamie implique alors que les efforts énergétiques de chaque sexe dans la production de gamètes diffèrent, ce qui devrait influencer les stratégies de reproduction et devrait donc être à la base de la sélection sexuelle (Togashi et Cox 2011).
7 1.2.2. Théorie de la sélection sexuelle
En 1859, Darwin a publié son livre L'origine des espèces (Darwin 1859). Ce dernier a proposé que les espèces changent au cours du temps à la suite de changements héréditaires de traits morphologiques et comportementaux. La théorie de la sélection naturelle stipule qu'un individu mieux adapté à son environnement aura un potentiel reproducteur plus élevé (Darwin 1859). Cependant, dans certains cas, comme pour la queue majestueuse du paon bleu (Pavo cristatus) mâle, certains traits semblent nuire à la survie de l'individu. Pour résoudre ce paradoxe, Darwin (1871) a alors proposé la sélection sexuelle, où les individus d'une population portant des traits coûteux pour la survie auront un avantage reproductif sur ceux du même sexe qui n’arborent pas ces traits. Cet avantage provient généralement d'un trait phénotypique héritable favorisant l'accès à un partenaire même si ce trait nuit à la survie de l'individu. Autrement dit, si un trait peut augmenter le succès reproducteur de l’individu qui l’exprime malgré le coût de son expression, il devrait augmenter en fréquence dans la population (Darwin 1871).
Deux formes de sélections sexuelles sont présentes dans la nature : la sélection intrasexuelle et la sélection intersexuelle (Andersson 1994). La sélection intrasexuelle favorise les traits phénotypiques exagérés (souvent des traits sexuels secondaires) qui offrent un avantage dans la compétition intrasexuelle, alors que la sélection intersexuelle favorise les traits qui augmentent les chances d'être choisi par le sexe opposé (Borkovic et Russell 2014).
1.2.3. Sélection intrasexuelle
La sélection intrasexuelle s’observe lorsque des individus du même sexe entrent en compétition pour augmenter leur succès reproducteur (Andersson 1994). Ce mécanisme s'observe lorsqu'un sexe devient le facteur limitant au succès reproducteur du sexe opposé (Hosken et House 2011). Le sexe limitant sera celui qui investit le plus de ressources dans sa progéniture. De manière générale, il s'agit de la femelle. En effet, comme mentionné ci-haut, cette différence débute déjà à la formation des gamètes qui diffèrent en taille, où les femelles produisent des macrogamètes,
8 alors que les mâles produisent des microgamètes (Parker et al. 1972). Le succès reproducteur des mâles est donc généralement limité par la quantité de partenaires qu’ils peuvent féconder (Bateman 1948). De plus, chez de nombreuses espèces d'animaux, une période de gestation est présente chez la femelle, ce qui impose une demande énergétique supplémentaire (Clutton-Brock et al. 1989, Munns 2013). Plus l’investissement énergétique envers un jeune augmente pour la femelle, plus le degré de compétition intrasexuelle des mâles est sujet à augmenter. Par exemple, chez les mammifères, la lactation augmente le coût énergétique de la reproduction pour les femelles, ce qui diminue la disponibilité des partenaires pour les mâles et tend à favoriser la compétition intrasexuelle (Hosken et House 2011). D’ailleurs, plus l’asymétrie en succès reproducteur des mâles est élevée chez une espèce, plus la force de sélection pour des traits sexuels secondaires devrait être forte puisque la capacité compétitive d’un individu aura un impact plus fort sur son aptitude phénotypique (Promislow 1992).
1.2.4. Sélection intersexuelle
Darwin (1871) a également proposé que certains traits sexuels secondaires évoluent par une préférence du sexe limitant plutôt que par la compétition intrasexuelle. Un exemple classique de ce type de trait est la queue majestueuse du mâle chez le paon bleu, qui apporte un avantage compétitif individuel à celui qui exprime le plus fortement ce trait face aux autres mâles (Darwin 1871, Loyau et al. 2005). Un autre exemple serait les couleurs flamboyantes du guppy (Poecilia reticulata), où l’intensité de la coloration du mâle influence le choix de la femelle (Houde et Torio 1992). Ainsi, la sélection intersexuelle tend à favoriser les traits qui augmentent les chances d'être choisi par le sexe opposé (Borkovic et Russell 2014) et s’intensifie lorsque le sexe limitant préfère certains traits phénotypiques chez le sexe opposé (Borkovic et Russel 2014). L’expression de traits sexuels secondaires peut offrir un bénéfice direct ou indirect à l'autre sexe (Hamilton et Zuk 1982, Trivers 1972, Zahavi 1975). Un bénéfice direct augmente le potentiel reproducteur de l'individu, alors qu’un bénéfice indirect augmente le potentiel reproducteur de ses rejetons (Andersson et Simmons 2006). Cependant, ce concept n'explique pas comment les gènes responsables de la préférence puissent se répandre entre les femelles
9 dans une population si ces traits ne leur offrent aucun avantage. Pour répondre à ce dilemme, trois hypothèses ont été retenues en suggérant un lien entre les gènes de préférence pour les femelles et le trait préféré du mâle : l'hypothèse de Fisher (1930), l'hypothèse du handicap (Zahavi 1975) et l'hypothèse des bons gènes (Hamilton et Zuk 1982).
L'emballement de Fisher (1930) suggère que le trait choisi de la femelle n'est pas arbitraire au départ et que sa variation influence la valeur adaptative des deux sexes. Cela dit, la fréquence de femelles qui auront cette préférence devrait croître au sein de la population jusqu'à ce que l’expression du trait pour le mâle lui apporte un avantage supplémentaire pour accéder aux femelles. Comme il y a une base génétique pour la préférence et le trait (Fisher 1930), les individus de la future génération porteront ces gènes et ces derniers seront exprimés dans chacun des sexes : les femelles devraient exprimer la préférence et les mâles devaient arborer le trait (Andersson et Simmons 2006, Lande 1981, Mead et Arnold 2004). L'accouplement d’un mâle arborant un trait et d'une femelle avec une préférence sexuelle pour ce trait augmente la proportion d'individus qui exprimeront le trait et qui auront une préférence. Une boucle de rétroaction positive peut ensuite apparaître, ce qui accentue l'intensité du trait ainsi que la préférence, appelé le processus d’emballement de Fisher (1930). L'intensité augmentera jusqu'à ce que des pressions de sélection opposées s'accentuent et ralentissent ou mettent fin à la boucle (Andersson 1982).
L'hypothèse du handicap suggère que les caractères exagérés des mâles agissent comme des signaux honnêtes sur la qualité potentielle des partenaires et que le seul avantage pour les femelles est de léguer cette qualité à leur progéniture (Zahavi 1975). En effet, l’expression du trait agirait comme un handicap pour le mâle puisque les coûts énergétiques de la production et du maintien du trait confèrent un désavantage à sa survie. Cependant, l’expression du trait suggère que l'individu est en mesure de survivre malgré son handicap et que des gènes utiles à la survie sont présents. L’expression du trait est donc un signal honnête de qualité génétique et non simplement un trait facilitant l'accès aux partenaires.
10 Finalement, l'hypothèse des bons gènes suggère que la femelle choisira un mâle en fonction de certains traits qui montrent une bonne qualité génétique (Hamilton et Zuk 1982). Par exemple, la masse corporelle, les couleurs flamboyantes du plumage ou la longueur des cornes peuvent être sélectionnées sexuellement (Coltman et al. 2002, Møller et Cuervo 1998). Bien que certains traits soient choisis pour des avantages directs à la femelle, comme une protection contre les autres mâles ou un apport garanti en nourriture (Dahlgren 1990), certains ne l'avantagent pas directement. Cependant, ces traits peuvent toujours avoir un impact positif indirect. En effet, les jeunes du sexe sous sélection porteront ces gènes et auront eux aussi une meilleure probabilité de se reproduire, ce qui augmente indirectement l'aptitude phénotypique de la mère (Wilkinson et Reillo 1994).
1.3 Principe de Bateman et l'investissement parental
Bateman (1948) a suggéré que le succès reproducteur des femelles drosophiles (Drosophila melanogaster) est limité par leur accès à la nourriture, alors que celui des mâles est limité par leur accès aux partenaires. Ce phénomène, le principe de Bateman, suggère alors que le succès reproducteur des mâles est limité par la compétition entre mâles, ce qui inflige une pression de sélection entre ces derniers, une sélection intrasexuelle. Selon Bateman (1948), la dichotomie en facteurs limitants favorise un système d’accouplement de promiscuité pour les mâles, mais pas pour les femelles. Il suggère également que la variance du nombre de partenaires des mâles est une cause majeure de la différence entre les sexes pour la variance de fitness. Trente ans plus tard, Trivers (1972) a reformulé de façon plus générale le principe de Bateman pour qu'il soit applicable à des niveaux plus larges. Ce dernier a proposé le concept d'investissement parental, définit comme étant toute allocation d'un parent envers un jeune qui augmente sa probabilité de survie et de se reproduire, mais qui a comme coût de réduire son succès reproducteur futur. L'investissement dans la reproduction ne tient pas seulement compte de la production de gamètes, mais également des autres allocations nécessaires à la production et croissance des rejetons jusqu'à maturité. Les soins parentaux en sont un exemple. Ainsi, le sexe qui investira
11 davantage dans la production d'un jeune sera une ressource limitante pour celui qui investira le moins, et ce dernier sera également soumis à la sélection sexuelle (Trivers 1972).
Chez les mammifères, les formes que prennent l'investissement parental sont différentes entre les sexes. Chez les femelles, l’investissement parental se manifeste par la gestation, la lactation et les soins post-nataux aux jeunes pour assurer leur survie (Gubernick et al. 2013). Cependant, les mâles sont soumis à un compromis entre l’investissement parental pour leurs jeunes et le temps passé à trouver des opportunités d’accouplement supplémentaires (Trivers 1972). Des soins paternels sont communément observés chez les mammifères monogames puisque l’allocation dans la survie des jeunes jusqu’à maturité sexuelle augmente l’aptitude phénotypique du père (Woodroffe et Vincent 1994). Cependant, chez les mammifères polygynes, les mâles allouent davantage de ressources sur l’effort d’accouplement que sur les soins paternels, bien que ces derniers puissent être présents à des niveaux plus faibles (Hoffman et al. 2008, Woodroffe et Vincent 1994). Le degré d’allocation dépend également de la qualité de la mère, du degré de certitude de paternité ou de la taille du groupe, où l’allocation du père diminue avec l’investissement alloparental qui est favorisé dans les gros groupes (Schubert et al. 2009). D’ailleurs, Adrian et al. (2005) ont étudié le degré de soins paternels apportés par trois différentes espèces de cochons d’Inde qui ont des systèmes d’accouplement différents : le cobaye du Brésil (Cavia aperea) est polygyne, le cobaye-belette (Galea musteloides) montre de la promiscuité (chaque sexe s’accouple avec plusieurs partenaires) et le cobaye à dents jaune de Muenster (Galea monasteriensis) est monogame. Ils ont trouvé que les mâles polygynes et monogames apportaient des soins paternels par des comportements de jeu et de toilettage, mais ces comportements étaient absents chez les mâles pratiquant la promiscuité, puisque le degré de certitude de paternité était plus faible.
De manière générale, lorsqu'un sexe est soumis plus fortement à la sélection sexuelle, un dimorphisme sexuel est sujet à évoluer. Ces espèces ont généralement un système d’accouplement polygyne (Promislow 1992). Puisque le succès reproducteur du mâle chez la majorité des mammifères est limité par le nombre de partenaires avec qui il peut s'accoupler
12 (Kokko et Monaghan 2001, Trivers 1972), la compétition intrasexuelle peut potentiellement créer une variance importante dans le succès reproducteur des mâles (Emlen et Oring 1977). Si les variations héritables du phénotype sont corrélées avec le succès d'accouplement, une sélection sexuelle pour certains traits peut s'établir (Coltman et al. 2002). Dans cette situation, les mâles ayant un phénotype qui lui confère un avantage dans la compétition intrasexuelle ou qui peut être choisi par les femelles auront un meilleur succès reproducteur. Cette hypothèse sur l'origine du dimorphisme sexuel suggère que les traits avantageux, ou leur niveau d'expression, devraient augmenter en proportion dans une population au fil des générations. Lorsque la majorité des individus d’une population arborent ces traits, les individus qui les expriment de manière encore plus accentuée seront alors favorisés, ce qui accentuera à son tour le dimorphisme (Andersson 1994).
1.4 Stratégies de reproduction et tactiques de reproduction alternatives
Une stratégie de reproduction se définit comme étant un ensemble de décisions et de règles sous une base génétique sur l’allocation des ressources entre le maintien de la condition corporelle et l’effort de reproduction (Gross 1996). Différents phénotypes peuvent être observée à partir d’une même stratégie de reproduction, ce qu’on appelle une tactique de reproduction (Gross 1996). Par exemple, chez plusieurs espèces avec un système de reproduction polygyne, la compétition intrasexuelle incite les mâles à combattre ou à se faufiler pour accéder aux femelles. L’ensemble de décisions qui mène un individu à combattre sera la stratégie de reproduction, dirigé par mécanismes développementaux, physiologiques et neurologiques. Les tactiques, dans cet exemple, seraient le combat (la tactique principale) et le faufilage (une tactique alternative) qui seront tous deux associés au phénotype de l’individu, comme son comportement ou sa morphologie (Gross 1996).
Ce type de tactiques alternatives est sujet à apparaître dans une population lorsque la compétition intrasexuelle mène à une variance importante dans le succès reproducteur des mâles (Emlen et Oring 1977). Quelques mâles dominants auront la majorité des paternités de la population, alors
13 que la majorité des subordonnés ne se reproduiront pas (Emlen et Oring 1977). Un exemple serait celui du babouin hamadryas (Papio hamadryas), où un dimorphisme sexuel s’observe par la présence de crocs et de fourrure autour du cou des mâles (Danish et Palombit 2014). Des combats entre mâles déterminent le rang social de chacun et quelques les dominants auront accès à un harem complet de femelles avec qui s’accoupler, alors que les subordonnés auront peu ou pas de partenaires (Danish et Palombit 2014). Cependant, chez une majorité d’espèces, les mâles adoptent des comportements non-compétitifs alternatifs, ce qui peut maintenir une certaine variation génétique au sein de la population (Dominey 1984).
1.5 Investissement dans la reproduction
Comme mentionné ci-haut, les mâles chez les mammifères polygynes sont soumis à une compétition intrasexuelle pour accéder aux femelles (Andersson 1994). L'investissement dans la reproduction peut alors s'observer par l’expression et le maintien de traits phénotypiques avantageux pour la compétition. Cette compétition est connue pour être coûteuse dû au combats intrasexuels (Møller et Cuervo 1998) ou aux coûts reliés aux traits eux-mêmes, comme les coûts reliés à la production, au transport ou au maintien du trait (Møller 1996). Par exemple, Yoccoz et al. (2002) ont suggéré que, chez le cerf élaphe (Cervus elaphus), il y a une perte en masse chez les mâles durant le rut. Cette perte était d'ailleurs maximale chez les mâles d'âge moyen, les plus impliqués dans les combats et la reproduction, suggérant qu’il y a un coût à participer au rut. Toutefois, Pelletier et al. (2006) ont montré que la longévité des béliers était positivement corrélée avec l'effort de reproduction entre deux et cinq ans, ce qui suggère que certains mâles ont la capacité d'acquérir plus de ressources et mieux performer que les autres (van Noordwijk et de Jong 1986).
Comme l'allocation à la reproduction peut réduire l’énergie disponible pour la croissance, l'âge auquel un individu itéropare commence à se reproduire peut avoir des conséquences majeures sur sa survie (Swain et al. 2007). Les individus capables d'acquérir plus de ressources devraient donc participer plus tôt à la reproduction (Descamps et al. 2006). La probabilité de mourir avant
14 de s'être reproduit augmente également avec chaque événement de reproduction manqué, ce qui incite les mâles à prendre des risques tôt dans la vie (Blomquist 2009). L'âge auquel un individu commence à se reproduire peut donc influencer son histoire de vie et son aptitude phénotypique (Markussen et al. 2019).
1.6 Système d'étude
1.6.1. Le mouflon d'Amérique
Le mouflon d'Amérique est un grand mammifère herbivore habitant les montagnes de l’Ouest de l’Amérique du Nord. Chez cet ongulé, le dimorphisme sexuel est très prononcé. Les béliers sont en moyenne 65% plus lourds que les brebis à l'âge de six ans (Festa-Bianchet et al. 1996). De plus, les mâles sont munis de cornes massives en guise de trait sexuel secondaire, alors que celles des femelles sont beaucoup plus petites. Ces cornes, pouvant atteindre 15% de la masse corporelle d’un mâle, sont utilisées comme armement pour déterminer le rang social de chaque individu dans la population (Geist 1971, Pelletier et Festa-Bianchet 2006). Cette hiérarchie est directement liée à l'accès aux femelles, donc au succès reproducteur du mâle (Pelletier et al. 2006, Pelletier et Festa-Bianchet 2006). Les béliers prennent une part active dans le rut à partir de l'âge de deux ans (Pelletier et Festa-Bianchet 2006). Cependant, les jeunes compétiteurs subordonnés ne peuvent pas faire de tending (voir définition plus bas) dû à leur faible masse corporelle, ce qui limite leur accès aux partenaires sexuelles (Pelletier et al. 2006, Pelletier et Festa-Bianchet 2006). Effectivement, la masse asymptotique n’est pas atteinte avant l’âge de sept ans (Festa-Bianchet et al. 1996), ce qui crée un grand écart en masse entre les jeunes et les compétiteurs plus âgés.
Chez les grands mammifères, le succès reproducteur des mâles est généralement fortement corrélé avec la masse corporelle, car elle améliore l'aptitude d'un individu à combattre les concurrents du même sexe et, ainsi, favoriser son accès aux partenaires (Coltman et al. 2002). Ce phénomène est observé chez les ongulés comme le mouflon d'Amérique, et est accompagné
15 d'un système d'accouplement polygyne qui bénéficie les mâles les plus lourds et avec les plus grandes cornes (Coltman et al. 2002). Les combats ont lieu dès octobre lorsque les béliers se regroupent et ils renforcent ainsi leur statut social avant le début du rut fin novembre (Pelletier et Festa-Bianchet 2006).
Chez les mouflons d'Amérique des Rocheuses canadiennes, les réserves énergétiques corporelles sont accumulées durant l'été et s'épuisent durant l'hiver (Douhard et al. 2018). De plus, chez les ongulés en général, la prise alimentaire diminue durant le rut (Miquelle 1990, Pelletier et al. 2009). Cette contrainte apporte un choix d'allocation du bélier durant le rut puisque la perte en masse (Yoccoz et al. 2002) et les blessures lors des combats pourraient potentiellement être cruciales à sa survie (Hogg et Forbes 1997).
1.6.2. Tactiques de reproduction du mouflon d’Amérique
Hogg (1984) a montré que les béliers du mouflon d’Amérique utilisaient trois tactiques de reproduction, soit le tending, le coursing et le blocking. Les mâles de deux à quatre ans utilisent presqu'exclusivement le coursing (Pelletier et Festa-Bianchet 2006). Voici la définition des tactiques selon Hogg (1984) :
Tending : Tactique utilisée seulement par les mâles ayant les plus hauts rangs de dominance sociale de la population, donc ceux avec une masse corporelle élevée et des cornes très développées. Ces béliers défendront chacun une brebis en œstrus.
Coursing : Cette tactique consiste à accéder aux femelles en déjouant le mâle défenseur. Les mâles pratiquant cette tactique sont situés plus bas dans le rang social de la population.
Blocking : Ces béliers subordonnés vont tenter d’isoler de force la brebis loin de la région utilisée par le mâle dominant et attendre qu'elle devienne réceptive pour copuler. Cette tactique est observée moins fréquemment.
16 Parmi ces tactiques, le tending est de loin celle qui apporte le meilleur succès reproducteur (Hogg et Forbes 1997, Pelletier et Festa-Bianchet 2006). Effectivement, Hogg et Forbes (1997) ont montré que les quelques mâles qui peuvent faire du tending étaient pères de plus de la moitié des agneaux de la population. Lors du rut, les femelles tombent en œstrus à différents moments. Bien que le nombre doive varier selon la taille de la population, des observations sur le terrain suggèrent qu’il y a rarement plus de trois femelles en œstrus à la fois (Hogg 1984). S’il y a seulement une femelle en œstrus à une journée donnée, le mâle au sommet du rang social aura priorité s’il réussit à la trouver. Le même principe s’applique pour le mâle au deuxième rang s’il y a deux femelles en œstrus dans cette même journée (Hogg 1984).
1.6.3. Chasse aux trophées
La chasse aux béliers est une activité bien appréciée et pratiquée des chasseurs de trophées, qui visent un trait phénotypique particulier, ici les cornes, plutôt que la viande (Brewer et al. 2014, Coltman et al. 2003). Au Canada, les règlements de chasse sont de compétence provinciale. En Alberta, pour qu'un bélier soit récolté, le chasseur doit avoir un permis et l'individu chassé doit respecter certains critères de légalité qui dépendent de la courbure des cornes (Pigeon et al. 2016). La plupart des régions oblige une courbure d'au moins 4/5e, bien que certaines demandent
une courbure dite full curl (Figure 1.2). De manière générale, ce seuil n'est pas atteint avant l'âge d’au moins 4 ans (Jorgenson et al. 1993). Chez les mouflons, le succès reproducteur des mâles adultes est fortement corrélé avec la taille des cornes, puisqu'elle favorise un haut rang social et donc la possibilité de pratiquer du tending (Coltman et al. 2002, Pelletier et Festa-Bianchet 2006).
17 Figure 1.2. Critères de légalité pour la récolte de mouflons d'Amérique en Alberta.
La figure A. illustre la taille minimale selon un critère 4/5e et B. selon un critère
full curl. Figure tirée de Pigeon et al. (2016).
1.6.4. Population de Ram Mountain
La prise de données à Ram Mountain en Alberta, Canada (52oN 115oO), a débuté en 1971 et la
collecte de données sur le succès reproducteur des mâles a commencé à partir de 1988. La population étudiée utilise environ 38 km2 et exploite tous les niveaux d'altitude de la montagne,
de 1080 m jusqu'à 2170 m du niveau de la mer (Jorgenson et al. 1993). La montagne est entourée par une forêt de conifères sur trois côtés et par la Rivière Saskatchewan Nord sur le quatrième côté (Coltman et al. 2002).
La période de suivi annuelle se fait de la fin mai jusqu'à la fin septembre par la capture et l’observation des individus. Les mouflons sont capturés dans une trappe de type corral avec du sel au centre pour les y attirer. Plusieurs mesures et prises d'échantillons sont effectuées à chaque
18 capture. Les mouflons sont marqués à l'aide d'étiquettes Allflex en plastique colorées et numérotées installées sur les oreilles. Les femelles ont également un collier visuel. Lors des captures, les mouflons sont pesés avec une balance à ressort Dectecto (Brooklyn, N.Y.) ayant une marge d'erreur de 250g (Festa-Bianchet et al. 1998). Les observations de terrain et les multiples captures annuelles permettent de suivre la taille de la population, la structure d'âge, les changements de masse, la croissance des cornes et les accouplements à l'intérieur de la population. Le taux annuel de recapture est de plus de 95% pour les femelles et d’environ 85% pour les mâles (Jorgenson et al. 1993), alors que leur taux de détection interannuel est d'environ 99% et 95% respectivement (Bonenfant et al. 2009)
Les captures annuelles et les échantillons d’ADN permettent la construction d'un pedigree de la population. De 1971 à 1988, la maternité est connue pour 709 des 894 mouflons marqués. Celle-ci était confirmée par des observations d’allaitement sur le terrain. À partir de 1988, les analyses d'identification par ADN ont permis d’assigner également la paternité des agneaux (Coltman et al. 2005). Jusqu'en 1993, l'analyse se faisait par échantillons de sang conservés dans un préservatif à -20oC. En 1994-1996, aucun échantillon d’ADN n’a été récolté. En 1997, la prise
d'échantillons s’est poursuivie avec des poils. Entre 50 et 100 poils étaient récoltés par individu et gardés dans une enveloppe de papier ou de plastique contenant environ 5 g de silice à température ambiante. Une ponction circulaire de 8 mm de diamètre de tissu d'oreille était prélevée de 1998 à 2002, de 5 mm de 2003 à 2011 (Coltman et al. 2005) et de 2 mm de 2012 à aujourd’hui. Ce dernier est conservé à -20oC dans l’éthanol (95%). Bien que le tissu employé
varie selon l'année d'échantillonnage, l'extraction d'ADN se fait en utilisant la trousse d'extraction QIAamp (Qiagen Inc., Mississauga, Ontario) (Coltman et al. 2003, Poissant et al. 2013). La paternité est assignée à l'aide du logiciel CERVUS 3.0 par une approche statistique de vraisemblance avec un intervalle de confiance à 95% (Coltman et al. 2002). Sur les 770 agneaux vus entre 1988-2018, un échantillon d'ADN a été obtenu pour 721 et 380 d’entre eux ont été assignés à un père connu. Il est important de noter que le succès reproducteur ne peut être déterminé que pour les individus qui ont été capturés, génotypés et assignés à un père connu. Un retour sur ce point sera fait au chapitre 3 dans les limitations de l’étude.
19 La masse corporelle est ajustée au 15 septembre pour chaque individu à l'aide d'un modèle de croissance linéaire avec la racine carrée de la date de capture pour les mâles capturés plus d'une fois entre le 25 mai et le 15 septembre (Martin et Pelletier 2011). Les individus non capturés une année ou capturés plus de 50 jours avant le 15 septembre sont exclus de l’analyse. Depuis mai 1988, 157 mâles adultes ont été échantillonnés. De ce nombre, 136 ont une masse corporelle connue, 76 se sont reproduits et 16 sont morts à la chasse. En retirant les individus de masse inconnue ou morts à la chasse, 51 reproducteurs demeurent à l’étude.
CHAPITRE 2
DÉTERMINANTS ET COÛTS À LONG TERME D'UNE REPRODUCTION EN JEUNE ÂGE CHEZ UN MÂLE D'UN MAMMIFÈRE POLYGYNE ET LONGÉVIF
2.1 Description de l’article et contribution
Il est crucial d’explorer et quantifier l’importance des déterminants du succès reproducteur des mâles afin de mieux comprendre le rôle de la sélection sexuelle sur l’évolution des espèces. Chez les espèces polygynes où la majorité des paternités proviennent de quelques individus dominants, des tactiques de reproductions alternatives assurent une certaine proportion de paternités provenant des jeunes mâles subordonnés, ce qui pourrait augmenter la variabilité génétique au sein d’une population. Cependant, peu d’informations sont connues à ce jour sur les déterminants du succès reproducteur des mâles en jeune âge. L’objectif de cet article est d’identifier ces déterminants au niveau phénotypique et démographique, ainsi que de quantifier les conséquences à long terme de la reproduction en jeune âge sur la longévité et le succès reproducteur à vie. Les résultats montrent qu’une augmentation de la masse corporelle, de la proportion de jeunes et de la proportion de femelles dans la population favorisent la probabilité de se reproduire en jeune âge, bien que ces effets soient significatifs seulement pour les mâles de 3 ans. Nous montrons également qu’une reproduction en jeune âge tend à augmenter le succès reproducteur à vie, mais au coût de mourir plus tôt. Fanie Pelletier, Marco Festa-Bianchet et moi avons développé les idées. J’ai fait les analyses statistiques, la rédaction initiale du manuscrit et participé à la collecte de données à Ram Mountain pour deux saisons de terrain. Fanie, Marco, David Coltman et moi avons discuté des résultats finaux pour tisser des liens biologiques et avons contribué à la rédaction finale du manuscrit.
22 DETERMINANTS OF EARLY REPRODUCTION IN MALES OF A LONG-LIVED
POLYGYNOUS MAMMAL
Sous presse à Ecology and Evolution
23 2.2 Abstract
In long-lived polygynous species, male reproductive success is often monopolized by a few mature dominant individuals. Young males are too small to be dominant and may employ alternative tactics. However, little is known about the determinants of reproductive success for young males using alternative mating tactics. Understanding the causes and consequences of variability in early reproductive success may be crucial to assess the strength of sexual selection and possible long-term trade-offs among life-history traits. We evaluated the determinants of reproduction in male bighorn sheep (Ovis canadensis) aged 2-4 years using 30 years of individual-level data. These young males cannot defend estrous ewes and must use alternative mating tactics. We also investigated how the age of first detected reproduction affected lifetime reproductive success and longevity. We found that reproductive success of males aged three years was positively affected by body mass, by the proportion of males aged 2-4 years in the competitor pool and by the number of females available per adult male. These results suggest that reproductive success likely depends on both population sex-age structure and competitive ability. None of these variables, however, had significant effects on the reproductive success of males aged 2 or 4 years. Reproduction before the age of five increased lifetime reproductive success but decreased longevity, suggesting a long-term survival cost of early reproduction. Our analyses reveal that both individual-level phenotypic and population-level demographic variables influence reproductive success by young males and provide a rare assessment of fitness offs in wild polygynous males, thus increasing knowledge on life-history trade-offs.
24 2.3 Significance Statement
Trade-offs among fitness components, such as between reproduction and survival, affect life-history evolution and population dynamics. Trade-offs have been well studied in females, but little is known about trade-offs in male mammals, particularly in long-lived polygynous species. Reproductive costs in polygynous males are difficult to measure because allocation to male-male competition does not necessarily lead to reproductive success, and parentage assignment requires genetic analyses. This study’s finding provides knowledge on life-history trade-offs in polygynous mammals by suggesting that early reproductive success is weakly affected by physical and demographic characteristic in male bighorn sheep. It also shows a long-term advantage of early allocation to reproduction but a cost in survival.
25 2.4 Introduction
Life history theory predicts diverse reproductive strategies among species and between individuals to maximise fitness (Stearns 1992). Individuals also vary in their capacity to acquire energy and, since resources are limited in a natural environment, allocation trade-offs between fitness components are expected (Hamel et al. 2010, van Noordwijk and de Jong 1986). Allocation refers to the assimilated energy that is allocated to reproduction, growth or survival (Perrin and Sibly 1993, van Noordwijk and de Jong 1986). Trade-offs between growth or survival and reproduction have been found in several species (Cox et al. 2010, Folkvord et al. 2014). For example, female red squirrels (Tamiascirus hudsonicus) with greater resource acquisition capacity started to reproduce earlier at the expense of decreased longevity, while females with fewer resources delayed maturity and had greater longevity (Descamps et al. 2006). Blomquist (2009) pointed out that the risk of dying before reproducing increases with each reproductive opportunity missed. Thus, variation in age at first reproduction may substantially influence fitness and has been studied in birds (Cooper et al. 2009, Aubry et al. 2009), terrestrial mammals (Martin and Festa-Bianchet 2012, Neuhaus et al. 2004, Markussen et al. 2019), marine mammals (Hadley et al. 2006, Lloyd et al. 2020), fish (Swain et al. 2007), and reptiles (Bonnet et al. 2002). However, in wild vertebrates, most studies testing early and late-life trade-offs in just one sex were conducted on females (Lemaître et al. 2015). Trade-offs affecting females may not be relevant for males in polygynous species, because the drivers and consequences of reproductive effort are substantially different between sexes. Male fitness is limited by fertilization opportunities, female fitness is mostly limited by forage resources, and males can produce many more offspring than females (Bateman 1948, Clutton-Brock 1988, Emlen and Oring 1977, Trivers 1972). Little information is available on reproductive costs for polygynous and promiscuous males, partly because allocation to competition with other males is not directly linked to actual reproductive success. A male may expend much effort competing with other males but not obtain any matings (Festa-Bianchet 2012, Hamel et al. 2010, Lemaître et al. 2020, Lloyd et al. 2020, Pelletier et al. 2006). In addition, parentage assignment is challenging in wild populations because it requires genetic material from juveniles and most putative fathers (Coltman et al. 2005).
26 In iteroparous species, different allocations of limited energy among growth, reproduction and survival may maximise fitness (Stearns 1992). Excessive allocation to reproduction during one reproductive event may decrease survival (Chase 1999) or compromise future reproduction (Nilsson and Svenssonn 1996). On the other hand, an individual that allocates most available energy to survival may not succeed at the next reproductive event if mortality is high or the level of competition increases (Bell 1980, Chase 1999, Wittenberger 1979).
When ability to acquire resources differs among individuals, the consequences of allocation to reproduction may also vary (Hamel et al. 2010, van Noordwijk and de Jong 1986). Differences in lifetime reproductive success are likely to be particularly important in polygynous systems, where a few highly competitive males can monopolize reproduction over one or a few breeding seasons (Andersson 1994). In polygynous species, male reproductive success is generally associated with dominance, which is often determined by body mass and size of secondary sexual traits (Bergeron et al. 2010, Lloyd et al. 2020, Martin et al. 2013, Pelletier and Festa-Bianchet 2006). When reproductive success is monopolized by a few dominant males, most males do not reproduce (Coltman et al. 2002). Thus, different reproductive strategies may develop, including differences in allocation to growth or the age at which individuals first attempt to reproduce (Pianka and Parker 1975).
In some species, young subordinate males adopt alternative mating tactics (Hogg 1984, Pelletier et al. 2006, Willisch et al. 2012). Allocation to reproduction at early ages could have substantial consequences on male lifetime reproductive success (Bergeron et al. 2010). Those consequences could be positive if early attempts to reproduce increase experience and therefore subsequent reproductive success, or negative if competitive interactions involve a risk of injury or substantial energy costs (Bergeron et al. 2010, Weladji et al. 2008). Longevity can also be influenced by early reproductive success if early allocation to reproduction has long-term costs (Descamps et al. 2006). For example, male ungulates that participate actively in the rut risk injuries and lose body mass, which can decrease survival probabilities year after year, thus decreasing longevity (Bergeron et al. 2010, Yoccoz et al. 2002). Because only a few studies
27 have the necessary long-term data on individual males, the causes and consequences of early reproductive success for males of polygynous species are mostly unknown. The main objective of this study is to identify the factors influencing early male reproductive success in a polygynous species, the bighorn sheep (Ovis canadensis), and evaluate the consequences of early reproduction on longevity and lifetime reproductive success.
Reproductive success in male bighorn sheep is strongly associated with social rank, which tends to increase with age and mass (Pelletier and Festa-Bianchet 2006). Agonistic interactions in the two months preceding the rut, from late September to early November, are used to establish rank (Pelletier and Festa-Bianchet 2006). Rutting activities involve a risk of injury (Festa-Bianchet 1987). During the rut, males typically use either a tending or a coursing tactic (Hogg 1984, Hogg 1987, Hogg and Forbes 1997). Tending is used exclusively by dominant males and consists of defending a single estrous ewe against competitors. The alternative tactic, coursing, is used by subordinates and involves attempts to separate the tending male from the ewe and force a copulation before the tending male regains control (Hogg 1984, Hogg 1987). Although the tending tactic is most efficient, about 40% of paternities are obtained by coursing males (Hogg and Forbes 1997).
Reproductive success in subordinate males is influenced by secondary sexual traits, such as body mass, and sexual selection increases with the number of competitors (Martin et al. 2016). Phenotypic and demographic parameters likely affect early reproductive success. Body mass is a major determinant of male reproductive success in many polygynous mammals including bighorn sheep (Pelletier and Festa-Bianchet 2006), but it is unclear if body mass is important for young males, that are always too small to adopt the tending tactic. Age structure, sex ratio and population density are expected to affect reproductive success in ungulate males (Komers et al. 1997, Markussen et al. 2019, Newbolt et al. 2017) because reproductive success depends upon the ability to overcome competitors (Festa-Bianchet 2012) and the level of competition may decrease when more breeding females are available (Clutton-Brock et al. 1997).
28 We investigated how body mass, age structure, sex ratio and population density affected reproductive success for males aged two to four years. We chose this age group because these males are expected to use exclusively the coursing tactic, based on rut observations at Sheep River, Alberta (Pelletier et al. 2006). We predicted that body mass should increase reproductive success at all ages, because larger males should be better able to outcompete rivals, or to sustain greater effort in coursing competition. We examined the effects of age structure under the expectation that a high proportion of young males within the competitive pool may make it harder for tending males to defend estrous ewes, and young males should obtain a greater share of paternities (Bonenfant et al. 2004). A breeder sex ratio skewed toward males should increase competition between males due to a lower ratio of estrous ewes in the population, and young males may be completely excluded from reproduction (Bonenfant et al. 2004). High population density may decrease reproductive success of young adult males due to an increase in pre-rut competition for resources, potentially leading to poorer condition when starting the rut (Yoccoz et al. 2002). However, the effect of population density on reproductive success is highly variable in the literature (Komers et al. 1997, Mysterud et al. 2003, 2004, Yoccoz et al. 2002). An alternative prediction would be a positive relationship with reproductive success due to an increase in the number of available estrous females (Komers et al. 1997).
We then investigated how early reproduction affects long-term fitness by analyzing its effects on longevity and lifetime reproductive success. If early reproduction was only possible for males that had acquired substantial resources, then early reproductive success should have a positive relationship with longevity and lifetime reproductive success. If early reproductive success led to a substantial drain on body resources, however, it should have a negative relationship with longevity and lifetime reproductive success (Metcalfe and Monaghan 2003).
29 2.5 Materials and Methods
2.5.1. Study area and population
Bighorn sheep have been monitored since 1971 on Ram Mountain (52oN, 115oW), Alberta,
Canada. The study area is approximately 38 km2 and the sheep population is mostly isolated by
coniferous forests, which surround the mountain except on the North-West side where the North Saskatchewan River separates Ram Mountain from Shunda Mountain, which harbours another small population of bighorn sheep. From late May to late September, sheep were captured in a corral trap baited with salt. Most adults were captured two to five times each summer. At each capture body mass was measured to the nearest 250 g using a Detecto spring scale (Brooklin, NY). Repeated measurements from each individual each year allowed us to adjust mass to September 15 using individual growth curves. Linear mixed models with a restricted maximum likelihood were used to adjust mass fitted as a function of date with May 25 as day 1 (Martin and Pelletier 2011). Adjusted mass was not estimated for individuals not captured within 50 days of September 15. Males were marked using unique combinations of colored and numbered Allflex ear tags. Lambs were marked with numbered Ketchum metal tags (Ketchum Manufacturing, Ottawa, Ontario) and colored strips of Safeflag plastics (Pawtucket, R.I.), which were replaced by Allflex tags at one year of age. The probability of detection for surviving sheep is more than 95% for males and 99% for females (Bonenfant et al. 2009).
Analyses included every male aged at least two years during the ruts 1987-2017. Our sample thus began with rams aged at least three years in May 1988, when DNA sampling was initiated (Coltman et al. 2002). A male aged three in 1988 would have participated in the 1987 rut as a 2-year-old. We analyzed males aged at least two years during the rut because no paternity has ever been assigned to male lambs or yearlings. Of 157 males captured between 1988 and 2018, 136 had known annual reproductive success and body mass adjusted to September 15. Of those 136 males, 76 sired at least one lamb throughout their lives. Hunters could harvest males with horns describing at least four-fifths of a curl until 1995, and only full-curl individuals from 1996