IL-7 receptor blockade blunts antigen-specific memory T cell responses and chronic inflammation in primates
Texte intégral
Figure
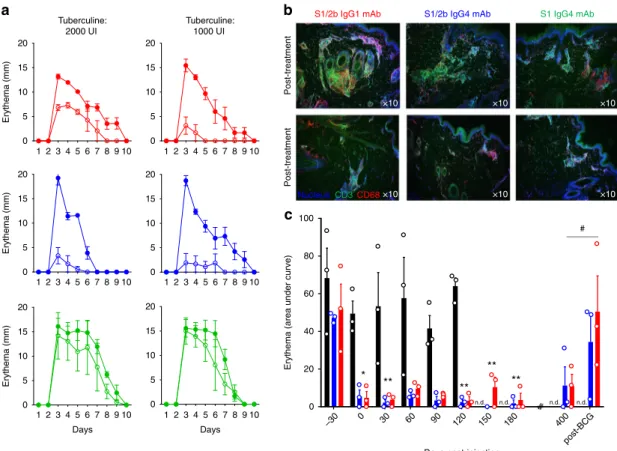
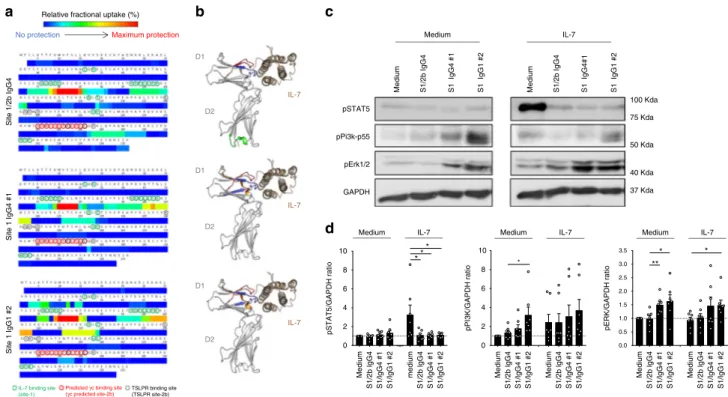
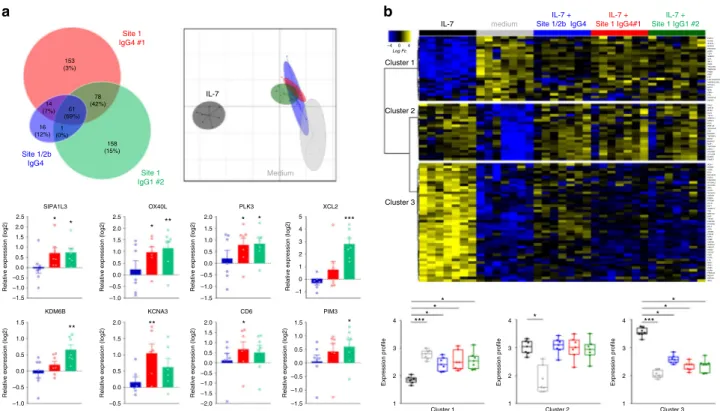
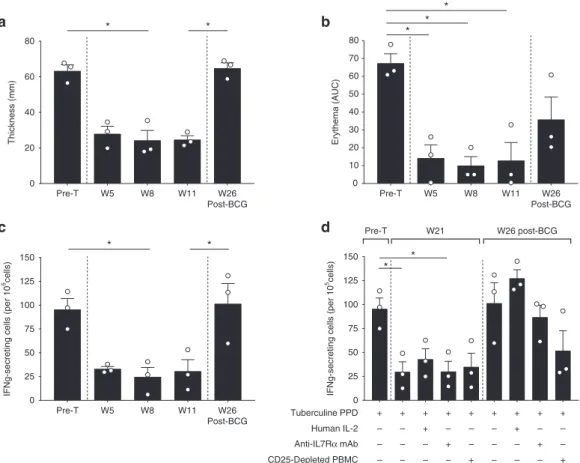
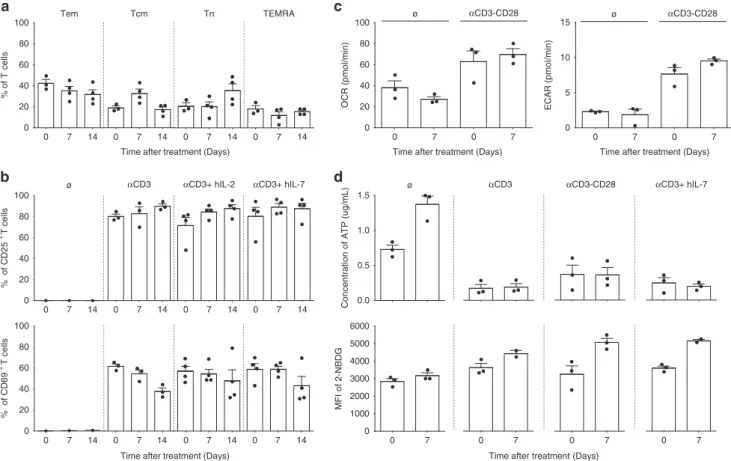
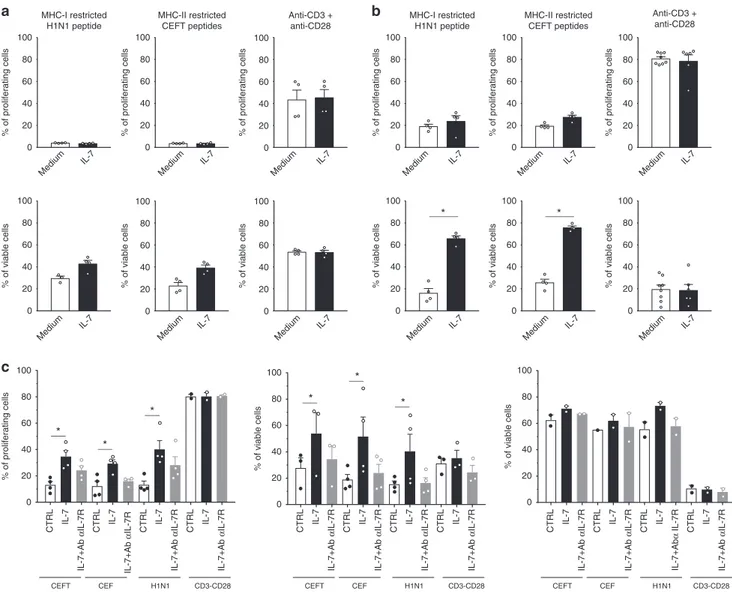
Documents relatifs
Figure 5: Evolution of the main metabolic molecules and uptake rates of glucose and oxygen as a function of the cell energetic demand from low to high. Oxygen
CS regulated inflammation induced in vivo in mice through inhibition of cell infiltration and cytokine gene expression In order to better understand the mechanisms under- lying
Interestingly, both studies revealed that although these different microglia subtypes up-regulated several classes of genes associated with myeloid cell activation and
This paper aims to examine the ways in which the British contemporary writer Gabriel Josipovici raises question relating to memory, reminiscence, and commemoration in
The aim of the present study was to investigate the effect of HER1/2/3 blockade through a combination of cetuximab and pertuzumab, with or without photon irradiation, on
subtilis spores could act as potential vaccine adjuvants that synergistically pro- vide antigen-specific immune responses against the avian influenza virus H9N2 in White
may thus compound the precise analysis of ROQUIN level of expression in tumor T FH , we thus performed additional tran- scriptomic analyses of CD4 + /CXCR5 + /ICOS + sorted T FH
The in vitro and in vivo anti-cancer potential of Mycobacterium cell wall fraction (MCWF) against canine transitional cell carcinoma of the urinary bladder... , Belleville ON K8N
