Universite de Sherbrooke
Sk
^^
31156008108873
<y
ETUDE DE LA VARIATION INTERSPECIFIQUE DE LA
VITESSE SPECIFIQUE DE CROISSANCE ET
MODELISATION DE VEFFET DES ATTRIBUTS
MORPHOLOGIQUES, PHYSIOLOGIQUES ET
D'ALLOCATION DE BIOMASSE
'nr
\Ad.j par Driss Mezianethese presentee au Departement de biologie
en vue de 1'obtention du grade de docteur es sciences (Ph.D.)
FACULTE DES SCIENCES
UNIVERSITE DE SHERBROOKE
1*1
National Library of Canada Acquisitions and Biblioqraphic Services 395 Wellington Street Ottawa ON K1AON4 Canada Bibliotheque nationale du Canada Acquisitions et services bibliographiques 395, rue Wellington Ottawa ON K1AON4 CanadaYour file Votre reference
Our file Notre reference
The author has granted a
non-exclusive licence allowing the
National Library of Canada to
reproduce, loan, distribute or sell
copies of this thesis in microform,
paper or electronic formats.
The author retains ownership of the
copyright m this thesis. Neither the
thesis nor substantial extracts from it
may be printed or otherwise
reproduced without the author's
permission.
L'auteur a accorde une licence non
exclusive pennettaat a la
Bibliofheque nationale du Canada de
reproduire, prefer, distribuer ou
vendre des copies de cette these sous
la forme de microfiche/fihn, de
reproduction sur papier ou sur fonnat
electronique.
L'auteur conserve la propriete du
droit d'auteur qui protege cette these.
Ni la these ni des extraits substantiels
de celle-ci ne doivent etre imprimes
ou autrement reproduits sans son
autonsation.
Le /o/.^/rhW^ /7/t ,/le jury suivant a accepte cette tt^ese dans sa version finale. date
President-rapporteur: Mme Colette Ansseau
Departement de biologie
Membre: M. Bill Shipley
Departement de biologie Membre: M. Gilles Grenier
Departement de biologie
Membre: M. Robert van Hulst
Universite Bishop
Membre exteme: M. Eric Garnier
C.N.R.S.
3
^
~,^£
z^
J
77 \^_
SOMMAIRE
Cette etude a porte sur 22 especes herbacees sauvages de la region de Sherbrooke, au sud de la province de Quebec, Canada. La croissance des plantes s'est deroulee dans quatre combinaisons differentes de conditions controlees : avec une forte ou une faible concentration minerale et une forte ou une faible intensite d'eclairement. La duree de croissance des plantes a ete de 35 jours. Pendant cette periode, plusieurs parametres ont ete mesures. Pour analyser nos resultats, nous avons eu recours a 1'analyse des pistes. Cette technique permet entre autres de quantifier les liens directs et indirects entre les variables d'un modele et propose un test statistique pour tester la validite des modeles.
Les resultats ont montre que la morphologie des feuilles et particulierement la surface speciflque foliaire joue un role determinant dans Ie comportement des attributs physiologiques et d'allocation. En effet, la surface speciflque determine les echanges gazeux des plantes en agissant sur la concentration d'azote dans les feuilles. Aussi, son augmentation provoque une augmentation directe de la vitesse specifique de croissance des plantes. Par contre, 1'augmentation de la surface specifique foliaire provoque une diminution de la vitesse d'assimilation nette et du rapport de biomasse des feuilles sur la biomasse des racines. Comme ces deux derniers attributs provoquent une augmentation directe de la vitesse specifique de croissance, leur effet est reduit par leur lien negatif avec la surface specifique foliaire. Par ailleurs, la surface speciflque foliaire est determinee en grande partie par la concentration d'eau dans les feuilles (densite des tissus) et 1'epaisseur des limbes. L'epaisseur de la nervure centrale joue un role moins important dans Pexplication des variations interspecifiques de la surface specifique foliaire.
Le changement des conditions de culture des plantes n'a pas cause de changements au niveau des liens entre les differents attributs qui determinent la vitesse specifique de croissance. Toutefois, les moyennes et variances de ces attributs ont beaucoup change avec les changements des conditions environnementales. Done, tout se passe comme si les plantes
modifiaient quantitativement leurs attributs morphologiques, physiologiques et d'allocation tout en gardant inchanges les liens entre ces parametres en vue de minimiser les reductions de leur vitesse specifique de croissance.
REMERCIEMENTS
Je tiens a remercier vivement Ie Dr. Bill Shipley pour la conflance qu'il m'a accordee en m'accueillant dans son laboratoire, pour ses conseils et son soutien constmctif tout Ie long de mon travail. Je dois beaucoup a mes collegues de 1'Universite Sidi Mohamed Ben Abdellah de Fes, Maroc pour leur comprehension et leur aide qui m'ont permis de realiser ce projet. Mes remerciements vont aussi aux professeurs membres de mon comite de doctorat Gaston Beaumont, Gilles Grenier et Robert van Hulst qui m'ont aide et m'ont soutenu du debut a la fin de ce travail. Je dois aussi remercier Ie professeur Eric Gamier pour avoir accepte de participer au jury, Eric Cloutier pour avoir realise Ie dosage d'azote dans les feuilles et Caroline Cloutier pour avoir accepte de relire ce memoire et Denis Charlebois qui a mesure les spectres de reflectance dans la chambre de croissance.
L'universite de Sherbrooke et 1'agence canadiemie du developpement international (ACDI) m'ont offert une bourse qui m'a permis de realiser mon projet dans de bonnes conditions materielles. II m'est agreable de leur exprimer ici ma reconnaissance.
Je tiens enfin a remercier tous les professeurs et techniciens du laboratoire d'ecologie pour leurs discussions fructueuses, leur support, et leur gentillesse et leur bonne humeur qui rendent Ie travail agreable au sein de cette equipe. Je ne saurais oublier tous mes amis, et specialement Anne Genier, qui n'ontjamais cesse de m'encourager et avec qui j'ai passe de bans moments lors de mon sejour a Sherbrooke.
TABLE DES MATIERES
SOMMAIRE... i
REMERCIEMENTS... iii
TABLE DESMATIERES... iv
LISTEDES TABLEAUX... vii
LISTEDES FIGURES... viii
LISTEDESABREVIATIONS... x LISTEDES ANNEXES... xi WTRODUCTION... 1 1 Importance de la VSC... 1 a) Especesruderales... 5 b) Especes competitrices... 5
c) Especes tolerantes au stress... 6
2 Approches pour decrire la VSC ... 7
2.1 Evans 1972... 7
2.2 Potter et Jones 1977... 9
2.3 Ingestad 1982... 10
2.4 Lambers et Poorter 1992... 10
3 Les attributs qui controlent les variations de la VSC... 13
3.1 La photosynthese... 13
3.2 Respiration... 15
3.3 Allocation de biomasse... 15
3.4 Morphologie des feuilles... 17
CHAPITRE 1 MATERIELS ET METHODES... 22
1.1 Choix des especes... 22
1.2 Recolte des graines et germination... 22
1.3 Conditions de culture... 24
1.4 Solution nutritive... 26
1.5 Mode de recolte et de mesure des differents attributs ... 29
1.6 Analyses statistiques ... 36
1.6.1 Analyse de variance et correlation... 36
1.6.2 Analyse des Pistes... 36
CHAPITRE 2 - RESULTATS ET DISCUSSIONS... 50
2.1 Analyse de la variance et les reponses des attributs des especes (moyennes) en fonction des traitements experimentaux... 50
2.1.1 Composantes de la variance... 50
2.1.2 Effet de 1'intensite lumineuse et de la disponibilite des mineraux nutritifs surles attributs... 51
2.1.2.1 Attributs relies aux echanges gazeux... 51
2.1.2.2 Morphologie des feuilles...58
2.1.2.3 Parametres d'allocation de biomasse... 64
2.1.2.4 Vitesse specifique de croissance (VSC)... 68
2.2 Analyse des pistes... 69
2.2.1 Surface specifique des feuilles (SSF)... 69
2.2.1.1 Patron de correlation entre la surface specifique
foliaire et les variables dont elle depend
en fonction des traitements... 692.2.1.2 Modele de Shipley... 70
2.2.1.3 Effet de 1'eclairement et de la disponibilite en mineraux nutritif sur Ie comportement de la surface specifique des feuilles... 74
2.2.1.4 Discussion... 83
2.2.2 Photosynthese nette (PN)... 88
2.2.2.1 Photosynthese nette exprimee par rapport a la surface (PNs)... 89
2.2.2.2 Photosynthese nette exprimee par rapport a labiomasse des feuilles (PNm)... 97
2.2.2.3 Discussion... 102
2.2.3 Vitesse specifique de croissance (VSC)... 114
2.2.3.1 Effet des traitements sur les moyennes et les variances des composantes de la croissance... 114
2.2.3.2 Modelisation... 118 2.2.3.3 Discussion... 125 2.2.3.4 Modeles generaux... 130 2.2.3.5 Discussion... 136 CONCLUSIONS GENERALES... 139 ANNEXES... 146 BIBLIOGRAPHIE... 151
LISTE DES TABLEAUX
1. Liste des 22 especes utilisees etleur affiliation taxinomique... 23
2. Composition chimique de la solution nutritive riche en elements nutritifs (N)... 30
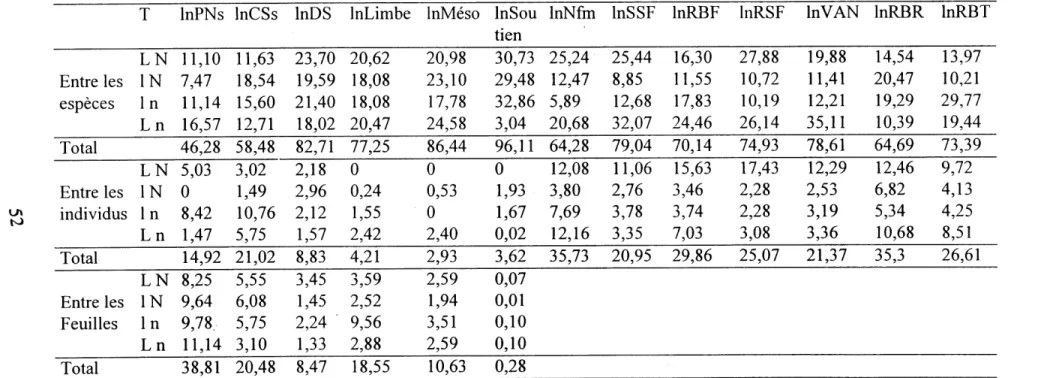
3. Pourcentages de variabilite des attributs aux differents niveaux de la hierarchisation et leurs sommes en fonction des traitements... 52
4. Moyennes et ecarts-types des attributs en fonction des differents tmitements... 53
5. Logarithmes neperiens des moyennes et ecarts-types des attributs en fonction des differents traitements... 54
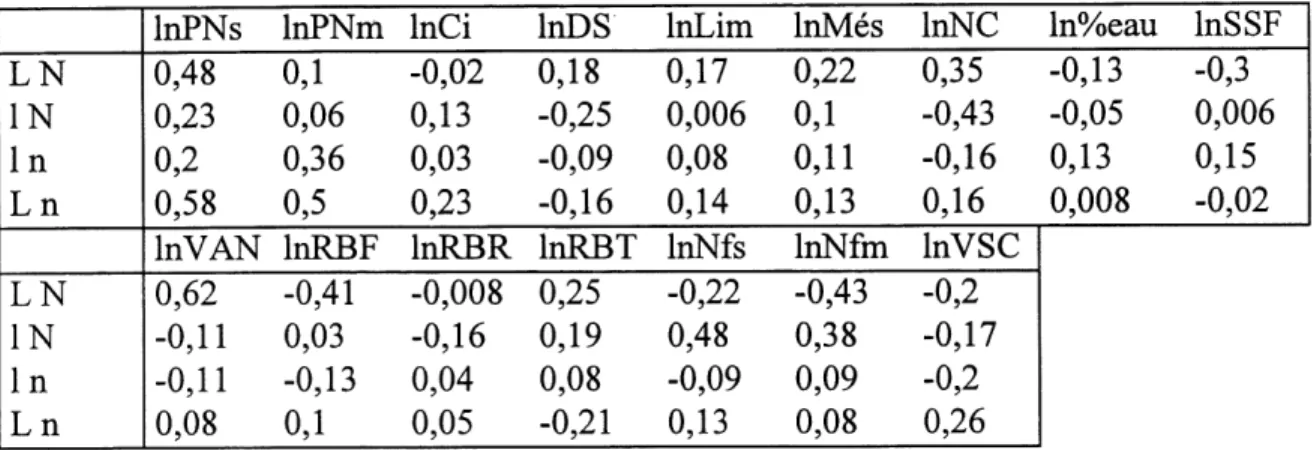
6. Correlations entre la photosynthese nette par unite de surface foliaire et les autres attributs... 56
7. Correlations entre la photosynthese nette par unite de biomasse foliaire et les autres attributs... 56
8. Correlations entre la conductance des stomates par unite de surface foliaire et les autres attributs... 57
9. Correlations entre la concentration intracellulaire de CO^ et les autres attributs... 57
10. Correlations entre la vitesse d' assimilation nette et les autres attributs... 58
11. Correlations entre la densite des stomates et les autres attributs... 59
12. Correlations entre Fepaisseur du mesophylle et les autres attributs... 61
13. Correlations entre 1'epaisseur du tissu de soutien et les autres attributs... 62
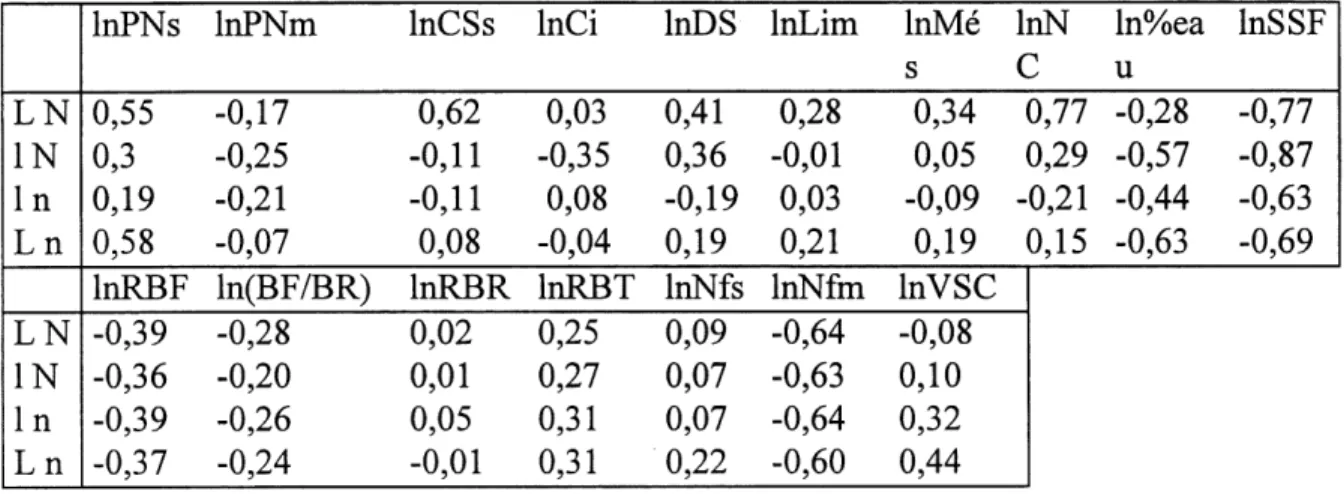
14. Correlations entre la surface specifique foliaire et les autres attributs ... 63
15. Correlations entre Ie rapport de biomasse foliaire et les autres attributs... 64
16. Correlations entre Ie rapport de biomasse des racines et les autres attributs... 66
17. Correlations entre Ie rapport de biomasse des tiges et les autres attributs ... 67
18. Correlations entre la concentration d'azote dans les feuilles et les autres attributs... 68
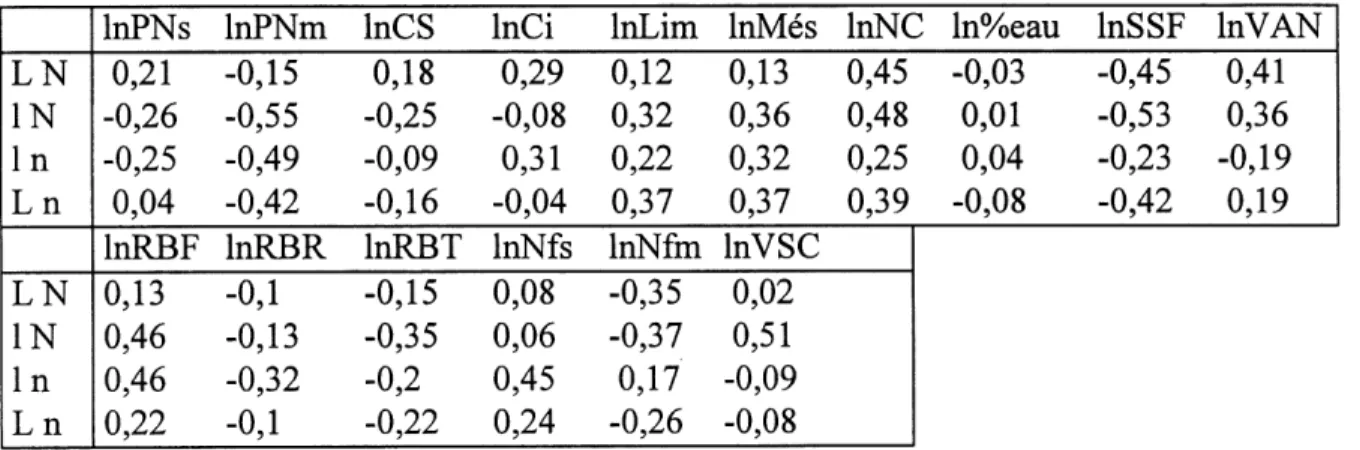
19. Correlations entre la vitesse specifique de croissance et les autres attributs... 69
20. Logarithmes neperiens des moyennes des differents attributs qui determinent la croissance des plantes en fonction des differents traitements... 114
LISTE DES FIGURES
1. Spectre de reflectance a 1'interieur de la chambre de croissance
pendantl'eclairement fort... 27
2. Spectre de reflectance a 1'interieur de la chambre de croissance pendant 'eclairementfaible... 28
3. Exemple de modele obtenu avec 1'analyse des pistes... 37
4. Exemple de modele obtenu avec des donnees generees artificiellement... 41
5. Exemple de modele proche du modele reel... 42
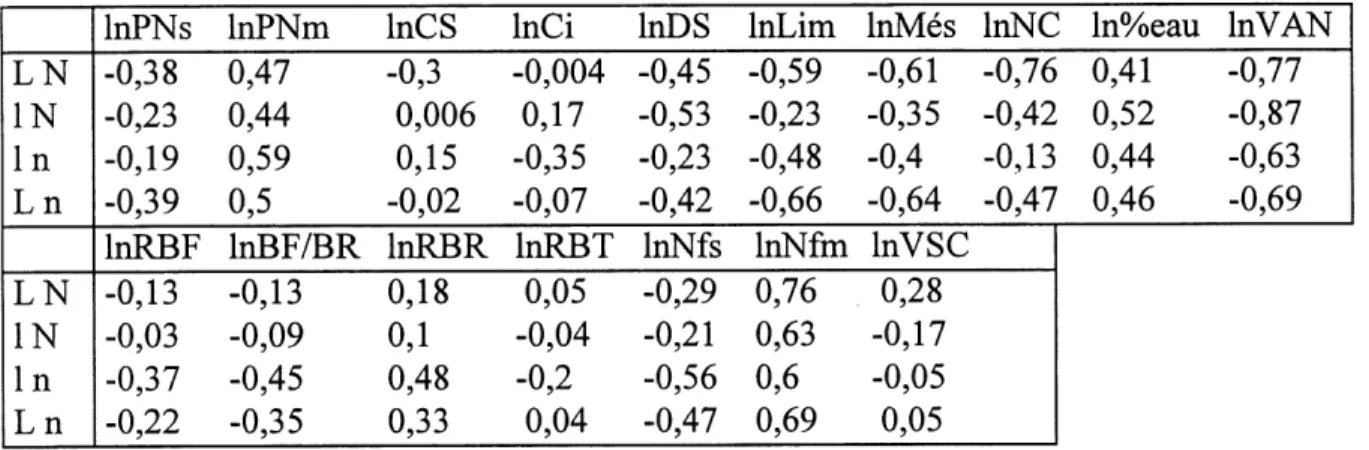
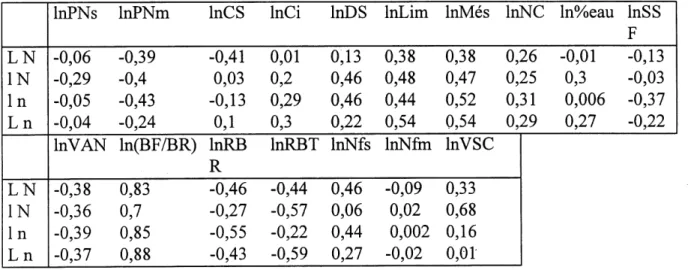
6. Correlations entre les logarithmes des differents attributs qui determinent la surface specifique foliaire pendant les traitements 'L N' et '1 N'... 71
7. Correlations entre les logarithmes des differents attributs qui determinent la surface specifique foliaire pendant les traitements '1 n' et 'L n'... 72
8. Premier modele de la surface specifique foliaire d'apres Shipley (1995)... 73
9. Deuxieme modele de la surface specifique foliaire d'apres Shipley (1995)... 73
10. Modeles de la surface specifique foliaire en fonction des quatre traitements... 75
11. Modele detaille de la surface specifique foliaire en fonction des quatretraitements... 80
12. Modele des echanges gazeux par unite de surface foliaire d'apres Shipley et Lechowicz... 90
13. Modele des echanges gazeux par unite de surface foliaire en fonction des quatretraitements... 93
14. Modele des echanges gazeux par unite de surface foliaire incluant la densite des stomates en fonction des quatretraitements... 96
15. Modele des echanges gazeux par unite de biomasse foliaire en fonction desquatretraitements... 98
16. Modele des echanges gazeux par unite de biomasse foliaire incluant la densite des stomates... 101
17. Correlations entre les logarithmes neperiens des differents attributs qui
determinent la VSC pendant les traitements 'LN' et 'IN'...116
18. Correlations entre les logarithmes neperiens des differents attributs qui
determinent la VSC pendant les traitements '1 n' et 'L n'... 117 19. Modele de la vitesse specifique de croissance selon Shipley et McKenna... 120 20. Modele de la vitesse specifique de croissance en fonction des quatre traitements... 121 21. Modele de la vitesse specifique de croissance obtenu avec les donnees de Poorter et
Remkes (1990)... 127
22. Modele general par unite de surface foliaire pendant Ie traitement 'L N'... 132 23. Modele general par unite de biomasse foliaire pendant Ie traitement 'L N' ... 135
LISTE DES ABREVIATIONS
%eau : Pourcentage d'eau dans les feuilles (%)
BF/BR : Biomasse des feuilles sur biomasse des racines (g.g )
Ci : Concentration intracellulaire de C02 (ppm)
CSm : Conductance des stomates par unite de biomasse foliaire (^imoles.g .s ) CSs : Conductance des stomates par unite de surface foliaire (jLimoles.m .s ) DS : Densite des stomates (nombre de stomates.mm'2)
Lim : Epaisseur du limbe (p-m)
Meso : Epaisseur du mesophylle (|J-m) NC : Epaisseur de la nervure centrale ((Jm)
Nfm : Concentration d'azote par unite de biomasse foliaire (mg.g'1) Nfs : Concentration d'azote par unite de surface foliaire (mg.cm'^)
PNm : Photosynthese nette par unite de biomasse foliaire (|Limoles.g'l.s ) PNs : Photosynthese nette par unite de surface foliaire (Hmoles.m''''.s'1)
PS : Folds sec des feuilles (g)
RBF : (LWR), Rapport de biomasse foliaire (g.g )
RBR : (RWR), Rapport de biomasse des racines (g.g )
RBT : (SWR), Rapport de biomasse des tiges (g.g )
RSF : (LAR), Rapport de surface foliaire (cm^.g'1)
SF : Surface foliaire (cm^)
Sout: Epaisseur du tissu de soutien (pm)
SSF : (SLA), Surface specifique foliaire (cm2.g-l)
VAN : CNAR), Vitesse d'assimilation nette (mg.cm'2.j ) VSC : (RGR), Vitesse specifique de croissance (g.g .j )
LISTES DES ANNEXES
Annexe 1. Moyenne par espece des parametres anatomiques, physiologiques et de
INTRODUCTION
La vitesse specifique de croissance (VSC) est un element clef du fonctionnement des plantes superieures dans leur habitat naturel. La VSC est definie comme etant la biomasse produite par unite de temps par unite de biomasse deja existante. Plusieurs chercheurs ont constate depuis longtemps que les especes vegetales ne possedent pas la meme VSC malgre 1'uniformite des conditions environnementales (Jarvis et Jarvis, 1964 ; Clarkson, 1965 ; Higgs et James, 1969 ;
Grime et Hunt, 1975 ; Chapin, 1980 ; Shipley et Keddy, 1988 ; Poorter, 1989 ; Poorter et
Remkes, 1990 ; Gamier, 1991 ; Davidson, 1993 ; Dudt et Shure, 1994). Ces auteurs aussi ont remarque que cette difference inherente aux especes persiste meme lorsque ces conditions sont optimales. En effet, les especes qui occupent en general des milieux productifs possedent une VSC plus elevee que celui des especes qui colonisent des milieux infertiles. Cette difference persiste meme quand ces deux types d'especes se trouvent dans des conditions identiques et productives. Par ailleurs, les especes qui proviennent des endroits ou il n'y a pas de stress physique (manque chronique d'elements nutritifs, de lumiere, d'eau) ont une VSC plus elevee que celui des especes qui occupent naturellement les zones ombragees (Pons, 1977 ; Corre, 1983), les zones arides (Rozijn et van der Werf, 1986) et les zones salees (Ball, 1988). Done,
la difference interspecifique des VSC pourrait etre adaptative (Hunt et Burnett, 1973 ; Grime
et Hunt, 1975 ; Chapin, 1980 ; Hunt et Nicholls, 1986 ; Poorter, 1989 ; Comellisen et
^,.,1996).
1. Importance de la VSC
L'etude de la VSC presente un interet majeur au niveau agronomique, forestier et ecologique. En effet, les agronomes etaient les premiers a s'interesser a la VSC en vue de comprendre les mecanismes qui determinent Ie potentiel de croissance des plantes cultivees afin d'ameliorer leur rendement. De meme, les forestiers etudient la VSC en vue de comprendre les parametres qui Ie controlent pour choisir les especes qui presentent Ie meilleur potentiel pour augmenter la
production du bois d'oeuvre et du bois de pate. En ecologie, 1'interet d'etudier la VSC est qu'il permet de comprendre les mecanismes qui controlent Ie fonctionnement et la distribution des especes vegetales dans les habitats naturels. En effet, des chercheurs ont montre a 1'aide de la VSC,^ mesuree dans des conditions controlees et optimales qu'il y a un lien entre Ie type d'habitat et Ie type d'espece qui 1'occupent. Ainsi, les plantes qui ont une croissance rapide occupent surtout des milieux productifs, alors que les especes qui ont une croissance lente
colonisent plutot des milieux infertiles (Grime et Hunt, 1975 ; Chapin, 1980 ; Shipley et
Keddy, 1988 ; Poorter, 1989 ; Poorter et Remkes, 1990 ; Garnier, 1991).
Pendant la succession secondaire, les plantes investissent la biomasse produite dans differents organes en fonction de la ressource pour laquelle elles se competitionnent (Tilman, 1988). Ainsi, F allocation de biomasse vers les racines est importante lorsque les ressources nutritives du sol sont limitees. Par contre, dans les habitats riches, les plantes augmentent plutot leur biomasse aerienne en investissant dans les feuilles et aussi les tiges qui leur permettent d'aller chercher la lumiere necessaire a 1'assimilation photo synthetique du carbone.
Plusieurs chercheurs se sont interesses aux differences interspecifiques de la VSC pour comprendre la distribution des especes vegetales dans les milieux naturels (Evans, 1972 ;
Grime et Hunt, 1975 ; Shipley et Keddy, 1988 ; Poorter et Remkes, 1990 ; Grime et Campbell,
1991 ). Tous ces auteurs s'accordent pour dire que les especes qui occupent des milieux infertiles possedent des VSC^gx plus faibles que celui des especes qui occupent des milieux fertiles. La question est de savoir pourquoi il y a cette difference et quel avantage les especes vegetales tirent du fait d'avoir une VSC faible ou eleve selon Ie type d'habitat. Dans les milieux productifs, la dominance des especes ayant des VSC,^ elevees est comprehensible. Ces especes sont capables de coloniser rapidement les espaces a cause de leur forte capacite de puiser et d'utiliser les ressources pour accroitre leur biomasse. Ainsi, ces especes arrivent a priver les especes a croissance lente de la lumiere et des ressources du sol. Chapin (1980) a explique que la presence des especes a faible VSC,nax dans les milieux pauvres en elements
nutritifs est due a leur faible acquisition et utilisation des elements nutritifs. Selon cet auteur, ceci leur permet d'utiliser les ressources de £09011 logique sans compromettre leur existence. Mais d'apres Lambers et Poorter (1992), 1'habilite a utiliser les ressources serait une consequence plutot qu'une cause de la difference interspecifique de la VSC. Autrement dit, dans les milieux infertiles, les plantes possedent une suite d'attributs qui ont comme effet de reduire la VSC^- Chapin (1980) a explique ce phenomene par Ie fait que les especes qui ont une VSC faible possedent un certain nombre de traits qui leur permettent de coloniser les milieux pauvres. Selon cet auteur, ces especes ont une faible capacite d'acquerir les ressources, faible transpiration, un faible taux de photosynthese nette (PN), une faible allocation de biomasse vers les meristemes et leurs tissus sont riches en substances de defense contre les herbivores. Cette hypothese suggere aussi que les especes qui ont une croissance lente incorporent moins de glucides dans leur biomasse de structure. Ainsi, elles peuvent former des reserves pour une croissance ulterieure au cas ou il y aurait un manque de ressources. Mais les feuilles de ces plantes sont souvent plus riches en sclerenchyme et en composes secondaires. Ceci les differencie des especes qui ont une croissance rapide et qui favorisent la croissance en allouant leurs glucides aux meristemes de croissance. Toutefois, il semble que la mise en reserve des glucides chez les especes a croissance lente ne leur permette pas de sm-vivre pendant les periodes de manque de ressources. Elle leur permet plutot de tamponner les fluctuations de la disponibilite des ressources (Lambers et Poorter, 1992).
II est clair que Ie deficit en elements nutritifs et les perturbations affectent independamment plusieurs processus structurels et physiologiques a la fois (Mahmoud et Grime, 1976 ; Chapin et al., 1988 Grime et al., 1988 ; Evans, 1989b ; Chapin, 1991). Niemann et al. (1992), Elberse et Berendse (1993) et Cornellisen et al. (1996) pensent que la pression selective a pousse les especes qui occupent les milieux infertiles a investir beaucoup de biomasse foliaire par unite de surface foliaire. Done, ces especes ont une faible surface specifique foliaire (SSF, rapport de la surface des feuilles sur la biomasse des feuilles). Aussi, ces especes ont un faible investissement dans la biomasse totale des feuilles, done dans Ie rapport de biomasse foliaire
(RBF, rapport de biomasse seche des feuilles sur la biomasse seche totale de la plante). Enfin, les feuilles de ces especes ont une longevite plus importante, leur permettant ainsi d'eviter des investissements repetes dans la formation de nouvelles feuilles (Reich et al,, 1991 ; Reich et al., 1992). La faible SSF chez les plantes qui ont une croissance faible peut avoir plusieurs causes. En effet, la cause peut etre un investissement important dans les materiaux de paroi cellulaire et les composes secondaires de defense centre les herbivores (Coley, 1988 ; Lambers et Poorter, 1992), une epaisseur de feuilles importante (Pammenter et al., 1986 ; Komer et Diemer, 1987 ; Kaminsky et al., 1990 ; Shipley, 1995), une faible densite de tissus dans les feuilles (Dijkstra et Lambers, 1989; Witkowski et Lamont, 1991 ; Kebede et al., 1994; Gamier et Laurent, 1994 ; Van Arendonk et Poorter, 1994), ou un faible pourcentage d'eau
dans les feuilles (Dijkstra et Lambers, 1989 ; Witkowski et Lamont, 1991 ; Shipley, 1995).
Meme dans les conditions de faible disponibilite en ressources minerales, du moins dans les quelques etudes qui ont etudie cette question sur une courte duree, les especes qui ont une croissance rapide gardent une croissance plus rapide que les especes qui ont une croissance inherente lente (Christie et Moorbey, 1975 ; Grime et Hunt, 1975 ; Poorter et Remkes, 1990). Nous pouvons done penser que la selection naturelle n'agit pas directement au niveau de la VSC, mais plutot au niveau des attributs qui Ie determinent comme la vitesse d'assimilation nette, 1'allocation de biomasse ou la morphologie des feuilles (Lambers et Dijkstra, 1987 ; Lambers et Poorter, 1992 ; Niemann et al., 1992). Done, la VSC,^ a- elle seule n'explique pas 1' absence d'especes qui ont une croissance rapide dans les milieux infertiles. Par ailleurs, d'autres chercheurs pensent que les especes qui ont une croissance rapide gardent toujours une translocation importante vers les feuilles, meme dans les conditions de faible disponibilite en ressources. Puisque ces especes ont des feuilles de courte longevite, les mineraux captes par leur feuilles vont retoumer rapidement dans Ie sol. Les especes a faible VSC possedent des feuilles d'une longue esperance de vie, done ces especes peuvent capter et garder les mineraux perdus par les premieres especes. Ceci va contribuer a 1'appauvrissement rapide des sols et a 1'elimination des especes a forte VSC au profit des especes qui ont une faible VSC^ (Berendse et Elberse, 1989). En effet, ces demieres especes emmagasinent plus de mineraux
dans leurs tissus (plus de longevite) et par consequent economisent mieux les ressources que les especes qui out une VSC elevee. Done, la disparition des especes qui ont une VSC^ elevee des milieux infertiles serait due a leur inadaptation a ces milieux.
Plusieurs chercheurs ont etudie la distribution naturelle des especes vegetales et les facteurs qui la determinent (Grime et Hunt, 1975 ; et Tilman, 1988). Ces auteurs ont mis 1'accent sur Fimportance de la VSC et des attributs qui Ie determinent dans la distribution des especes vegetales. Ainsi, Grime et Hunt (1975) ont compare la VSC,^ chez 132 especes herbacees originaires de differents milieux naturels des environs de Sheffield en Angleterre. Ces especes ont ete soumises a differentes conditions de stress et de perturbations et les conclusions de leur etude classent les especes en trois groupes.
a) Especes ruderales
Ce sont les especes qui ont une VSC elevee et qui colonisent les milieux riches en elements nutritifs (absence de stress nutritionnel), mais perturbes. Ces conditions sont reunies en general dans les prairies cultivees, et la majorite de ces especes constituent ce que les agronomes appellent communement les 'mauvaises herbes'. Ces especes ont une courte esperance de vie et sont caracterisees par un investissement precoce d'une proportion importante de leur biomasse dans leur appareil reproducteur.
b) Especes competitrices
Ce sont les especes que 1'on trouve dans les milieux riches en elements nutritifs (absence de stress nutritionnel), non perturbes et dans les sites bien eclaires. Ce groupe de plantes possede aussi une VSC elevee du a leur allocation importante de biomasse vers les meristemes de croissance mais elles investissent moins dans la reproduction sexuelle et plus dans la reproduction vegetative. Aussi, elles ont une esperance de vie plus longue.
c) Especes tolerantes au stress
Ce groupe est compose par les especes qui occupent les milieux ou il y a presence de stress nutritionnel, mais ou les perturbations sont rares. Ce sont les especes que 1'on trouve habituellement dans les regions arides, pauvres en elements nutritifs ou peu eclaire. Contrairement aux deux premiers groupes, les especes tolerantes au stress nutritionnel ont une VS C faible. En outre, ces especes allouent leur biomasse, en partie, aux stmctures de reserve pour affronter d'eventuelles periodes de carence en elements nutritifs.
Tilman (1988) quant a lui, a propose une theorie intuitive basee sur Ie mode d'allocation de biomasse pour expliquer la distribution des especes vegetales dans les milieux naturels. Pour cet auteur, c'est la maniere dont les especes vegetales allouent leur photosynthetates qui determine leur VSC^ Tilman (1988) suppose que puisque Ie poids sec des plantes est constitue en grande partie de carbone (40 a 50%), tout investissement dans les tissus qui fixent cet element contribue a augmenter Ie taux de croissance. Par centre, 1'allocation vers les tissus non assimilateurs de carbone et qui respirent diminue Ie taux d'accumulation nette du carbone et done la croissance. Done, selon Tilman, les especes qui investissent les produits de la photosynthese dans les tissus non assimilateurs (tiges ou racines), ont une VSC,^^ plus faible que celle des especes qui investissent relativement plus de photosynthetates dans les tissus assimilateurs ( feuilles). II a aussi suggere que dans les milieux riches, la competition se fait surtout pour la lumiere. Done, les especes qui ont une VSC^^ plus elevee et une taille plus elevee sont favorisees. Par centre, dans les milieux infertiles, la competition se joue plutot au niveau des elements nutritifs du sol, et par consequent, les especes qui ont un systeme radiculaire plus important et plus performant sont plus competitives. Tilman a propose Ie modele suivant pour resumer sa theorie.
VSC = (BF/B)PNm - r
BF : Biomasse de feuilles B : Biomasse totale
PNm: Photosynthese maximale r : Taux de respiration
Ce modele mathematique de prediction rend cette theorie plus interessante que celle de Grime et Hunt. En effet, ce modele nous donne la possibilite de tester la theorie et de la refuter eventuellement. Ainsi, plusieurs etudes ont supporte la theorie de Tilman (Poorter et Remkes, 1990 mais seulement chez les dicotyledones ; Walters et al., 1993b ; Comellisen et al., 1996). Cependant, la correlation de la VS C avec la SSF etait toujours plus importante que celle avec Ie RBF (BF/B). Par centre, d'autres chercheurs ont partiellement ou entierement refute 1'hypothese de Tilman (Shipley et Peters, 1990b ; Aerts et al., 1991 ; Ingestad et Agren, 1991 ; Komer, 1991 ; Gamier, 1992 ; Oiffet al., 1990 ; Gleeson et Tilman, 1994).
2. Approches pour decrire la VSC
Plusieurs chercheurs ont propose des modeles pour decrire les facteurs qui determinent les variations de la VSC. Ces differents auteurs ont utilise differents attributs selon ce qu'ils pensent etre Ie facteur clefdans les mecanismes de croissance.
2.1 Evans 1972
Dans ce modele, Evans a decompose la VSC en trois attributs ; la vitesse d'assimilation nette
(VAN), Ie rapport de biomasse foliaire (RBF) et la surface specifique foliaire (SSF). II faut
noter que Ie produit de la SSF et Ie RBF donne Ie rapport de surface foliaire (RSF).
VSC=VAN*RSF (1)
RSF=RBF*SSF (2)
VSC=VAN*RBF*SSF (3)
Par consequent, la formule (3) devient:
(l/B)*(dB/dt) = [(l/SF)*(dB/dt)]*[BF/B]*[SF/BF] (4)
VSC : Vitesse specifique de croissance = (l/B)*(dB/dt) g/g/j
VAN: Vitesse d'assimilation nette = (l/SF)*(dB/dt) g/m2/j
RSF : Rapport de surface foliaire = SF/B m2/g
RBF : Rapport de biomasse foliaire = BF/B g/g
SSF : Surface specifique foliaire = SF/BF m /g
B : Biomasse seche totale de la plante g SF : Surface des feuilles m BF : Biomasse seche des feuilles g
Dans ce modele, la VAN est consideree comme un attribut physiologique car elle est la resultante des echanges gazeux : la photosynthese nette et la respiration. Mais cet attribut est loin d'etre une composante purement physiologique, car il depend aussi de 1'allocation de la biomasse, de la composition chimique des feuilles et de la surface des feuilles. Le RBF est considere comme un attribut d'allocation, car il represente Ie taux de biomasse investie dans les feuilles par rapport a celle investie dans les autres parties de la plante. Enfin, la SSF est consideree comme un attribut morphologique, car elle est la resultante de la morphologie des feuilles, a savoir la quantite de tissus dans les feuilles par rapport a leur surface.
Ce modele empirique laisse croire que tout investissement de biomasse dans les tissus autres que les feuilles reduit la VSC. Cette approche considere done les racines et tiges comme de
simples consommateurs de glucides, et ne tient pas compte du role des racines dans F acquisition des ressources du sol.
2.2 Potter et Jones 1977
Ces deux chercheurs ont propose une approche basee sur la variation de la vitesse specifique de croissance des feuilles en fonction du temps. Selon eux, cet attribut donne une meilleure prediction des variations de la VSC. Leur modele est forme par Ie produit de la VAN et du taux de changement du RSF. Done, a 1'opposition du modele d'Evans, ce modele est dynamique.
VSCF = VAN*dRSF (5)
VSCF : VSC des feuilles = (l/SF)*(dSF/dt) m2/m2/j
VAN : Vitesse d'assimilation nette = (l/SF)*(dB/dt) g/m2/j
dRSF : Variation du RSF = (dSF/dt)/(dB/dt) m2/g
Done, la formule (5) devient:
(l/SF)*(dSF/dt) = (l/SF)*(dB/dt);l![(dSF/dt)/(dB/dt)] (6)
Comme nous pouvons Ie constater, ce modele est base sur les proprietes des feuilles. En effet, ces deux auteurs pensent que la VSC des feuilles va dormer une bonne idee de la VSC de la plante entiere. Car, les feuilles sont les organes principaux de la fixation de carbone, et done, de la production de biomasse. Malheureusement, cette suggestion n'est pas toujours vraie, surtout lorsque la plante investit ses produits de photosynthese dans des organes autres que les feuilles. Dans ce cas, la VSC de la plante continue d'augmenter alors que celle des feuilles
reste stable ou augmente legerement. Done, nous pouvons reprocher a ce modele les memes lacunes que celles du modele d'Evans.
2.3 Ingestad 1982
Ce chercheur propose un modele ou la VSC depend de la productivite et de la concentration d'azote dans la plante. Selon Ingestad, bien que Ie carbone et 1'azote soient les elements principaux dans la nutrition des plantes, c'est souvent 1'azote qui est Ie premier element limitant dans les milieux naturels.
VSC = PA*N (7)
VSC : Vitesse specifique de croissance = (l/B)(dB/dt) g/g/j
PA: Productivite d'azote = (l/N)*(dB/dt) g/mg/j
N : Concentration d'azote = N/B mg/g
La formule (7) devient:
(l/B)(dB/dt) = (l/N)*(dB/dt)*(N/B) (8)
Malgre 1'originalite de ce modele, nous pouvons lui reprocher Ie fait qu'il ne tient compte ni de la morphologie, ni des echanges gazeux des plantes.
2.4 Lambers et Poorter 1992
Ces auteurs ont tenu a completer les lacunes du modele d'Evans. Pour cela, ils ont ajoute dans leur modele les pertes de biomasse par respiration, volatilisation et exsudation.
VSC = [(PNs*SSF*RBF)-(RFm;(SRBF)-(RTm*RBT)-(RRm*RBR)] / [C-Exm-VOLm] (9)
VSC : Vitesse specifique de croissance = (l/B)*(dB/dt) g/g/j
PNs : Photosynthese nette exprimee par unite de surface foliaire ^mole/m/s
SSF : Surface specifique foliaire = SF/BF m2/g
RBF : Rapport de biomasse foliaire = BF/B g/g
RFm : Respiration foliaire par unite de biomasse foliaire nmole/g/s RTm : Respiration des tiges par unite de biomasse des tiges nmoles/g/s
RBT : Rapport de biomasse des tiges = BT/B g/g
RRm : Respiration radiculaire par unite de biomasse radiculaire nmoles/g/s RBR : Rapport de biomasse des racines = BR/B g/g
Exm : Taux d'exsudation par unite de biomasse mg/g/j
VOLm : Taux de volatilisation par unite de biomasse mg/g/j
C : Concentration en carbone mmole C/g
Ces ajouts ont rendu ce modele tres complexe car, en plus du nombre eleve de variables, plusieurs des variables sont difficiles a mesurer ou meme a estimer.
Ces chercheurs ont imagine un deuxieme modele qui rendrait mieux compte des variations de la VSC lorsque Ie rapport entre 1'augmentation des elements nutritifs (azote) et celui de la biomasse est constant.
VSC = RBR*TAA/N (10)
TAA: Taux d'absorption (radiculaire) d'azote N: Concentration d'azote dans la plante
Avec ce deuxieme modele, qui ne tient compte que de 1'activite des racines dans 1'acquisition de 1'azote, tous les autres attributs morphologiques et physiologiques sont delaisses, ce qui Ie rend incomplet par rapport au premier modele.
Enfin, il semble que 1'investissement dans les composes secondaires joue aussi un role important dans la determination de la VSC. En effet, plusieurs auteurs pensent qu'il y a une correlation negative entre la VSC et la concentration des composes secondaires dans les tissus des plantes (Bryant et al., 1983 ; Coley, 1983 ; Rosenthal et Kotanen, 1994). Cependant, aucun des modeles presentes plus haut n'en tient compte.
Ces differentes approches illustrent bien les difficultes de decrire la VSC a cause du nombre eleve de facteurs qui peuvent controler ses variations. Aussi, les variables utilisees sont quelquefois difficiles a mesurer et Ie probleme de causalite entre les variables n'est pas aborde. De plus, il est clair que I'influence des differents attributs varie en fonction des conditions environnementales. Ainsi, a chaque fois qu'un attribut change en fonction des conditions externes, il affecte une chaine de processus a 1'interieur de la plante. Ces effets se manifestent par des modifications au niveau de la morphologie, de la physiologie ou de la constitution chimique des tissus. La question est done de savoir pourquoi les modeles cites plus haut ne sont pas capables de cerner tous ces problemes.
A notre avis, si ces modeles n'ont pas tente de decrire les liens directs et indirects entre les attributs, ni de decrire les reponses des plantes dans des conditions environnementales differentes, c'est parce qu'il leur manque un moyen efflcace pour Ie faire. Justement, une des originalites de notre travail est de combler cette lacune en utilisant 1'analyse de pistes (Path analysis) comme outil statistique. Cette technique permet, 1) de quantifier les liens directs et indirects entre les differentes variables en fonction des changements des conditions de culture des plantes. 2) De representer tous ces liens en forme de modeles de prediction qui offrent la
possibilite de les refuter. En plus, ces modeles tiennent compte de la causalite hypothetique entre les variables au lieu de s'en tenir a leurs correlations. Enfin, cette technique offre la possibilite de tester les resultats (voir la section materiel et methode). Le premier probleme que nous pouvons rencontrer avec cette technique est 1'absence des modeles dans la litterature. Dans ce cas, pour tester Ie modele, Ie chercheur utilise les memes donnees qui ont servi a 1' elaboration du modele. Ce probleme peut etre depasse facilement en utilisant les resultats des etudes d'autres chercheurs. Done, la confiance dans les resultats viendra d'abord de 1'utilisation d'un grand nombre d'echantillons, mais surtout de la repetition des memes resultats.
3. Les attributs qui controlent les variations de la VSC
Plusieurs etudes menees dans des conditions controlees chez plusieurs especes ont montre que la VSC depend des attributs morphologiques, physiologiques et d'allocation. Parmi ces facteurs, il y a la taille des graines (Stebbins, 1976 ; Fenner, 1983 ; Gross, 1984 ; Maranon, 1988 ; Shipley et Peters, 1990a; Comellisen et al., 1996), la photosynthese nette. Ie RBF, la SSF et la teneur en composes secondaires. Avant de voir comment tous les attributs interagissent entre eux, nous aliens d'abord passer en revue leur effet separement. Bien entendu, nous savons que la variation d'un attribut va affecter les autres attributs d'une £09011 directe ou indirecte.
3.1 La Photosynthese
La relation entre la photosynthese nette, (PN, la photbsynthese brute moins la photorespiration) et la VSC n'est pas tres claire. En effet, Mooney et al. (1978), Field et
Mooney (1983), Oberbauer et al. (1985) et Walters et al. (1993a) ont constate que la VSC est
correlee positivement avec la PN exprimee par unite de surface foliaire (PNs). Des resultats
(1990) quand il ont mesure la PN par rapport a la biomasse foliaire (PNm). En revanche,
d'autres auteurs n'ont pas obtenu une correlation entre la VSC et la PNs quand ils ont compare des especes ayant des VSC differentes mais possedant des morphologies semblables, comme c'est Ie cas des especes herbacees (Dijkstra et Lambers, 1989 ; Poorter et Remkes, 1990). Cependant, les especes cultivees et les 'mauvaises herbes' qui les accompagnent, et qui possedent des VS C elevees, ont un taux de PNs plus eleve que celui des arbustes et les arbres a feuilles persistantes, qui eux possedent des VSC plus faibles (Evans, 1983 ; Field et Mooney, 1983 ; Langenheim et al., 1984 ; Sage et Pearcy, 1987 ; Makino et al., 1988). Aussi, par rapport aux especes d'ombre qui ont une VSC faible, les especes de lumiere, qui ont des VSC elevees, ont des niveaux de saturation de PNs plus eleves (Pons, 1977 ; Bjorkman, 1981 ; Seemann et al., 1987) et contiennent moins de chlorophylles (Poorter et al., 1990). Cet investissement dans la chlorophylle augmente la capacite d'assimilation dans les conditions d'ombre (Evans, 1989a). L'absence de consensus entre les chercheurs sur Ie role de la PN dans
la determination de la VSC serait due selon Zelitch (1982) au fait que les mesures de PN ne
sont effectuees qu'instantanement. De plus, ces mesures sont realisees seulement sur quelques feuilles au lieu de considerer toute la plante. Enfin, les mesures sont rarement prises dans un milieu naturel et pendant toute la duree de vie de la plante.
La difference dans les taux de PN entre les especes peut aussi s'expliquer par la difference de la concentration intracellulaire en CO^ (Ci), la respiration et la conductance des stomates (CS). En effet, tout changement dans la CS affecte a la fois la PN et la Ci car c'est a travers les stomates que s'effectuent la majorite des echanges gazeux entre la plante et 1'atmosphere (Farquhar et Sharkey, 1982). En fonction des conditions environnementales, les especes vegetales ont developpe differentes strategies pour garder une bonne efficacite dans Futilisation de 1'eau sans compromettre leur assimilation photosynthetique.
Enfm, la variation de la nutrition azotee peut affecter la PN (Evans, 1983 ; Field et Mooney,
Reich et al., 1994). Toutefois, Gamier (1991) a observe une correlation positive entre la PN et la concentration de 1'azote uniquement chez les monocotyledones. Aussi, 1'investissement de 1'azote dans les composes secondaires de defense centre les herbivores affecte la concentration des enzymes qui interviennent dans la PN, et par consequent, la croissance des plantes est
affectee (Coley, 1986).
3.2 Respiration
Environ 50% des photosynthetates produits quotidiennement sont perdus par la respiration (mitochondriale et photorespiration) des differents organes des plantes (Lambers, 1985). Cependant, les especes qui ont une VSC elevee ont des taux de respiration plus eleves que ceux des especes qui ont des VSC faibles (Dijkstra et Lambers, 1989 ; Poorter et al., 1990 et
1991 ; Walters et al., 1993b). L'effet de la lumiere sur les variations des taux de respiration n'est pas toujours clair. En effet, les feuilles des plantes tolerantes a 1'ombre qui ont des VSC faibles, possedent des taux de respiration plus faibles que ceux des feuilles des plantes de lumiere, qui par centre ont des VSC elevees (Pons, 1977). Toutefois, Kuiper et Smid (1985) ont observe que les especes cultivees sous fort eclairement ont des taux de respiration tres proches de ceux des especes cultivees sous faible eclairement. Mais, quand les plantes sont transferees d'un milieu bien eclaire vers un milieu faiblement eclaire, Ie taux de respiration diminue. Done, Ie role de la respiration dans Ie controle de la croissance des plantes reste mal connu et souvent mal estime. Ceci pourrait etre due a la photorespiration (au moins chez les especes €3) et les problemes techniques pour mesurer la respiration des racines.
3.3 Allocation de biomasse
Les parametres d'allocation utilises par les differents chercheurs sont souvent des rapports de biomasse des feuilles, des tiges et des racines qui representent respectivement les rapports de biomasse allouee aux feuilles, aux tiges et aux racines par rapport a la biomasse totale. Les
plantes montrent une grande plasticite dans leur maniere d'allouer leur biomasse (Wilson,
1988 ; Berendse et Elberse, 1989 ; Causton et Dale, 1990 ; Grime et Campbell, 1991).
Toutefois, les especes qui ont une croissance rapide montrent plus de plasticite morphologique que les especes qui ont une croissance lente quand ces especes sont cultivees a differents niveaux de richesse en azote (Robinson et Rorison, 1987). Cependant ce resultat n'est pas general (Olff et al., 1990). Les conditions environnementales peuvent influencer la maniere dont les plantes allouent leur biomasse. En effet, un eclairement fort pendant la croissance des plantes provoque plutot une allocation importante vers les feuilles ou les tiges, tandis que 1'ombre reduit la VSC, et surtout 1'investissement a court tenne dans les racines (Dale et Causton, 1992). Dans les milieux pauvres en elements nutritifs, les plantes allouent plus de biomasse vers les racines que vers les parties aeriennes (Chapin, 1980 ; Lambers et Dijkstra,
1987 ; Tilman et Cowan, 1989 ; Shipley et Peters, 1990b ; Fichtner et Schulze, 1992 ; Lambers
et Poorter, 1992). Ceci se traduit par des RBF faibles et des RBR eleves. Toutefois, d'autres chercheurs ont observe que les especes originaires des milieux pauvres investissent plus de biomasse dans leurs tissus aeriens que dans leurs racines quand la disponibilite de mineraux et de lumiere est gardee constante (Olffet al., 1990). Par contre, Garnier (1991) a trouve que les monocotyledones investissent en general plus de biomasse dans les racines que les dicotyledones. Get auteur a aussi observe que la VSC n'est pas liee a la concentration d'azote dans les plantes ou au type d'allocation de biomasse, mais plutot au produit de ces deux attributs.
La relation entre la VSC et Ie RBF depend des conditions environnementales. Ainsi, chez des especes herbacees. Hunt et al. (1987) ont obtenu une correlation negative sous eclairement faible, alors que Shipley et Peters (1990b) ont obtenu Ie meme resultat sous des eclairements plus forts. Ces deux resultats vont dans Ie sens contraire de la theorie de Tilman (1988) decrite precedemment. En revanche, d'autres chercheurs ont observe une correlation positive a des niveaux d'eclairement intennediaire (Ingestad, 1981 ; Poorter et Remkes, 1990). Toutefois, ces deux derniers auteurs ont observe que la SSF etait plus correlee avec la VSC que Ie RBF.
Enfm, Walters et al., (1993b) ont observe une forte correlation positive entre la VSC et 1'allocation de biomasse vers les feuilles et les tiges chez desjeunes arbres.
Les especes qui colonisent les milieux riches en ressources minerales et qui ont une VSC elevee ont une plus grande capacite a utiliser les ressources que les especes qui ont une VSC faible. Ceci peut expliquer 1'absence des especes qui ont une croissance rapide dans les milieux pauvres car ces especes tendent a garder une forte translocation de ressources vers les feuilles malgre leur faible disponibilite de ces ressources. Ceci provoquerait un appauvrissement rapide des sols, et done la disparition de ces especes de ces milieux infertiles (Berendse et Elberse, 1989).
II semble done que la difference des VSC chez les especes vegetales est liee plutot a 1'activite des differents organes qu'au type d'allocation (Garnier, 1991). Selon Rodgers et Bameix (1988) et Lambers et Poorter (1992), cette difference des VSC serait une consequence plutot qu'une cause des differences dans la capacite a utiliser les ressources. Puisque les conditions environnementales jouent un role important dans 1'allocation de la biomasse chez les plantes (Poorter et Lambers, 1991), nous avons decide de travailler dans des conditions de nutrition minerale et d'eclairement differentes.
3.4 IVIorphologie des feuilles
La SSF est un tres bon attribut pour decrire la morphologie des feuilles car cet attribut tient compte a la fois de la surface des feuilles et du taux de biomasse investie par unite de surface. En general, les especes qui colonisent les milieux riches en elements nutritifs ont des SSF plus elevees que celles des especes originaires des milieux pauvres (Sage et Pearcy, 1987 ; Poorter, 1989 ; Elberse et Berendse, 1993). En outre, la faible SSF des plantes qui colonisent les milieux infertiles s'accompagne toujours d'une longevite elevee (Coley, 1988 ; Reich et al.,
milieux infertiles investissent plus de biomasse dans leurs racines que dans leurs feuilles car la competition dans ces milieux se fait pour les elements nutritifs du sol et non pour la lumiere.
La composition chimique des feuilles peut etre a Forigine de ces variations interspecifiques de la SSF. En effet, les especes qui ont une croissance lente investissent relativement plus dans les composes de defense contre les herbivores, comme les tanins, la lignine et les composes secondaires, que les especes qui ont une croissance rapide (Bryant ei al., 1983 ; Coley, 1983, 1986 et 1987 ; Merino et al., 1984 ; Waring et al., 1985). Aussi, les especes qui ont une VSC faible contiennent plus de materiel dans leurs parois cellulaires, comme la lignine et 1'hemicellulose (Dijkstra et Lambers, 1989b) que les especes ayant une VSC elevee qui, elles sont plus riches en elements cytoplasmiques, comme les proteines (Dijkstra et Lambers, 1989 ; Niemann et al., 1992). La question de la relation entre la richesse en composes de defense et la croissance reste a elucider. Jusqu'a maintenant, peu de chercheurs ont quantifie 1'impact de 1'investissement des plantes dans ces composes sur leur potentiel de croissance (Coley, 1986 ; Baas, 1989 ; Lambers et Rychter, 1989 ; Almeida-Cortez , 1997). Coley (1986) a trouve une correlation negative entre la production des feuilles et leur concentration en taimin chez Cecropia peltata. En revanche, McCanny et al. (1990) et Almeida-Cortez (1997) n'a trouve aucune correlation apparente entre la toxicite de 28 especes herbacees pour une espece de daphne (test biologique) et la VSC de ces especes. II semble done que 1'accumulation des composees secondaires chez les especes qui ont une croissance lente serait un resultat phenotypique exactement comme la longevite des feuilles et leur sclerophyllie.
L'epaisseur des differents tissus du limbe peut aussi etre a 1'origine des differences de SSF observees entre les especes qui occupent des milieux differents (Shipley, 1995). En effet, les especes de lumiere qui ont une VSC elevee ont des limbes plus epais que les especes d'ombre
qui ont une VSC plus faible (Young et Smith, 1980 ; Bjorkman, 1981). Cette difference est
associee aussi a 1'epaisseur des tissus assimilateurs (Pons, 1977 ; Bjorkman, 1981 ; van Arendonk et Poorter, 1994 ; Garnier et Laurent, 1994), a 1'epaisseur des tissus de soutien
(Baruch et al., 1985 ; Pammenter et al., 1986 ; Garnier et Laurent, 1994), ou aux variations de
la taille de la nemire centrale (Shipley, 1995).
La relation entre Fepaisseur du limbe et la SSF n'est pas toujours significative (Dijkstra et
Lambers, 1989 ; Witkowski et Lamont, 1991 ; Kebede et al., 1994 ; Gamier et Laurent, 1994 ;
van Arendonk et Poorter, 1994). Par ailleurs, selon van Arendonk et Poorter (1994), la variation de la SSF et de la VSC chez 14 especes herbacees ne presente pas de lien avec les epaisseurs du mesophylle ou des nervures centrales. Par contre, 1'epaisseur du sclerenchyme et de 1'epiderme est correlee negativement avec la VSC. Gamier et Laurent (1994) n'ont pas trouve de differences interspecifiques au niveau de 1'epiderme. Ces resultatscontradictoires nous ont amenes a chercher d'abord les sources de variations interspecifiques des SSF, en cherchant dans un premier temps les liens de cet attribut avec les epaisseurs des limbes et celles des nervures centrales et Ie pourcentage d'eau dans les feuilles. Puis, dans un deuxieme temps, nous avons cherche les liens entre ces variables et la masse seche des feuilles et leur surface. Enfin, nous avons cherche la relation entre la VSC et les differents tissus foliaires en fonction de la richesse du milieu en elements nutritifs et de lumiere.
Les plantes sont des ensembles d'organes dont Ie fonctionnement est complexe. Ces organes ont souvent des demandes en ressources differentes et quelquefois conflictuelles. Bien que tout Ie monde s'accorde pour dire que les attributs morphologiques, physiologiques et d'allocation interagissent entre eux, tres peu de chercheurs ont fait la synthese de toutes les interactions pour produire des modeles mathematiques de prediction. La difficulte vient du fait que meme dans des conditions controlees, nous ne pouvons pas controler toutes les variables comme la morphologie et les echanges gazeux. D'ou la necessite d'utiliser un outil statistique qui permet d'avoir un controle sur ces variables. La technique d'analyse de pistes est un bon choix pour realiser cette tache. C'est pour cela que nous 1'avons utilisee dans cette etude afm de developper d'abord des modeles de prediction en decomposant chacun des attributs, pour ensuite en arriver a des modeles generaux de prediction incluant la VSC.
Cette technique a ete invente par Ie geneticien et biometricien Sewall Wright (1920, 1921).
Malheureusement, ce biologiste eminent n'a jamais reussi a imposer cette technique dans Ie
domaine de la biologie. Selon Shipley (1998), la raison de son echec est que les biologistes de
1'epoque avaient plus de confiance dans les methodes de Fisher, un autre biometricien contemporain, qui paraissaient plus rigoureuses a cause des permutations des blocs experimentaux. Par ailleurs, la methode originale de Wright n'offrait pas la possibilite de tester les modeles batis ce qui faisait du developpement des modeles une pure speculation. Ce n'est que pendant les annees 60 que des chercheurs des sciences sociales ont redecouvert les potentialites de 1'analyse des pistes ainsi que celui des correlations partielles qui permettent d'avoir un controle statistique sur les variables non mesurees. Ces chercheurs ont par la suite ameliore cette methode en y ajoutant une statistique (Khi carre de maximisation de vraisemblance) avec une distribution de probabilite connue (Bollen, 1989) qui nous permet de tester et potentiellement de refuter nos modeles. Par la suite, les chercheurs des sciences politiques et des autres sciences sociales ont adopte cette technique. L'apparition des logiciels a contribue a augmenter Ie nombre des utilisateurs grace a la simplification de son utilisation,
par exemple LISREL (Joreskog et Sorbom's, 1986) et EQS par Bentler (1995).
Ironie du sort, bien que 1'inventeur de cette technique fat un biologiste, c'est dans Ie domaine de la biologie que cette methode a eu Ie moins de succes. Toutefois, de plus en plus de chercheurs commencent a 1'utiliser (Sokal et Rohlf, 1995). Malheureusement, lorsqu'elle est mal utilisee comme c'est Ie cas dans 1'etude de Smith et al. (1997), elle donne des resultats incorrects, toute comme n'importe quelle autre methode. En effet, dans leur etude. Smith et al. n'ont meme pas pu teste leur modele car Us n'ont pas eu assez de degre de liberte.
4. Objectifs
Comme il a ete mentionne plus haut, la VSC des especes vegetales depend a la fois de leur morphologie, de leur physiologie et de la maniere dont elles allouent leur biomasse. Le
premier objectif de ce travail etait done d'une part de determiner 1'importance de chacun de ces attributs dans la determination de la VSC. D'autre part, puisque les conditions environnementales joueirt un role tres important dans Ie comportement de chacun des attributs qui determinent la VSC (Lambers et Poorter, 1992 ; Walters et al., 1993b), nous avons teste cette hypothese en travaillant dans des conditions de nutrition minerale et d'eclairement differentes.
Le deuxieme objectif etait de decomposer les differents attributs (VAN, SSF) pour chercher
les sources de variation a 1'interieur de chaque attribut. Ainsi, pour 1'attribut physiologique, au lieu de ne considerer que la VAN comme seul attribut physiologique, des mesures d'echange gazeux ont ete effectuees pour mieux cemer la reponse physiologique des differentes especes (PN, CS et Ci). En ce qui concerne la composante morphologique, des mesures des surfaces foliaires et des epaisseurs des differents tissus des feuilles ont ete effectuees afiin de mieux expliquer les variations de la SSF ainsi que Ie comptage des stomates sur les deux faces des feuilles. Enfm, pour 1'allocation de biomasse, des mesures de poids sees des differentes parties des plantes (racines, tiges et feuilles) en fonction du temps ont ete realisees. Ces mesures nous renseignent sur Ie mode d'allocation de la biomasse produite en relation avec la VSC.
II est clair que les differents attributs interagissent continuellement entre eux. En effet, Pinfluence d'un attribut sur la VSC peut etre directe ou indirecte par 1'intermediaire d'autres attributs. C'est pour cela que notre demier objectif etait d'analyser ces interactions et les representer sous forme de modeles mathematiques de prediction pour decrire ces liens directs et indirects. Pour cela, la methode statistique d'analyse de pistes (Path analysis) a ete utilisee.
CHAPITRE 1
MATERIELS ET METHODES
1.1 Choix des especes
Pour realiser ce projet. Ie choix a ete porte sur 22 especes vasculaires d'angiospermes. De ces especes herbacees sauvages, 13 appartiennent a la classe des Magnoliopsidees (dicotyledones) et 9 a la classe des Liliopsidees (monocotyledones). Ces 22 especes comprenaient 8 ordres, 9 families et 21 genres. Le tableau 1 montre la classification taxinomique de chaque espece. Mis a part Panicum capillare qui appartient au groupe des plantes de type €4, toutes les autres especes sont de type €3.
1.2 Recolte des graines et germination
Les graines ont ete ramassees dans les champs ouverts des environs de la ville de Sherbrooke. Ces especes sont typiques du sud ouest de la province du Quebec. Ensuite, les graines ont ete sechees a la temperature ambiante du laboratoire puis stockees a froid a environ 4°C jusqu'au jour de la mise a germination. A ce moment, les graines ont ete disposees dans des boites de petri en presence d'eau distillee. La germination des graines s'est deroulee a 1'interieur de la chambre de croissance. Par ailleurs, 1'imbibition des graines avec la giberelline ne s'est pas averee necessaire pour enlever la dormance, d'autant plus que cette hormone peut activer la croissance pendant les premiers stades.
Une fois que les graines ont commence a sortir leur radicule, elles ont ete transplantees dans des pots quadrangulaires en papier plastifie d'une capacite de 1300 cm3, remplis avec du sable siliceux tres fm (gram<lmm). Ainsi Ie lessivage etait moins rapide. Deux series de 3 trous ont
Tableau 1. Liste des 22 especes utilisees et leur affiliation taxinomique. Classes Ordres Magnoliopsidees Asterales Plantaginales
Caryophyllales
Polygonales
Myrtales LaminalesLiliopsidees Cyperales
Graminales AralesFamilies
Asteracees Plantaginacees Caryophyllacees Polygonacees Onagracees Brassicassees Lamiacees Cyperacees Poacees Aracees Genres Chrysanthemum Cichorium Eupatorium Hieracium Leontodon Plantago Plantago Silene Polygonum Rumex Oenothera Erysimum Prunella Carex Agropyron Bromus Deschampsia Hordeum PanicumPhleum
Poa Acorus Especes leucanthemum L. intybus L. maculatum L. aurantiacum L. autumnalis L. lanceolata L. major L. cucubalus Wibel.lapathifolium L.
acetosa L. biennis L. cheirantoides L. vulgaris L. crinita Lam. r opens (L.) Beauv. inermis Leyss. cespitosa (L.) Beauv. jubatum L. capillare L. pratense L. pratensis L. calamus L.ete realisee sur chaque face des pots et ce, sur deux niveaux; Ie premier niveau a la base des pots et Ie second a 3 cm de la surface. Deux a trois plantules ont ete plantees dans chaque pot. Mais une fois que les phntules ont ete bien etablies, un seul individu choisi au hasard a ete maintenu par pot pour eviter les problemes de carence en element nutritifs et 1'enchevetrement des racines.
1.3 Conditions de culture
L'etude s'est deroulee sous quatre traitements (details plus bas), deux niveaux de concentration des mineraux nutritifs (N n) combines avec deux intensites d'eclairement (L 1). Dans chaque traitement, 22 especes ont ete utilisees et chaque espece etait representee par 15 individus. Done, un total de 1320 plantes ont ete utilisees pour realiser ce travail.
Les cultures se sont deroulees dans une chambre de croissance (Conviron CMP 3244) ou 1'intensite cTeclairement, la temperature et 1'humidite relative etaient controlees. La photoperiode etait fixee a 15h et 9h pour les durees de jour et de nuit. La temperature du jour etait fixee a 25°C et celle de la nuit a 15°C. Enfm, 1'humidite relative de 1'air ambiant variait entre 80 et 90% selon les periodes d'arrosage.
Comme 1'espace a 1'interieur de la chambre de croissance n'etait pas suffisant pour mettre les individus des 22 especes en meme temps, seulement 11 especes ont ete utilisees a la fois. Done chaque traitement etait etale sur deux periodes de 35 jours. Pendant chaque experience, 15 individus par espece ont ete utilises pour assurer 5 recoltes de 3 individus par espece aux jours
15, 20, 25, 30 et 35 apres la germination. Done, la chambre de croissance comprenait 165 individus a la fois.
Pour que les recoltes soient faites d'une fa9on aleatoire, chaque individu des differentes especes a d'abord ete place dans un pot a part, en gardant un espace suffisant entre les pots
atm d'eviter 1'ombrage (7cm entre les plantes voisines). Puis, les 15 individus des 11 especes out ete repartis aleatoirement sur 15 blocs dans une bassine en plastique inerte de 150 cm de longueur, 69 cm de largeur et 26 cm de hauteur. La bassine a ete placee a 1'interieur de la chambre de croissance. De cette maniere, 1'arrosage se faisait automatiquement a 1'aide d'une pompe qui remplissait la bassine avec la solution nutritive et une autre qui la vidait apres une duree de 30 minutes. L'operation etait repetee 3 fois par 24 heures. Toutes ces operations automatiques d'arrosage et de vidange etaient controlees par des minuteries. II faut aussi noter que la bassine ou les pots etaient disposes a ete soulevee de 80 cm pour la rapprocher des lampes, augmentant ainsi 1'intensite d'eclairement. Malheureusement, la temperature a aussi augmente pendant Ie jour a 28°C au niveau des plantes. Pour remedier a ce probleme, un ventilateur a ete installe pendant les quatre traitements dans la chambre de croissance pour ameliorer la circulation d'air et done homogeneiser 1'humidite relative et la temperature a 1'interieure de la chambre. Ainsi, la temperature au niveau des plantes a baisse a 25-27°C pendant Ie jour. Par centre, la temperature noctume est restee stable a environ 15°C. Le ventilateur etait place sous la bassine pour eviter 1'exposition directe des plantes a la ventilation, ce qui aurait affecte les echanges gazeux des plantes.
Les intensites d'eclairement pour simuler Ie milieu ouvert bien ensoleille (L) et Ie milieu ombrage (1) etaient fixees respectivement a 1100±100 jLimoles/m/s et a 200±50 |Limoles/m2/s. Pour obtenir la faible intensite d'eclairement, nous avons place des filtres gris entre les lampes et les plantes. De cette fa^on, 1'intensite de lumiere a baisse mais non la qualite du spectre de reflectance. (figures 1 et 2). Ces spectres de reflectance sont obtenus avec un spectrometre de type Kodak (Personnal spectrometer II). Les eclairements etaient assures par 5 lampes au sodium a haute pression et 5 lampes a halogenure de metal. Chaque lampe avait une puissance de 400 watts. La combinaison de ces deux intensites d'eclairement avec deux concentrations d'elements nutritifs, une forte (N) et 1'autre faible (n) , a donne les quatre traitements sous lesquels les plantes ont ete cultivees pour realiser cette etude a savoir LN, IN, 1 netL n.
1.4 Solution nutritive
La composition chimique de la solution nutritive a etc determinee apres la consultation des travaux de plusieurs chercheurs. Smith et al. (1983) donnent une comparaison entre les solutions utilisees par differents auteurs. La solution presentee dans Ie tableau 2 a ete choisie pour simuler un milieu riche en elements nutritifs, alors que sa dilution six fois, c'est-a-dire la division de la concentration de tous elements par six, a ete utilisee pour simuler Ie milieu pauvre. Les solutions meres (1 M) de chaque macroelement ont ete preparees separement dans des contenants a part, alors que la solution contenant les microelements (a part Ie fer) a ete preparee en melangeant les volumes des solutions de stock comme indique dans Ie tableau 2 dans un volume final toujours de 1 litre.
Pour preparer la solution riche en elements mineraux, nous avons pris 100 ml de cette solution de microelements et 1'avons melangee dans Ie volume final de 100 1, tandis que pour preparer la solution pauvre en elements mineraux, nous n'avons pris que 16,66 ml de cette solution et 1'avons melange dans Ie volume final de 100 1.
Pour preparer la solution de fer, nous avons melange 7,5 mg d'EDTA dans un volume proche de 1 litre. Afin de faciliter la dissolution de 1'EDTA, nous avons ajoute du NaOH pour baisser Facidite. Ensuite, nous avons ajoute 5 g de FeS047H20 et complete jusqu'a un volume final de 1 litre. Pour la solution riche en elements nutritifs, nous avons mis 150 ml de FeEDTA dans 100 1. Par contre, pour la solution pauvre en elements nutritifs, nous avons melange 25 ml de FeEDTA avec 100 1 de solution finale. En ce qui conceme Papport d'azote, 5 et 0,833 mmoles etaient fournies sous forme de nitrate et 1 et 0,167 mmoles sous forme d'ammonium respectivement pour les milieux riches et pauvres.
s» n- rs as —• -( re
3
re B <^- 1-?S G' 1 orq "ff -1 fl? 1-* (/) la ffi 0ft-3
c- w "1 ft'tt
? n as 3 0 ftNombres digitaux
01 0 0 0 0 0 01 0 0 M 0 0 0 ro 01 0 0 oa 0 0 0 fc» 01 0 0 *. 0 0 0rt>< -( ? c -{ &- ft (0 0
y
» cr ft e- ft rs -( c un aa ?fl B n ft T3 ft D&
&s S3era ^ro t^>
Nombres digitaux
.^v
TO rs <-»-h"»s as' -{ n G. ft ft'a
ft n 0- ;4 n 63 B n ft n" -t re e I CL re as rs S3- 6S5
a- ft CL ft 0 0 v>. 3 0 ft)v
n> B 0- ^ S3r
0 B era ft 1 0- ^ 0 s CL ft ~9B
Done, Ie milieu riche contenait 6 mmoles d'azote et Ie milieu pauvre en contenait seulement 1 mmole. Deux raisons majeures nous ont pousses a utiliser 1'ammonium en plus du nitrate comme source d'azote. D'abord parce que la presence de 1'ammonium dans la solution nutritive reduit les variations du pH, car 1'absorption de 1'azote sous forme anionique augmente Ie pH, alors qu'une absorption d'azote sous forme cationique baisse Ie pH. Done Ie melange des deux formes d'azote s'avere tres utile pour stabiliser Ie pH de la solution nutritive. L'autre raison, est que la presence d'ammonium faciliterait 1'absorption des nitrates (Asher et Edwards, 1973 ; Smith et al., 1983). Enfm, malgre toutes ces precautions, Ie pH de la solution etait controle constamment (chaque 2 jours) et ajuste avec du NaOH ou du N2804 pour Ie garder proche de 5,8.
1.5 Mode de recolte et de mesure des differents attributs
Les recoltes de 3 individus par espece ont ete effectuees lesjours 15, 20, 25,30 et 35 apres la germination. A chaque recolte, les differentes parties des plantes (racines, tiges, feuilles et fleurs) ont ete separees, (les racines sont lavees et essuyees) et pesees avec une balance de precision pour prendre Ie poids frais. Ensuite, la surface de feuilles a ete mesuree avec un analyseur d'image (Agvision, 1992), compose d'une camera et un ordinateur muni d'un programme special pour calculer les surface des feuilles a partir de 1'image de la camera. Enfm, les differentes parties ont ete sechees separement a 70 °C pendant au moins 36 heures avajit de prendre leur poids sec qui a servi dans Ie calcul des VSC. Pour cette demiere fin, nous avons d'abord transforme les poids sees des individus recoltes pendant les 5 periodes de temps en leur logarithme neperien. Puis, nous avons calcule la valeur moyenne de la VS C qui est egale a la valeur de la pente de 1'equation de regression suivante :
ln(B)=a+b(t)+erreur
Tableau 2. Composition chimique de la solution nutritive riche en elements
nutritifs (N)
Macroelements(PM)
KN03 (101,1)
Ca(N03)24H20 (236)
MgS047H20 (246)
KH2P04(136)
(NH4)2S04(100)
Microelements (PM)
MnS04(151)
ZnS047H20 (287)
H3B03 (62)
Na^McAHp (224)
CuS04 (159,5)
FeEDTA(348)
mg/1
N=28 K=78,2 N=42 Ca=60 Mg=49 S=64,5 K=39,l P=31 N-14 S=31 Mn=0,5 S=0,3 Zn=0,05 S=0,023 B=0,5 Na=0,002 Mo=0,01 Cu=0,02 8=0,012 Fe=2 Somme totale(mg/1)
N=84
P=31
K=117
Mg=49
S=96
Mn=0,5 Zn=0,05 B=0,5 Mo=0,01 Cu=0,02 Fe=2 ml solution 1Mdans 1001
200
150
201
100
50
Dans 100 ml
137
21
287
2
6
150
Les mesures des echanges gazeux (PN, CS et Ci) ont ete effectuees seulement au 34 jour apres la germination a 1'aide d'un analyseur a infrarouge de gaz carbonique de marque Li-Cor (Li-6200). Cet appareil mesure la difference de la concentration de 002 et de la vapeur d'eau dans F air avant et apres passage dans la chambre de mesure qui contient la feuille sur laquelle les mesures d'echanges gazeux vont s'effectuer. La chambre de mesure contient une electrode pour mesurer la temperature de la feuille. Pour realiser les mesures d'echanges gazeux, deux feuilles de rang intermediaire de chacun des 3 individus de chaque espece ont etc choisies au hasard pour prendre une mesure par feuille. Lorsque les feuilles n'etaient pas assez larges ou 1'intensite d'eclairement etait faible, plusieurs feuilles ont send a prendre les mesures des
surface et leur biomasse separement. Pour 1'eclairement pendant les mesures, il etait Ie meme que celui de la culture.
Conductance des stomates (CS)
Le principe de mesure de la CS est basee sur les modeles de Von Caemmerer et Farquhar (1981) qui stipulent que ce parametre peut etre represente sous la forme suivante (Li-6200,
1990):
CSt = E*[l-(Wi+We)/2]/W;-We
Aussi, la conductance totale des stomates peut etre representee sous forme de deux composantes :
l/CS,=l/CS+l/g,,
CS = l/[(e,*Tre)/E(P-(e,*T,+e)/2) - (K2+l)/g^*(K+l)2]
CSt: Conductance totale de la vapeur d'eau Tf: Temperature foliaire
E : Taux de transpiration
w,: Fraction molaire de vapeur d'eau intracellulaire Wg: Fraction molaire de vapeur d'eau extracellulaire es : Pression de vapeur d'eau saturante
ei: Pression de vapeur d'eau extracellulaire P : Pression atmospherique
CS : Conductance stomatique pour la vapeur d'eau
g^ : Conductance de la couche limite pour la vapeur d'eau
moles c°
H
mmoles e,/P e,/Pmb
mb
mb
moles molesH
H
zO/m2/
7s
H20/m2/sP/m2/
P/m2/
!/sVs
;clo K:
Conductance de la couche limite pour une face foliaire moles H20/m /s Rapport de stomates
K=l si les stomates se trouvent sur une seule face du limbe K=0,5 si les stomates se trouvent sur les deux faces du limbe
Photosynthese nette (PN)
Le taux de photosynthese nette est calcule a 1'aide de la formule suivante :
PN = [F/e*C,/100SISS*P] - [P*G'tiV/8,314(T,+273)*S) * (dC/dt)] - C*E*G
Sachant que
G=(l-W2)/(l-W2+fliW2)
PN : Taux de photosynthese nette F(} : Flux d'air traversant Ie dessechant
e : Pression de vapeur d'eau dans la chambre de mesure Cg: Concentration de €03 dans 1'air ambiant
S : Surface foliaire
P : Pression atmospherique
V(: Volume total de la chambre de mesure Ta: Temperature ambiante
dC/dt: Taux de changement de CO^ C : Concentration de CO^
E : Taux de transpiration foliaire
f: Fraction d'air traversant Ie dessechant F^: Flux d'air maximal
W2: Fraction de vapeur d'eau
(J-moles C02/m2/s p-moles/s