HAL Id: hal-02597031
https://hal.inrae.fr/hal-02597031
Submitted on 15 May 2020HAL is a multi-disciplinary open access
archive for the deposit and dissemination of sci-entific research documents, whether they are pub-lished or not. The documents may come from teaching and research institutions in France or abroad, or from public or private research centers.
L’archive ouverte pluridisciplinaire HAL, est destinée au dépôt et à la diffusion de documents scientifiques de niveau recherche, publiés ou non, émanant des établissements d’enseignement et de recherche français ou étrangers, des laboratoires publics ou privés.
Une méthode hiérarchique pour le diagnostic de la
biodiversité forestière dans les espaces protégés : étude
de cas dans le Vercors Isérois
M. Redon
To cite this version:
M. Redon. Une méthode hiérarchique pour le diagnostic de la biodiversité forestière dans les espaces protégés : étude de cas dans le Vercors Isérois. [Rapport de recherche] irstea. 2012, pp.40. �hal-02597031�
1
Une méthode hiérarchique pour le diagnostic
de la biodiversité forestière dans les espaces
protégés : étude de cas dans le Vercors Isérois
Rapport scientifique
Étude réalisée par Mathilde REDON
11
Irstea/Cemagref centre de Grenoble, Unité de recherche Ecosystèmes Montagnards, 2, rue de la papeterie 38402 Saint-Martin d’Hères
Avec la participation du Conseil Général de l’Isère dans le cadre du
Pôle départemental de recherche sur la biodiversité en Isère – 2011
CemOA
: archive
ouverte
d'Irstea
2
S
OMMAIRE
1
C
ONTEXTE ET OBJECTIF DE L’
ÉTUDE... 3
1.1
Enjeux autour de la préservation des écosystèmes forestiers ...3
1.2
Mécanismes associés à la préservation de la biodiversité ...3
1.3
Objectif ...5
2
S
ITES D’
ÉTUDE... 5
3
C
ONSTRUCTION DE LA GRILLE DE NOTATION... 7
3.1
Principe du diagnostic de la biodiversité potentielle ...7
3.2
Quelques notions d’écologie du paysage et d’écologie forestière ...10
3.3
Supports cartographiques ...11
3.4
Justification et calcul des critères et indicateurs utilisés dans la grille de
notation ……….14
3.4.1 Choix des critères et indicateurs ... 14
3.4.2 Milieux forestiers ... 14
3.4.3 Milieux forestiers publics ... 17
3.4.4 Interface forêt/non-forêt ... 19
3.4.5 Milieux non forestiers ... 20
4
R
ÉSULTATS... 22
4.1
Comparaison des ENS et des RBIs ...23
4.1.1 Comparaison des RBIs entre elles ... 23
4.1.2 Comparaison des ENS entre eux ... 24
4.1.3 Comparaison entre les modalités gérées (ENS) et non-gérées (RBIs) ... 24
4.1.4 Interprétation des résultats ... 25
4.1.5 Conséquences en termes de biodiversité potentielle dans les modalités gérées et non gérées ………28
4.2
Comparaison des potentialités dans les sites d’Engins et des Ecouges ...28
5
D
ISCUSSION GÉNÉRALE... 30
6
B
IBLIOGRAPHIE... 32
7
T
ABLES DES ILLUSTRATIONS... 35
7.1
Figures ...35
7.2
Tableaux ...35
8
A
NNEXES... 37
CemOA : archive ouverte d'Irstea / Cemagref3
1 C
ONTEXTE ET OBJECTIF DE L’
ÉTUDE1.1
Enjeux autour de la préservation des écosystèmes forestiers
L’accroissement des pressions sur les ressources naturelles au cours des dernières décennies a conduit à un déclin de la biodiversité1 à un rythme sans précédent (Butchart et al. 2010). Les écosystèmes forestiers comptent parmi les plus affectés par les changements d’occupation des sols et l’artificialisation des habitats naturels résultant de ces pressions (EEA 2010). Or, ces écosystèmes fournissent de nombreux services indispensables au bien-être et au bon fonctionnement des sociétés humaines, tels que la production d’oxygène, le stockage du carbone, la fertilisation des sols, l’épuration de l’eau, la production de ressources alimentaires variées (baies, champignons…), de matériaux de construction et de chauffage (bois), des zones de récréation ainsi qu’une atténuation des risques naturels (ex. chutes de blocs rocheux) (TEEB 2010; EU 2011). La réalisation de ces services est étroitement liée à la biodiversité : des décomposeurs aux prédateurs, des myriades d’espèces souterraines et aériennes contribuent au recyclage de la matière organique, à l’absorption des nutriments par les arbres, à la dispersion des graines, à la régulation des populations ou à la lutte contre les parasites et maladies (André 2005; Dajoz 2007). Maintenir la biodiversité forestière représente alors un enjeu important pour le fonctionnement des écosystèmes forestiers à long terme, condition indispensable pour assurer la pérennité des services marchands et non marchands qui en dépendent (TEEB 2010).
1.2
Mécanismes associés à la préservation de la biodiversité
La conférence de Rio sur la Conservation de la Diversité Biologique (CBD) en 1992 a marqué un premier pas vers l’institutionnalisation du concept de biodiversité, contribuant à l’accélération du développement d’outils d’évaluation, de suivi et de protection de la biodiversité au niveau international (Levrel 2007). En Europe, c’est le processus paneuropéen de la Conférence Ministérielle pour la Protection des Forêts en Europe (CMPFE)2, dit processus d’Helsinki, qui a pris le relai de la CBD. Il a notamment contribué au développement du concept de gestion forestière durable et initié des réflexions sur la définition de critères et d’indicateurs permettant de suivre indirectement l’évolution de l’état de conservation des forêts et de leur biodiversité (MCPFE 2007).
1
La biodiversité peut être définie comme la diversité des organismes vivants quelle que soit leur origine, incluant la diversité des processus et des fonctions qui permettent leur survie. Elle inclut la diversité des gènes, des espèces et des écosystèmes (adapté de CBD (2002)).
2
Aujourd’hui Forest Europe
CemOA
: archive
ouverte
d'Irstea
4 Dans le même temps, plusieurs Directives européennes ciblant plus directement la conservation des espèces et des habitats naturels, incluant les espèces et habitats forestiers, ont été successivement mises en œuvre. Les Directives dites « Habitats-Faune-Flore » (92/43/CEE, du 21 mai 1992) et « Oiseaux » (79/409/CEE, du 2 avril 1979), en particulier, sont à l’origine de la création du réseau paneuropéen Natura 2000, dont l’objectif est d’assurer la préservation cohérente des habitats naturels ainsi que de la faune et de la flore sauvage à l’échelle de l’Europe. En France, comme dans de nombreux pays, le réseau Natura 2000 est intégré dans un réseau plus vaste d’espaces protégés plus ou moins permissifs en regard des activités qui peuvent y être menées, et avec des objectifs différents en termes de conservation (ex : Parcs Nationaux, Réserves Naturelles, Parcs Naturels Régionaux, Espaces Naturels Sensibles de départements et, plus spécifiquement en forêt, les Réserves Biologiques Dirigées et Intégrales). En France, environ 20 % du territoire est inclus dans des espaces protégés (CBD 2005). Dans la région Rhône-Alpes, plus de 12% des forêts sont incluses dans des réseaux de protection.
Les réseaux d’espaces protégés représentent un des principaux moyens de protection du patrimoine naturel à l’échelle locale et régionale. Cependant, il existe encore peu d’outils d’évaluation opérationnels permettant de rendre compte de l’efficacité réelle de ces réseaux pour la protection du patrimoine naturel et pour y établir des suivis de la biodiversité dans le temps (Hanski & Walsh 2004; Devictor et al. 2007). Les listes rouges d’espèces menacées, par exemple, sont souvent limitées aux vertébrés et aux végétaux supérieurs. De plus, au-delà des espèces remarquables ou patrimoniales, les espèces « ordinaires » et la diversité globale des espèces et de leurs fonctions sont tout aussi importantes pour le maintien du fonctionnement des écosystèmes. Il existe donc une forte lacune d’outils spatialisés et opérationnels, complémentaires à ces inventaires d’espèces, qui contribueraient efficacement au diagnostic de l’état de la biodiversité dans les espaces protégés.
Dans ce domaine, les indicateurs indirects de biodiversité peuvent jouer un rôle important. En forêt, leur utilisation est fondée sur l’hypothèse que l’analyse de l’hétérogénéité de la structure, de la composition et de l’organisation spatiale des peuplements3 dans un paysage forestier (ex : abondance et diversité des bois morts ou des gros arbres, diversité et continuité des peuplements dans l’espace) permet de renseigner sur la diversité des espèces présentes dans ce paysage (Noss 1999). Ces indicateurs peuvent être classés en deux catégories : 1° Ceux qui sont basés sur l’analyse de l’hétérogénéité locale de la structure des peuplements. Ils partent du principe que plus la
3
Un peuplement forestier est une surface forestière qui se distingue de la forêt environnante par sa structure (répartition en classes d’âge ou de diamètres, stratification de la canopée, proportion d’arbres vivants et morts) et par sa composition en essences. Un paysage forestier est souvent constitué d’une mosaïque de peuplements différents.
CemOA
: archive
ouverte
d'Irstea
5 structure d’un peuplement est hétérogène, plus celui-ci offre une grande variété de ressources permettant d’accueillir un nombre élevé d’espèces animales et végétales. 2° les indicateurs basés sur l’analyse de l’hétérogénéité spatiale du paysage forestier, qui joue également un rôle essentiel dans le maintien de la biodiversité. En effet, un paysage constitué d’une diversité d’habitats forestiers et de peuplements aux structures différentes permet à davantage d’espèces de cohabiter qu’un paysage homogène (Rosenzweig 1995; Tamme et al. 2010). Dans cette étude, nous proposons de combiner ces deux types d’indicateurs indirects de biodiversité pour développer une méthode innovante de diagnostic de la biodiversité forestière potentielle. Cette méthode sera appliquée à titre d’exemple dans deux Espaces Naturels Sensibles (ENS) de l’Isère.
1.3
Objectif
L’objectif de ce projet est de réaliser un diagnostic comparatif de la biodiversité forestière potentielle sur un gradient de gestion dans deux ENS du Vercors isérois. Le diagnostic s’appuie sur la construction d’une grille de notation basée sur le principe qu’il existe une relation positive entre l’hétérogénéité d’un paysage forestier (composition, agencement dans l’espace, hétérogénéité de la structure des peuplements) et le nombre d’espèces qui peuvent y être présentes. Cette grille comprend trois niveaux d’analyse emboîtés : (i) catégories de milieux, (ii) critères et (iii) indicateurs, ces derniers reflétant différents aspects de l’hétérogénéité du paysage forestier en lien avec la biodiversité. Les résultats permettent de quantifier la biodiversité potentiellement présente dans les différents sites étudiés et de mettre en évidence leurs « points forts » et leurs « points faibles » respectifs pour la conservation de la biodiversité forestière.
2 S
ITES D’
ÉTUDELa zone d’étude correspond à la partie iséroise du massif du Vercors, qui comprend différents types d’espaces protégés dont 4 sites Natura 2000, 7 Espaces Naturels Sensibles de département (ENS) et 2 Réserves Biologiques Intégrales (RBIs) (Figures 1a et b).
CemOA
: archive
ouverte
d'Irstea
7 potentiel de biodiversité dans les deux statuts de protection avec un diagnostic à l’échelle du site dans sa globalité. Pour le site d’Engins, il semble qu’aucune étude similaire n’ait été réalisée à ce jour.
3 C
ONSTRUCTION DE LA GRILLE DE NOTATION3.1
Principe du diagnostic de la biodiversité potentielle
Dans cette étude, la biodiversité potentielle désigne la diversité maximale en espèces qui peut être présente dans un écosystème à différentes échelles en fonction de ses caractéristiques actuelles, sans préjuger de sa biodiversité réelle.
La biodiversité potentielle est évaluée sur la base d’une grille de notation multicritères comprenant trois niveaux d’analyse :
1/ Quatre catégories de milieux qui correspondent au découpage du paysage forestier en différents secteurs : les milieux ouverts (non forestiers), l’interface forêt/non-forêt (lisières), les milieux forestiers et les forêts publiques. Les forêts publiques ont été considérées à part car davantage d’informations y étaient disponibles, notamment des cartographies des structures de peuplements,
2/ Plusieurs critères au sein de chaque catégorie, qui définissent la nature des éléments mesurés dans le paysage forestier pour l’évaluation du potentiel de biodiversité,
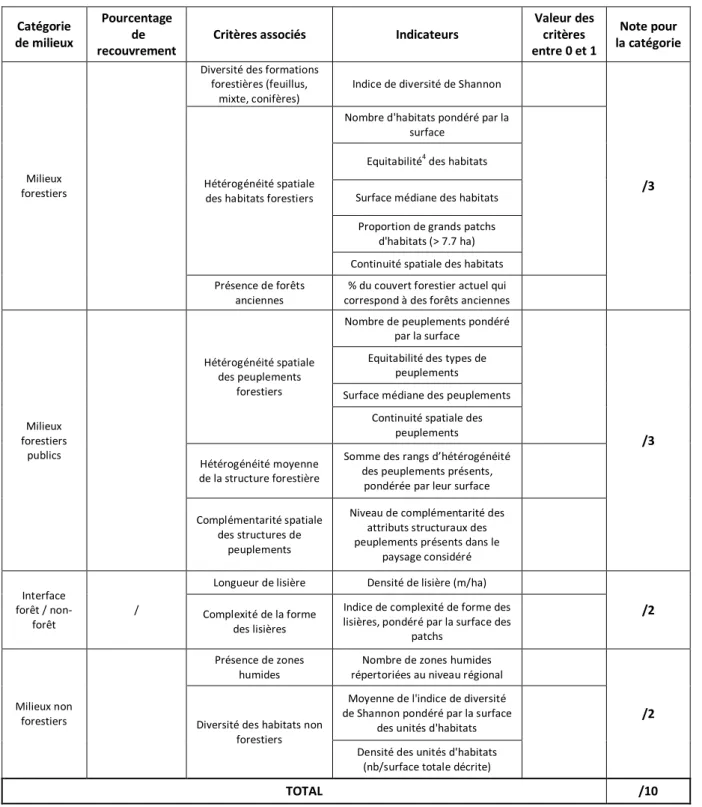
3/ Un ou plusieurs indicateurs par critère, qui correspondent à des mesures quantitatives permettant d’évaluer les critères (Tableau 1). Tous les indicateurs reflètent des caractéristiques du paysage forestier qui sont considérées comme ayant un effet positif et de même importance (pas de pondération) sur la biodiversité. Une valeur élevée pour un indicateur peut donc compenser une valeur faible d’un autre indicateur.
Le principe de fonctionnement de la grille de notation est le suivant : chaque indicateur est d’abord calculé à partir de différentes sources de données puis sa valeur est « transposée » entre 0 et 1. La transposition permet de rendre comparable des indicateurs avec des unités et/ou des échelles de valeurs très différentes. Elle permet également de hiérarchiser les sites étudiés entre eux, en prenant comme références les deux sites ayant respectivement la plus forte et la plus faible valeur pour l’indicateur considéré. La transposition se fait de la manière suivante :
CemOA
: archive
ouverte
d'Irstea
8
Où I = la valeur prise par l’indicateur considéré pour un site donné ; Imin = la valeur minimale atteinte par l’indicateur parmi l’ensemble des sites et I max = la valeur maximale atteinte par l’indicateur parmi l’ensemble des sites.
Ainsi, le site ayant la valeur la plus faible pour un indicateur donné obtient la valeur 0 pour cet indicateur, celui qui a la valeur la plus élevée, la valeur 1 et les deux autres sites ont des valeurs intermédiaires qui dépendent de leur proximité aux sites de référence pour l’indicateur considéré.
Lorsqu’un critère comprend plusieurs indicateurs, ceux-ci sont d’abord additionnés puis le résultat est transposé entre 0 et 1 afin d’obtenir un résultat comparable entre critères. Ensuite, les valeurs obtenues pour les critères sont additionnées au sein de chaque catégorie, ce qui donne une note globale par catégorie, la note maximale correspondant au nombre de critères dans la catégorie. La note globale pour le site correspond ensuite au total des notes obtenues pour les quatre catégories (Tableau 1). CemOA : archive ouverte d'Irstea / Cemagref
9
Tableau 1. Grille de notation utilisée pour le diagnostic de la biodiversité forestière potentielle dans les sites d’Engins et des Ecouges. Catégorie de milieux Pourcentage de recouvrement
Critères associés Indicateurs
Valeur des critères entre 0 et 1 Note pour la catégorie Milieux forestiers
Diversité des formations forestières (feuillus,
mixte, conifères)
Indice de diversité de Shannon
/3
Hétérogénéité spatiale des habitats forestiers
Nombre d'habitats pondéré par la surface
Equitabilité4 des habitats
Surface médiane des habitats Proportion de grands patchs
d'habitats (> 7.7 ha) Continuité spatiale des habitats Présence de forêts
anciennes
% du couvert forestier actuel qui
correspond à des forêts anciennes
Milieux forestiers publics Hétérogénéité spatiale des peuplements forestiers
Nombre de peuplements pondéré par la surface
/3
Equitabilité des types de peuplements
Surface médiane des peuplements Continuité spatiale des
peuplements Hétérogénéité moyenne
de la structure forestière
Somme des rangs d’hétérogénéité des peuplements présents,
pondérée par leur surface
Complémentarité spatiale des structures de
peuplements
Niveau de complémentarité des attributs structuraux des peuplements présents dans le
paysage considéré Interface forêt / non-forêt /
Longueur de lisière Densité de lisière (m/ha)
/2
Complexité de la forme des lisières
Indice de complexité de forme des lisières, pondéré par la surface des
patchs Milieux non forestiers Présence de zones humides
Nombre de zones humides
répertoriées au niveau régional
/2
Diversité des habitats non forestiers
Moyenne de l'indice de diversité de Shannon pondéré par la surface
des unités d'habitats Densité des unités d'habitats
(nb/surface totale décrite)
TOTAL /10
4
L’équitabilité mesure le rapport entre une distribution « équitable » de la surface des peuplements ou habitats où chacun d’eux occuperait une surface identique, et une distribution observée des surfaces dans la zone considérée.
CemOA
: archive
ouverte
d'Irstea
11 de leur structure. La structure d’un peuplement forestier peut être définie comme la distribution des arbres (vivants et morts) en classes de diamètres et d’âges, en strates de canopée et par leur agencement dans l’espace. Lorsqu’un peuplement est caractérisé par une forte diversification de ses caractéristiques structurales (ex : diversité en classes d’âges ou en essences) et par l’abondance d’éléments particuliers de la structure (ex : gros arbres vivants et morts, bois mort), on dit qu’il présente une structure hétérogène.
Une mosaïque forestière comprenant des peuplements à la structure hétérogène et une forte hétérogénéité spatiale des patchs, fournit une grande diversité de ressources et d’habitats à différentes échelles spatiales, permettant le maintien d’une importante biodiversité forestière.
Dans cette étude, plusieurs mosaïques de patchs différentes seront analysées (ex : mosaïque de peuplements et mosaïque d’habitats forestiers). L’analyse de chacune d’entre elles apporte des informations complémentaires pour le diagnostic de la biodiversité potentielle dans un espace protégé donné.
3.3
Supports cartographiques
La grande majorité des données utilisées dans ce travail ont été rassemblées ou produites dans le cadre de la thèse de M. Redon (2012). L’analyse de l’hétérogénéité de la mosaïque forestière s’appuie sur plusieurs supports cartographiques, qui ont été préparés avec le logiciel ArcGIS 9.3.
Dans le cas des milieux forestiers, deux supports cartographiques ont été utilisés. Le premier est un raster de 10 m de résolution représentant les habitats forestiers selon la typologie européenne Corine Biotopes niveau 2 (10 classes) : hêtraies, hêtraies-sapinières, pessières, sapinières, forêts de pente et de ravins, forêts thermophiles de chêne, pinèdes à crochets, pinèdes sylvestres, forêts alluviales et zones ouvertes (Figure 4b). Ce raster a été produit par classification d’images satellites SPOT, dans le cadre du projet de CartoVege financé par le MEDDTL (voir Breton (2011) pour les détails de la méthodologie). Le second support correspond à la carte des forêts anciennes réalisée par l’INRA de Nancy (voir Dupouey (2007) pour la méthodologie).
Dans la catégorie « milieux forestiers publics », l’évaluation de l’ensemble des critères est basée sur l’analyse d’un raster de 10 m de résolution représentant 7 types de peuplements forestiers : futaies irrégulières (≥ 3 strates), futaies à deux strates, futaies à une strate, mélange taillis – futaie, taillis simple, jeunes plantations et jeunes futaies (Figure 3 et Figure 4a). Ce raster est issu de l’assemblage, de la simplification puis de l’harmonisation de cartes vectorisées des structures de peuplements présentes dans les documents d’aménagement de l’ONF (donc disponibles uniquement
CemOA
: archive
ouverte
d'Irstea
12 pour les forêts publiques, nous n’avons pas trouvé d’équivalent pour les forêts privées dans la zone d’étude). La carte résultante a été complétée et/ou actualisée par photo-interprétation et validée par des visites de terrain et par consultation de plusieurs agents de l’ONF. Elle a ensuite été convertie en raster.
L’analyse de l’interface forêt/non-forêt est basée sur la cartographie du couvert forestier produite par le Joint Research Centre (JRC) au niveau européen avec une résolution de 25 m (Pekkarinen & Reithmaier 2009). Cette carte sous format raster est en libre accès. Elle a été ré-échantillonnée à une résolution de 10 m pour être homogène avec les autres rasters utilisés.
Dans le cas des milieux non forestiers, deux supports différents ont été utilisés. Le premier est une cartographie des habitats naturels non forestiers, issue de la carte des habitats réalisée par le Conservatoire Botanique National Alpin sur le massif du Vercors. Cette carte est sous format vecteur (polygones) (Figure 4c). Le deuxième support correspond à la cartographie des zones humides réalisée pour la région Rhône-Alpes par le conservatoire des espaces naturels AVENIR, qui prend en compte toutes les zones humides d’une surface supérieure à 1 ha (cf. Figure 1b).
Afin d’obtenir un jeu complet de données cartographiques pour chaque ENS et pour chaque RBI, les différents supports ont été rassemblés dans un même environnement SIG puis découpés avec des outils adaptés disponibles dans la boîte à outils du logiciel ArcGIS 9.3.
CemOA
: archive
ouverte
d'Irstea
14
3.4
Justification et calcul des critères et indicateurs utilisés dans la grille de notation
3.4.1 Choix des critères et indicateurs
Les critères et indicateurs utilisés dans l’analyse ont été sélectionnés sur la base d’une importante étude bibliographique. Ils reflètent tous des caractéristiques des peuplements ou de la mosaïque forestière dont les relations avec la biodiversité ont été démontrées. Ils permettent de quantifier l’hétérogénéité de la mosaïque forestière résultant notamment des gradients environnementaux (ex : hétérogénéité spatiale des habitats forestiers) et de la gestion forestière ancienne et actuelle (ex : hétérogénéité moyenne des structures de peuplements). Ces critères et indicateurs apportent des informations complémentaires : chacun d’entre eux reflète des caractéristiques favorables à des groupes d’espèces différents et leurs valeurs sont faiblement corrélées.
3.4.2 Milieux forestiers
Diversité des formations forestières
Dans une mosaïque forestière, la présence de différentes formations forestières (i.e. forêts feuillues, de conifères ou mélangées feuillus/conifères) crée une hétérogénéité du milieu forestier dans l’espace (ex : variation de la composition des sols ou de l’éclairement du sous-bois au cours de l’année). Cela favorise la diversification des espèces forestières dans la mosaïque paysagère, en particulier pour la végétation de sous-bois et les oiseaux (Kati et al. 2009; Chávez & Macdonald 2010). La diversité des formations forestières est calculée à l’aide d’un indice de diversité de Shannon :
Où ni est le nombre de patchs du type de forêt i et N est le nombre total de formations forestières dans la mosaïque considérée (= 3 dans cette étude).
Pour calculer cet indice, la carte des habitats a été reclassifiée en quatre classes : feuillus, conifères, mélange et non-forêt. L’indice varie entre 0 lorsqu’un seul type de forêt est présent et ln(Nmax) (i.e. ln(3) = 1.1 dans la zone d’étude) lorsque les trois types de formations forestières sont présents et occupent des surfaces équivalentes (la classe non-forêt est exclue des calculs).
CemOA
: archive
ouverte
d'Irstea
15 Hétérogénéité spatiale des habitats forestiers
La présence d’une diversité d’habitats dans une mosaïque paysagère donnée permet le maintien simultané d’espèces spécialistes de chacun des habitats présents et d’espèces qui dépendent de plusieurs habitats (Fahrig et al. 2011). Par exemple, de nombreuses espèces (ex : certains insectes, amphibiens et vertébrés) occupent des habitats différents au cours de leur cycle de vie ou au cours de l’année et sont dépendants de leur proximité dans l’espace (Noss 1987; Fahrig 2003). De nombreuses études ont montré des relations positives entre l’hétérogénéité des habitats dans la mosaïque paysagère et la diversité de plusieurs groupes d’espèces (ex : la flore (Dufour et al. 2006), les mammifères (Fraser 1998), les oiseaux (Ewers et al. 2005) et les invertébrés (Werner & Raffa 2000)).
Ce critère est évalué avec cinq indicateurs. Quatre d’entre eux sont des indices quantitatifs souvent utilisés en écologie du paysage : 1° nombre d’habitats différents répertoriés dans la zone considérée, pondéré par sa surface ; 2° équitabilité des habitats (rappelons que l’équitabilité mesure le rapport entre une distribution « équitable » de la surface des types de patchs où chaque type occuperait une surface identique et la distribution observée des surfaces dans la zone considérée) ; 3° surface médiane des habitats et 4° continuité spatiale des habitats. Ces indices ont été calculés à l’aide du logiciel libre Fragstats 3.3 (McGarical & Marks 1994), les formules mathématiques sont détaillées dans le tableau 2.
Le cinquième indicateur correspond à la proportion de grands patchs d’habitats présents dans la mosaïque forestière considérée. La taille des patchs est en effet un facteur clé à prendre en compte puisque dans une mosaïque paysagère donnée, si les patchs d’habitats sont trop petits par rapport aux besoins des espèces qui en dépendent, celles-ci peuvent être amenées à disparaître (Hanski & Walsh 2004; Warren et al. 2005). Cependant, c’est un facteur difficile à prendre en compte car chaque espèce perçoit la mosaïque paysagère à une échelle et d’une manière qui lui est propre (Wiens 1989). Quelques études se sont intéressées à la détermination de la taille minimale des patchs nécessaires à certaines espèces, mais elles sont encore rares (Freemark & Merriam 1986; SCF 2004). Ce facteur a été de ce fait très peu utilisé pour réaliser des diagnostics de biodiversité. L’indicateur de taille des patchs développé ici est basé sur la largeur de la lisière et sur la taille de la zone cœur. CemOA : archive ouverte d'Irstea / Cemagref
17 Présence de forêts anciennes
L’ancienneté de l’état boisé reflète la continuité du couvert forestier qui assure le maintien de conditions du milieu forestier stables au cours du temps et constitue un facteur essentiel pour de nombreuses espèces strictement forestières (ex : flore de sous-bois (Honnay et al. 1998; Susanne & Lawesson 2000), lichens (McMullin et al. 2010) et insectes saproxyliques (Siitonen 2001; Sverdrup-Thygeson 2002)). L’indicateur associé correspond à la proportion des forêts actuelles qui étaient déjà mentionnées sur la carte d’Etat-Major de 1850.
3.4.3 Milieux forestiers publics
Hétérogénéité spatiale des peuplements forestiers
Dans une mosaïque intra-forestière, la présence de différents types de peuplements forestiers contribue à la diversification de la composition en essences, de la structure et des stades de développement (ex : stades jeunes, matures et vieux) de la forêt et fournit ainsi une grande diversité de ressources permettant à de nombreuses espèces de cohabiter (Beese & Bryant 1999; Haveri & Carey 2000; Werner & Raffa 2000).
Ce critère est évalué à l’aide de quatre indicateurs quantitatifs : 1° nombre de types de peuplements répertoriés dans la zone considérée, pondéré par sa surface ; 2° équitabilité des types de peuplements; 3° surface médiane des peuplements et 4° continuité spatiale des différents types de peuplements. Ces indicateurs sont calculés de la même manière que dans le cas du critère d’hétérogénéité spatiale des habitats forestiers (cf. tableau 2) ; ils ont également été calculés à l’aide du logiciel libre Fragstats 3.3 (McGarical & Marks 1994).
Hétérogénéité moyenne de la structure forestière
De nombreuses études ont montré des relations positives entre l’hétérogénéité de la structure des peuplements forestiers et l’abondance ou la richesse de différents groupes d’espèces forestières (ex : espèces saproxyliques5 (Martikainen et al. 2000; Bütler et al. 2004), oiseaux (Willson 1974; Kati
et al. 2009), espèces cavernicoles (Ranius 2002; Humphrey et al. 2004) et végétation de sous-bois
(Bagnaresi et al. 2002; Ares et al. 2009)).
Ce critère est basé sur l’analyse des différents niveaux d’hétérogénéité structurale présents dans une mosaïque forestière donnée en fonction des types de peuplements qui y sont présents. Une étude préalable (voir thèse M. Redon 2012) a permis de hiérarchiser les types de peuplements
5
Liées pendant tout ou une partie de leur cycle de vie à du bois mort ou mourant (Good & Speight 1996).
CemOA
: archive
ouverte
d'Irstea
18 cartographiés dans la zone d’étude en fonction de l’hétérogénéité de leur structure (cf. Figure 2 et Tableau 3 pour le classement des peuplements). L’hétérogénéité moyenne des structures de peuplements correspond à la somme pondérée des rangs d’hétérogénéité des peuplements présents dans la mosaïque considérée :
Où ci est le rang d’hétérogénéité structurale du type de peuplement i (i.e. 7 pour les futaies irrégulières et 1 pour les jeunes futaies) ; Sij est la surface du patch j du type de peuplement i et TA est la surface totale de la mosaïque considérée.
Ce critère est compris entre 0 dans le cas d’une zone non forestière et 7 lorsque la mosaïque est composée uniquement de futaies irrégulières.
Tableau 3. Classement des peuplements forestiers pour quatre indices d’hétérogénéité structurale et en fonction de leur hétérogénéité globale. Les lettres représentent des différences significatives dans les valeurs moyennes des indices entre peuplements ; « a » correspond au peuplement ayant la valeur la plus élevée, « b » à une valeur intermédiaire et « c » à la valeur la plus faible, selon les résultats de tests de comparaison de Holm-Bonferroni.
Peuplement Hétérogénéité des diamètres Volume de bois mort Richesse en essences Densité des gros bois vivants RANGS d’hétérogénéité structurale Jeune futaie b c b c 1 Jeune plantation bc ab bc bc 2 Taillis simple c bc a c 3 Futaie monostrate c a c a 4 Mélange taillis-futaie ab b a bc 5
Futaie à deux strates ab ab b a 6
Futaie irrégulière a a b ab 7
Complémentarité spatiale des structures de peuplements
Ce critère permet de prendre en compte l’environnement des patchs : par exemple, un peuplement irrégulier entouré de plantations ne sera sans doute pas aussi favorable au maintien de la biodiversité forestière que le même peuplement irrégulier entouré d’un mélange de taillis et de futaies. La complémentarité spatiale des structures de peuplement reflète le nombre de caractéristiques structurales (i.e. volume de bois mort (m3/ha), hétérogénéité des diamètres (coefficient de variation), densité des gros bois vivants (Ø ≥ 45 cm ; m²/ha) ou richesse en essences) représentées avec un rang « a » dans la mosaïque forestière considérée, en fonction de sa composition en types de peuplements (cf. tableau 3). Le statut de jeune peuplement compte également comme une catégorie complémentaire. Ce critère est catégoriel et varie entre 1 et 4. Il est égal à 1 lorsque la mosaïque est dominée par un seul type de peuplement. Il est maximal quand la mosaïque comprend au moins quatre types de peuplements différents, qui permettent d’obtenir des
CemOA
: archive
ouverte
d'Irstea
20
Où Pi correspond au périmètre du patch i et min Pi est égal au périmètre pris par une surface équivalente compactée au maximum (même nombre de pixels mais forme la plus simple possible : carré ou cercle).
3.4.5 Milieux non forestiers
Présence de zones humides
Les zones humides sont des milieux inondés une partie ou toute l’année qui sont caractérisés par une végétation spécifique, au moins en partie hygrophile. Elles peuvent être présentes sous le couvert forestier (ruisseaux, mares temporaires…) ou en bordure (tourbières, marais…). Ces zones humides représentent de petites surfaces sur les sites étudiés mais elles contribuent largement à leur biodiversité par l’apport de nombreuses espèces animales et végétales qui y sont inféodées.
L’indicateur associé correspond au nombre de zones humides répertoriées au niveau régional qui sont présentes dans les différents sites.
Diversité des habitats non forestiers
La diversité des habitats non forestiers contribue également à la richesse de la biodiversité dans la mosaïque forestière. De nombreuses espèces forestières, par exemple, utilisent les zones ouvertes (clairières, prairies…) pour chasser ou pour se nourrir (ex : chouettes, insectes floricoles…). La diversité des habitats ouverts contribue également à diversifier la composition des lisières, produisant une plus grande variété de ressources pour les espèces qui y sont inféodées.
Ce critère est calculé à partir de la cartographie des habitats naturels du CBNA, extraite sur les zones non forestières. Cette carte est constituée de polygones contenant chacun un ou plusieurs habitats naturels en proportions variables. Dans la plupart des cas, il n’est donc pas possible d’associer un polygone donné à un habitat donné. Par contre, pour chaque polygone, le nom des différents habitats présents et leurs proportions respectives sont connues, permettant d’évaluer la diversité des habitats qui y sont présents.
La diversité des habitats non-forestiers a été calculée en deux étapes : 1/ évaluation de la diversité des habitats présente dans chaque polygone avec un indice de diversité de Shannon (pour les polygones contenant un seul habitat l’indice est égal à 0, tandis que sa valeur est élevée pour les polygones contenant un grand nombre d’habitats différents et dont les proportions sont équivalentes) ; 2/ calcul du critère de la manière suivante :
CemOA
: archive
ouverte
d'Irstea
21
où SHi est l’indice de Shannon représentant la diversité des habitats dans le polygone i, si est la surface du polygone i (ha), N est le nombre total de polygones dans la mosaïque considérée et TA est la surface totale de la mosaïque (ha).
Ce critère est égal à lorsque tous les polygones contiennent un seul habitat (indice de Shannon nul pour tous les polygones). Sa valeur est élevée lorsque chaque polygone contient une diversité d’habitats et qu’il y a une forte densité de polygones dans le site considéré. Ce critère prend en compte la surface des polygones ainsi que celle de la mosaïque paysagère dans son ensemble, ce qui permet d’obtenir des valeurs comparables entre sites avec des surfaces très différentes.
CemOA
: archive
ouverte
d'Irstea
22
4 R
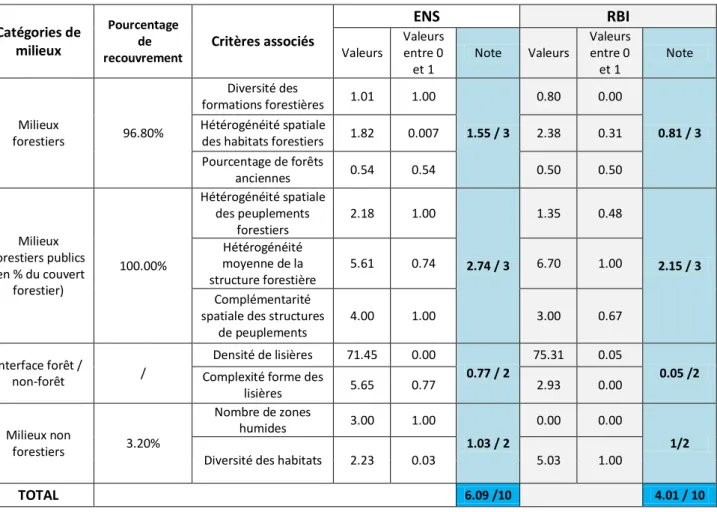
ÉSULTATSPour chacun des quatre sites, on obtient une grille de notation complète avec une valeur pour chaque critère et pour chaque catégorie de milieux résultant en une note globale pour le site (Tableaux 4 et 5). Les notes globales sont moyennes dans l’ensemble. L’Espace Naturel Sensible (ENS) des Ecouges a le plus fort potentiel de biodiversité selon les critères considérés, avec une note de 6.09 /10.
Tableau 4. Grilles de notation obtenues pour l’ENS et la RBI des Ecouges.
Catégories de milieux Pourcentage de recouvrement Critères associés ENS RBI Valeurs Valeurs entre 0 et 1 Note Valeurs Valeurs entre 0 et 1 Note Milieux forestiers 96.80% Diversité des formations forestières 1.01 1.00 1.55 / 3 0.80 0.00 0.81 / 3 Hétérogénéité spatiale
des habitats forestiers 1.82 0.007 2.38 0.31 Pourcentage de forêts anciennes 0.54 0.54 0.50 0.50 Milieux forestiers publics (en % du couvert forestier) 100.00% Hétérogénéité spatiale des peuplements forestiers 2.18 1.00 2.74 / 3 1.35 0.48 2.15 / 3 Hétérogénéité moyenne de la structure forestière 5.61 0.74 6.70 1.00 Complémentarité
spatiale des structures de peuplements 4.00 1.00 3.00 0.67 Interface forêt / non-forêt / Densité de lisières 71.45 0.00 0.77 / 2 75.31 0.05 0.05 /2 Complexité forme des
lisières 5.65 0.77 2.93 0.00 Milieux non forestiers 3.20% Nombre de zones humides 3.00 1.00 1.03 / 2 0.00 0.00 1/2 Diversité des habitats 2.23 0.03 5.03 1.00
TOTAL 6.09 /10 4.01 / 10 CemOA : archive ouverte d'Irstea / Cemagref
23
Tableau 5. Grilles de notation obtenues pour l’ENS et la RBI d’Engins.
Catégories de milieux Pourcentage de recouvrement Critères associés ENS RBI Valeurs Valeurs entre 0 et 1 Note Valeurs Valeurs entre 0 et 1 Note Milieux forestiers 47.00% Diversité des formations forestières 0.90 0.48 1.34 / 3 0.86 0.28 2.28 / 3 Hétérogénéité spatiale
des habitats forestiers 1.81 0.00 3.65 1.00 Pourcentage de forêts anciennes 0.86 0.86 0.99 0.99 Milieux forestiers publics (en % du couvert forestier) 86.50% Hétérogénéité spatiale des peuplements forestiers 1.40 0.52 1.48 / 3 0.56 0.00 0.67 / 3 Hétérogénéité moyenne de la structure forestière 3.75 0.29 2.52 0.00 Complémentarité
spatiale des structures de peuplements 3.00 0.67 3.00 0.67 Interface forêt / non-forêt / Densité de lisières 113.26 0.53 1.53 / 2 150.56 1.00 1.69 / 2 Complexité forme des
lisières 6.44 1.00 5.37 0.69 Milieux non forestiers 53.00% Nombre de zones humides 1.00 0.33 0.33/ 2 0.00 0.00 0.45 / 2 Diversité des habitats 2.14 0.00 3.43 0.45
TOTAL 4.68 /10 5.09 / 10
4.1
Comparaison des Espaces Naturels Sensibles et des Réserves Biologiques
Intégrales
La comparaison des ENS et des RBIs entre elles permet de voir s’il existe des caractéristiques propres aux sites gérés d’une part et non-gérés d’autre part. Nous comparons ensuite l’ENS et la RBI au sein de chaque site (Ecouges, Engins) afin de déterminer s’il existe un effet de la gestion sur les caractéristiques de la mosaïque forestière en lien avec la biodiversité. Ces comparaisons permettent également de faire apparaître des « points forts » et des « points faibles » des différents sites pour la conservation de la biodiversité.
4.1.1 Comparaison des RBIs entre elles
Le seul point commun aux deux RBIs est une forte complémentarité spatiale des structures de peuplements (Figure 7). La RBI des Ecouges se démarque nettement de celle d’Engins par des valeurs
nettement plus élevées pour trois critères : la diversité des habitats non forestiers (très élevée par rapport à la surface considérée, qui est beaucoup plus petite que dans le site d’Engins : 3.2 % contre 42.8 %), l’hétérogénéité spatiale des peuplements et l’hétérogénéité moyenne de la structure
CemOA
: archive
ouverte
d'Irstea
24 forestière. Pour tous les autres critères, les valeurs sont plus élevées dans la RBI d’Engins, qui a d’ailleurs une note globale plus élevée.
4.1.2 Comparaison des ENS entre eux
Les ENS ont en commun une très faible diversité des habitats non forestiers quelle que soit la surface considérée (53 % de la surface à Engins, 12.2 % aux Ecouges), une forte complexité de la forme des lisières et un pourcentage de forêts anciennes moyen à élevé (Figure 7 ; Tableaux 4 et 5).
Pour l’ensemble des autres critères sauf la longueur de lisières, les valeurs sont plus élevées dans l’ENS des Ecouges, qui a d’ailleurs une note globale plus élevée que celle d’Engins.
Ces résultats montrent que dans le secteur des Ecouges, l’ENS (géré) présente une biodiversité
potentielle prédite plus forte que la RBI (non gérée), tandis que dans le secteur d’Engins, c’est l’inverse.
4.1.3 Comparaison entre les modalités gérées (ENS) et non-gérées (RBIs)
La différence entre les modalités gérées et non-gérées est plus marquée dans les Ecouges (1.41 points d’écart sur 10) que dans le secteur d’Engins (1.08 points d’écart sur 10).
La comparaison des notes obtenues pour les critères dans les deux sites montre que sur 10
critères, 7 varient dans le même sens entre les modalités gérées et non gérées, même si les
différences sont parfois faibles. Quatre critères sont systématiquement plus élevés dans les ENS : la diversité des formations forestières, l’hétérogénéité spatiale des peuplements forestiers, la complexité de la forme des lisières et le nombre de zones humides. Trois critères sont au contraire
toujours plus élevés dans les RBIs : l’hétérogénéité spatiale des habitats forestiers, la longueur de
lisière et la diversité des habitats non forestiers (Tableaux 4 et 5).
Pour les trois critères restants, les situations sont variables. Dans les deux sites, le pourcentage de forêts anciennes est similaire entre l’ENS et la RBI. Pour les critères d’hétérogénéité moyenne de la structure des peuplements et de complémentarité spatiale des structures, les tendances sont inversées d’un site à l’autre.
Ainsi, les différences entre les modalités gérées et non gérées touchent à des critères reflétant des caractéristiques de la mosaïque forestière très variées et ne semblent pas être liées à une catégorie de milieux en particulier (Figure 8).
CemOA
: archive
ouverte
d'Irstea
26 particulières (lapiazs, en limite du subalpin) favorables au développement de peuplements purs d’épicéas ou de pin à crochets qui y sont largement majoritaires, laissant peu de place aux peuplements feuillus.
L’hétérogénéité spatiale des peuplements forestiers est également plus élevée dans les zones gérées et résulte sans doute de la transition entre la gestion passée (traitement en taillis pour le charbonnage pendant des dizaines d’années qui a perduré jusqu’au milieu du XXème siècle) vers les pratiques sylvicoles actuelles qui favorisent le développement d’autres types de peuplements plus diversifiés (ex : futaies irrégulières, mélanges taillis-futaies…).
Le nombre de zones humides est également plus élevé dans les ENS. Cependant, il semble peu probable que la gestion forestière favorise la création de zones humides bien que dans certains cas, elle puisse contribuer à éviter leur colonisation par la forêt. Le résultat obtenu est certainement dû à la localisation géographique des ENS dans des secteurs où il existe un réseau hydrographique de surface (ce qui n’est pas le cas, par exemple, dans la RBI d’Engins) et au fait que leur surface est bien plus grande que celle des RBIs, augmentant la probabilité d’y trouver des zones humides. Ce résultat peut également être lié à la faible précision de la carte utilisée, qui ne prend en compte que les zones humides d’au moins 1 ha, ce qui est très élevé, surtout dans le contexte du massif du Vercors où le substrat est calcaire et retient peu l’eau.
Les résultats montrent également que les espaces gérés ont conservé une complexité de la forme des lisières importante tandis que la longueur des lisières y est globalement plus faible que dans les secteurs non gérés. Les ENS apportent donc quantitativement moins d’habitats aux espèces de lisières que les espaces non gérés. Par contre, les lisières présentes sont plus complexes, permettant une plus grande diversification des ressources pour les espèces que dans les espaces non gérés.
Effets de la non-gestion
Les espaces non gérés sont caractérisés par une forte diversité des habitats non forestiers. L’hétérogénéité spatiale des habitats forestiers y est également plus élevée que dans les zones gérées, notamment en raison d’une très grande variété d’habitats. Ces tendances peuvent s’expliquer par la grande variabilité des gradients environnementaux dans les RBIs. Dans celle d’Engins, par exemple, toutes les orientations sont présentes et le site est marqué par une alternance de dalles lapiazées et de petites falaises, favorable au maintien d’une grande diversité d’habitats forestiers et non forestiers. La RBI des Ecouges présente également une grande diversité de situations géomorphologiques (falaises, éboulis…) qui conduit à une importante diversité des sols.
CemOA
: archive
ouverte
d'Irstea
28 est cependant moins évidente pour d’autres critères pour lesquels on aurait pu s’attendre à un effet négatif de la gestion, par exemple le nombre de zones humides ou la complexité de la forme des lisières. Pour plusieurs critères, la part d’explication liée à la gestion et aux conditions environnementales semble difficile à établir. Par contre, dans le cas de l’hétérogénéité spatiale des peuplements forestiers, les valeurs plus élevées dans les zones gérées semblent clairement liées à une situation de transition entre différentes pratiques de gestion.
4.1.5 Conséquences en termes de biodiversité potentielle dans les modalités gérées et non gérées
Les résultats ne permettent pas de conclure clairement quant à un effet de la gestion ou de la non-gestion sur la biodiversité potentielle dans les espaces étudiés. Chacun des quatre sites est
caractérisé par une combinaison de valeurs de critères spécifique, reflétant différentes compositions et configurations de la mosaïque forestière favorisant le maintien de différentes composantes de la biodiversité forestière. Certaines caractéristiques sont plus marquées dans les
ENS et d’autres dans les RBIs mais les différences observées ne semblent pas toujours dues à un effet de la gestion pratiquée. De plus, les variations observées concernent des critères appartenant aux quatre catégories de milieux définis, et il est difficile de dégager des tendances générales.
4.2
Comparaison des potentialités dans les sites d’Engins et des Ecouges
Nous avons pu mettre en évidence des tendances communes aux deux sites en regard des différences entre les modalités gérées et non gérées. Cependant, les résultats obtenus varient d’un site à l’autre, et l’effet du site semble très structurant pour les caractéristiques de la mosaïque forestière en lien avec la biodiversité. Les deux sites sont en effet localisés dans des contextes environnementaux très différents et ont une histoire d’usage de la forêt différente, ce qui peut expliquer la variation des résultats obtenus.
Quel que soit le statut, géré ou non géré, le site d’Engins est caractérisé par des peuplements forestiers globalement moins hétérogènes que dans le site des Ecouges. Cette caractéristique est particulièrement marquée dans la RBI, où le site étant peu accessible pour l’exploitation de bois (il n’a pas été exploité significativement depuis plusieurs dizaines d’années), la faible hétérogénéité des peuplements est certainement liée aux conditions édaphiques (lapiazs calcaires souvent apparents et qui ne retiennent pas l’eau) et climatiques très difficiles, peu favorables au développement des arbres. Par contre, la majorité des forêts sont anciennes et le site comprend une mosaïque de milieux forestiers et non forestiers, se traduisant par une forte densité de lisières et complexité de la forme des lisières, bien que les valeurs soient variables entre l’ENS et la RBI. Dans l’ensemble, ce site
CemOA
: archive
ouverte
d'Irstea
29
semble donc particulièrement favorable aux espèces de lisières ou occupant indifféremment les milieux forestiers et non forestiers. Les forêts y sont sans doute moins riches en biodiversité que dans le site des Ecouges, mais elles semblent particulièrement favorables au maintien des espèces dépendantes de la continuité de l’état boisé.
Le secteur des Ecouges est caractérisé par une forte complémentarité spatiale des structures de peuplement et hétérogénéité moyenne des structures. L’hétérogénéité spatiale des peuplements forestiers y est également généralement plus élevée que dans le site d’Engins. Le site est presque entièrement dominé par la forêt, laissant peu de place aux zones de lisière (densité de lisières faible) et aux milieux non forestiers, qui sont cependant très diversifiés dans la RBI (Figure 7). Cette diversité des habitats sur de petites surfaces permet sans doute le maintien de nombreuses espèces généralistes capables d’utiliser différents habitats. Dans l’ensemble, le site des Ecouges se démarque par de fortes valeurs pour les critères en lien avec l’hétérogénéité/complémentarité de la structure des peuplements cohérentes avec la localisation du site dans des milieux très favorables à la croissance des arbres. Cette caractéristique peut également refléter des effets encore présents de la gestion récente, majoritairement en futaie irrégulière. Les forêts anciennes y sont par contre moins représentées que dans le site d’Engins, suggérant qu’une partie des forêts actuelles est peut-être issue de la recolonisation d’anciens pâturages. Ce secteur semble donc globalement très favorable
aux espèces utilisant préférentiellement l’intérieur des forêts, avec peut-être un déficit en espèces dépendantes de la continuité de l’état boisé dans les secteurs de forêts récentes. La biodiversité du
site est également alimentée dans la partie RBI par la présence de secteurs non boisés peu nombreux mais comprenant une grande diversité d’habitats.
CemOA
: archive
ouverte
d'Irstea
30
5 D
ISCUSSION GÉNÉRALELa méthode présentée dans cette étude permet de hiérarchiser les quatre sites en fonction de la biodiversité potentielle qui y est prédite. Le diagnostic de l’état de la biodiversité est abordé de façon indirecte et évalue un potentiel de biodiversité en fonction d’un maximum théorique dépendant des caractéristiques de la mosaïque forestière. Les résultats permettent de quantifier et de spatialiser les niveaux de biodiversité potentiellement présente dans la mosaïque forestière. Le principal avantage de ce type de méthode est qu’elle peut être adaptée à différentes situations. Par exemple, dans cette étude, nous avons inclus une catégorie de milieux « forêt publiques », dont les critères ne s’appliquent qu’à un sous-ensemble des forêts. Ainsi, en fonction des données disponibles, différentes configurations de la grille de notation peuvent être imaginées, permettant d’approfondir ou non l’analyse dans différentes catégories de milieux. Les critères présentés dans cette étude ont été développés à partir des données qui étaient disponibles dans la zone d’étude et devront certainement être adaptés pour une utilisation dans d’autres sites. Ce qui importe dans la mise en œuvre de ce type de méthode n’est pas la nature des critères utilisés (qui doivent cependant toujours être sélectionnés pour leurs liens avec une partie de la biodiversité) mais le principe général sur lequel elle est basée. Ce principe est qu’il existe des relations positives à différentes échelles entre l’hétérogénéité spatiale de la mosaïque forestière et la diversité des espèces cohabitant dans cette mosaïque, ce qui permet d’utiliser la mesure de l’hétérogénéité de la mosaïque paysagère comme une méthode de diagnostic indirect de l’état de la biodiversité à un instant donné. Ainsi, d’autres critères reflétant d’autres aspects de l’hétérogénéité de la mosaïque connus pour leurs liens avec la biodiversité peuvent être ajoutés, et d’autres peuvent être enlevés, sans changer la philosophie générale du diagnostic. Par contre, pour comparer des sites entre eux dans un territoire donné, il est évident qu’il vaut mieux utiliser des critères similaires.
Un autre intérêt majeur de la méthode est qu’elle facilite une comparaison rapide d’environnements très hétérogènes sur de grandes étendues grâce à l’utilisation d’un système de notation. L’intérêt d’une grille de notation hiérarchique est qu’il est possible d’affiner peu à peu l’analyse jusqu’au niveau pertinent en fonction des objectifs recherchés. A chaque niveau, les résultats peuvent être présentés sous la forme de graphes radars (cf. Figures 7 et 8) afin de dégager des tendances dans les résultats. De plus, le système de transposition des valeurs entre 0 et 1 utilise les maximas et minimas observés dans la zone d’étude comme références, ce qui permet de s’affranchir de la recherche d’un état de référence extérieur. Cependant, l’interprétation des notes obtenues doit se faire avec précautions, sans perdre de vue qu’elles ont du sens pour caractériser et classer les sites entre eux uniquement dans la zone d’étude considérée. De plus, les valeurs obtenues
CemOA
: archive
ouverte
d'Irstea
31 sont relatives et ne peuvent être interprétées qu’en comparant les sites les uns par rapport aux autres.
La diversité des critères utilisés permet de faire apparaître les « points forts » et les « points faibles » des différents sites les uns par rapport aux autres en termes de caractéristiques de la mosaïque forestière permettant le maintien de la biodiversité. Cette information peut avoir différentes applications comme par exemple : i) l’aide à la désignation de futurs espaces protégés qui pourront être choisis en fonction de caractéristiques de la mosaïque forestière complémentaires à celles des sites existants ; ii) la proposition de recommandations de gestion visant à développer l’hétérogénéité locale et/ou spatiale des peuplements dans les secteurs gérés où les critères correspondants ont des valeurs faibles ; iii) l’utilisation des résultats comme un état initial pour établir un suivi dans le temps de caractéristiques de la mosaïque forestière en lien avec la biodiversité. D’autre part, la variabilité des tendances observées entre critères montre l’importance de diversifier les critères pour réaliser une évaluation de l’état de la biodiversité dans un site donné ; chaque critère reflétant différentes composantes de l’hétérogénéité spatiale en lien avec différentes physionomies de la mosaïque forestière favorables au maintien de différents sous-ensembles de la biodiversité.
Il est également important de noter que cette méthode évalue surtout une biodiversité « ordinaire ». Les résultats ne peuvent donc pas toujours être corrélés avec la présence ou l’absence d’espèces ciblées sauf, éventuellement, si elles sont fortement liées à l’un des indicateurs ou des critères utilisés pour le diagnostic. Cette méthode peut être complétée par d’autres approches plus précises lorsqu’on s’intéresse à la présence d’espèces ou d’habitats présentant un intérêt particulier pour la conservation. CemOA : archive ouverte d'Irstea / Cemagref
32
6 B
IBLIOGRAPHIEAndré J (2005) Activité et diversité des organismes hétérotrophes : les clés du bouclage des cycles biogéochimiques et sylvigénétiques. In: Bois mort et à cavité - une clé pour les forêts vivantes. Lavoisier, Paris, pp 89 - 98
Ares A, Shanti B, Puettmann KJ (2009) Understory vegetation response to thinning disturbance of varying complexity in coniferous stands. Applied Vegetation Science 12:472-487
Bagnaresi U, Giannini R, Grassi G, Minotta G, Paffetti D, Pini Prato E, Proietti Placidi AM (2002) Stand structure and biodiversity in mixed, uneven-aged coniferous forests in the easthern Alps. Forestry 75 (4):357-364
Beese WJ, Bryant AA (1999) Effect of alternative silvicultural systems on vegetation and bird communities in coastal montane forests of British Columbia, Canada. Forest Ecology and Management 115:231-242
Breton V, Renaud J, Luque S (2011) Comment les outils de la télédétection peuvent aider à la cartographie des habitats forestiers ? Mise au point d'une méthode sur le massif du Vercors. Rendez-Vous Techniques de l'ONF 31:69-73
Butchart SHM, Walpole M, Collen B, van Strien A, Scharlemann JPW, Almond REA, Baillie JEM, Bomhard B, Brown C, Bruno J, Carpenter KE, Carr GM, Chanson J, Chenery AM, Csirke J, Davidson NC, Dentener F, Foster M, Galli AE, Galloway JN, Genovesi P, Gregory RD, Hockings M, Kapos V, Lamarque J-L, Leverington F, Loh J, McGeoch MA, McRae L, Minasyan A, Hernández Morcillo M, Oldfield TEE, Pauly D, Quader S, Revenga C, Sauer JR, Skolnik B, Spear D, Stanwell-Smith D, Stuart SN, Symes A, Tierney M, Tyrrell TD, Vié J-C, Watson R (2010) Global biodiversity: Indicators of recent declines. Science 328:1164-1168
Bütler R, Angelstam P, Ekelund P, Schlaepfer R (2004) Dead wood threshold values for the three-toed woodpecker presence in boreal and sub-Alpine forest. Biological Conservation 119 (3):305-318
CBD (2002) The United Nations Convention on Biological Diversity - 2010 Biodiversity target United Nations Environment Programme, doi: http://www.cbd.int/2010-target/
CBD (2005) Protected areas for achieving biodiversity targets. Secretariat of the Convention on Biological Diversity, Montreal, 26 p., doi: http://www.cbd.int/doc/publications/news-pa-supplement-en.pdf
Chávez V, Macdonald SE (2010) The influence of canopy patch mosaics on understory plant community composition in boreal mixed wood forest. Forest Ecology and Management 259:1067-1075
Dajoz R (2007) Les insectes et la forêt, rôle et diversité des insectes dans le milieu forestier. 2ème édition edn. Lavoisier, Paris. 668 p.
Devictor V, Godet L, Julliard R, Couvet D, Jiguet F (2007) Can common species benefit from protected areas? Biological Conservation 139 (1-2):29-36
Dufour A, Gadallah F, Wagner HH, Guisan A, Buttler A (2006) Plant species richness and environmental heterogeneity in a mountain landscape: effects of variability and spatial configuration. Ecography 29:573-584
Dupouey JL, Bachacou J, Cosserat R, Aberdam S, Vallauri D, Chappart G, Corvisier de Villèle M-A (2007) Vers la réalisation d'une carte géoréférencée des forêts anciennes de France. Revue Forestiere Francaise 191:85-98
EEA (2010) Assessing Biodiversity in Europe - the 2010 report. EEA, 58 p., doi:
http://www.eea.europa.eu/publications/assessing-biodiversity-in-europe-84
EU (2011) Forestry in the EU and the world. European Union, Luxembourg, doi:
http://epp.eurostat.ec.europa.eu/cache/ITY_OFFPUB/KS-31-11-137/EN/KS-31-11-137-EN.PDF CemOA : archive ouverte d'Irstea / Cemagref
33 Ewers RM, Didham RK, Wratten SD, Tylianakis JM (2005) Remotely sensed landscape heterogeneity as a rapid tool for assessing local biodiversity value in a highly modified New Zealand landscape. Biodiversity and Conservation 14:1469-1485
Fahrig L (2003) Effects of Habitat Fragmentation on Biodiversity. Annual Review of Ecological Evolution and Systematics 34:487-515
Fahrig L, Baudry J, Brotons L, Burel FG, Crist TO, Fuller RJ, Sirami C, Siriwardena GM, Martin J-L (2011) Functional landscape heterogeneity and animal biodiversity in agricultural landscapes. Ecology Letters 14:101-112
Fraser RH (1998) Vertebrate Species Richness at the Mesoscale: Relative Roles of Energy and Heterogeneity. Global Ecology and Biogeography Letters 7 (3):215-220
Freemark KE, Merriam HG (1986) Importance of Area and Habitat Heterogeneity to Bird Assemblages in Temperate Forest Fragments. Biological Conservation 36:115 - 141
Good JA, Speight MCD (1996) Saproxylic invertebrates and their conservation throughout Europe. Conseil de l'Europe, Strasbourg. 58 p.
Hanski I, Walsh M (2004) How much, How to? - Practical tools for forest conservation. BirdLife European Forest Task Force, Latvia. 48 p.
Haveri BA, Carey AB (2000) Forest management strategy, spatial heterogeneity, and winter birds in Washington. Wildlife Society Bulletin 28 (3):643-652
Helzer CJ, Jelinski DE (1999) The relative importance of patch area and perimeter-area ratio to grassland breeding birds. Ecological Applications 9 (4):1448-1458
Honnay O, Degroote B, Hermy M (1998) Ancient-forest plant species in Western Belgium: a species list and possible ecological mechanisms. Belgium Journal of Botany 130 (2):139-154
Humphrey JW, Sippola AL, Lemperiere G, Dodelin B, Alexander KNA, Butler JE (2004) Deadwood as an Indicator of biodiversity in european forests : from theory to operational guidance. In: Marchetti M (ed) Monitoring and Indicators of Forest Biodiversity in Europe - From Ideas to Operationality, vol 51. European Forest Institute, Joensuu, pp 193-206
Kati V, Dimopoulos P, Papaioannou H, Poirazidis K (2009) Ecological management of a Mediterranean mountainous reserve (Pindos National Park, Greece) using bird community as an indicator. Journal of Nature Conservation 17:47-59
Kati V, Poirazidis K, Dufrêne M, Halley J, Koradis G, Schindler S, Dimopoulos P (2010) Towards the use of ecological heterogeneity to design reserve networks: a case study from Dadia National Park, Greece. Biodiversity and Conservation 19 (6):1585-1597
Laurance SGW (2004) Responses of understory rain forest birds to road edges in central Amazonia. Ecological Applications 14 (5):1344-1357
Levrel H (2007) Quels indicateurs pour la gestion de la biodiversité? Paris. 94 p.
Martikainen P, Siitonen J, Punttila P, Kaila L, Rauh J (2000) Species richness of Coleoptera in mature managed and old-growth boreal forests in southern Finland. Biological Conservation 94 (2):199-209
McGarical K, Marks BJ (1994) FRAGSTATS - Spatial pattern analysis program for quantifying landscape structure. Oregon State University, Dolores, Colorado, 134 p., doi:
http://www.umass.edu/landeco/pubs/mcgarigal.marks.1995.pdf
McMullin RT, Duinker PN, Richardson DHS, Cameron RP, Hamilton DC, Newmaster SG (2010) Relationships between the structural complexity and lichen community in coniferous forests of southwestern Nova Scotia. Forest Ecology and Management 260:744 - 749
MCPFE (2007) State of Europe's forests 2007 - The MCPFE report on sustainable forest management in Europe. MCPFE, FAO, UNECE, Warsaw, Poland, 247 p., doi:
http://www.foresteurope.org/filestore/foresteurope/Publications/pdf/state_of_europes_for ests_2007.pdf
MEA (2005) Ecosystems and Human Well-being: Biodiversity synthesis World Resources Institute, 86 p., Washington, DC, doi: http://www.maweb.org/documents/document.354.aspx.pdf
Mladenoff DJ, White MA, Pastor J, Crow TR (1993) Comparing spatial pattern in unaltered old-growth and disturbed forest landscapes. Ecological Applications 3 (2):294-306
CemOA
: archive
ouverte
d'Irstea