Implication des Galectines-3, 7 et 9 dans la réponse immunitaire
contre le parasite Leishmania major
Mémoire présenté
à la Faculté des études supérieures et postdoctorales de l’Université Laval dans le cadre du programme de maîtrise en microbiologie-immunologie
pour l’obtention du grade de Maître ès sciences (M. Sc.)
DÉPARTEMENT DE MICROBIOLOGIE-INFECTIOLOGIE ET D'IMMUNOLOGIE FACULTÉ DE MÉDECINE
UNIVERSITÉ LAVAL QUÉBEC
2013
Résumé
La galectine-3, une protéine cytosolique de l’hôte est impliquée dans la réponse immunitaire contre Leishmania major, un parasite protozoaire causant une infection cutanée. Lors d’une infection avec la souche LV39 de L. major, la galectine-3 se libère dans le milieu extracellulaire et participe au recrutement de neutrophiles au site d’infection. La galectine-3 ne semble cependant pas impliquée lors d’infections avec la souche L. major Friedlin qui possède un arrangement de saccarides différent sur ses phosphoglycans. In
vitro, les deux souches de L. major sont en mesure de cliver la galectine-3, mais LV39
semble plus efficace. Différents essais ont été effectués in vivo afin de vérifier si les galectines-7 et 9 peuvent aussi avoir un rôle à jouer lors d’infection avec L. major. Alors qu’aucun effet significatif n’a pu être attribué à la galectine-9, l’absence de galectine-7 semble apporter une guérison plus rapide des lésions engendrées par L. major Friedlin.
Abstract
Our recent data suggest that galectin-3, a host cytosolic protein is involved in the immune response against the Leishmania major strain LV39, which induces cutaneous infection. Following infection with this parasite, galectin-3 is released in the extracellular space and plays a role in neutrophil recruitment. In contrast, in another L. major strain called Friedlin, which expresses a different glycan structure on the phosphoglycans, galectin-3 has no impact on the host immune response and disease outcome. Our laboratory previously reported that L. major LV39 cleaves galectin-3. Our data suggest that both L. major strains are able to cleave galectin-3 in vivo, although the cleavage by LV39 is more efficient than that by Friedlin. We also addressed the role of galectins-7 and 9 in a murine L. major infection model. Lack of galectin-9 did not alter the pathogenesis of Leishmania infection. In contrast, lack of galectin-7 contributed to improve healing process.
Table des matières
Résumé ... ii
Abstract ... iii
Table des matières ... iv
Liste des tableaux ... vi
Liste des figures ... vii
Liste des abréviations ... ix
Introduction ... 1
Généralités sur le parasite Leishmania ... 1
Leishmania, le parasite ... 1
Leishmania, la maladie ... 2
Leishmania, le cycle parasitaire ... 4
Leishmania, les molécules de surface ... 5
Réponse immunitaire ... 8
Réponse immunitaire : La théorie du soi, non-soi ... 9
Réponse immunitaire : La théorie du danger ... 10
Le neutrophile ... 11
Neutrophiles et immunité : les granules du neutrophile ... 13
Neutrophiles et immunité : recrutement cellulaire ... 15
Le macrophage ... 17
La phagocytose ... 18
Peau et immunité ... 18
Peau et immunité : Les kératinocytes ... 20
Peau et immunité : Les cellules dendritiques ... 20
Peau et immunité : Les macrophages ... 21
Peau et immunité : Les lymphocytes T... 21
Leishmania et immunité ... 22
Neutrophiles et Leishmania ... 23
Macrophages et Leishmania ... 24
Lymphocytes T et Leishmania ... 25
Évasion de la réponse immunitaire ... 26
Traitements et prévention de la leishmaniose ... 27
Glycobiologie ... 29
Galectines ... 31
Galectines et immunité ... 33
Galectines et immunité : Neutrophiles ... 33
Galectines et immunité : Monocytes et macrophages ... 34
Galectines et immunité : Cellules dendritiques ... 36
Galectines et immunité : Lymphocytes T ... 37
Galectine-7 ... 38
Galectines et Leishmania ... 38
Matériel et méthodes ... 41
Souris ... 41
Galectine-3 recombinante ... 41
Parasites et cellules ... 43
Modèle d’infection du coussinet plantaire... 44
Modèle de la poche d’air... 45
Détection de la Gal-3 par Western Blot ... 45
Analyse des cytokines des échantillons de lavages de poche d’air par ELISA ... 46
Analyses statistiques ... 46
Résultats ... 47
Résultats connexes précédemment obtenus au laboratoire ... 47
Résultats obtenus au cours de ce mémoire de maîtrise ... 49
Implication de la Gal-3 dans la réponse immunitaire contre les souches LV39 et Friedlin du parasite Leishmania major ... 49
Clivage de la Gal-3 par les souches LV39 et Friedlin du parasite Leishmania major ... 51
Implication de la Gal-7 et de la Gal-9 dans la réponse immunitaire contre le parasite Leishmania : Expérience de la poche d’air ... 56
Implication de la Gal-7 et de la Gal-9 dans la réponse immunitaire contre le parasite Leishmania : Expérience d’infection du coussinet plantaire ... 58
Discussion ... 61
Implication de la Gal-3 dans le recrutement de neutrophiles lors d’infections avec le parasite L. major LV39. ... 61
Libération de Gal-3 dans la poche d’air lors de stimulations avec L. major LV39 ... 63
L. major LV39 et L. major Friedlin peuvent cliver la Gal-3, mais la cinétique de clivage est ralentie dans des conditions se rapprochant de ce qui est rencontré in vivo. ... 65
La Gal-7 inhibe le recrutement cellulaire induit lors de la stimulation avec L. major Friedlin ... 68
Les souris Gal-7 KO ont une phase de guérison plus rapide que les souris WT lors de l’infection du coussinet plantaire avec L. major Friedlin ... 70
La Gal-9 a un effet inhibiteur sur le recrutement cellulaire lors d’infection avec le parasite L. major Friedlin. ... 71
L’infection du coussinet plantaire avec L. major ne diffère que très peu d’un point de vue global entre les souris Gal-9 KO et les souris WT ... 72
Conclusion... 74
Liste des tableaux
Tableau 1: Classification des différentes espèces de Leishmania en fonction du séquençage du gène HSP70 ... 2 Tableau 2: Comparaison du nombre de répétitions de Galβ1-3 retrouvées sur les chaines
latérales du LPS de différentes souches et espèces de Leishmania. ... 7 Tableau 3: Les granules et vésicules sécrétoires du neutrophile ... 13
Liste des figures
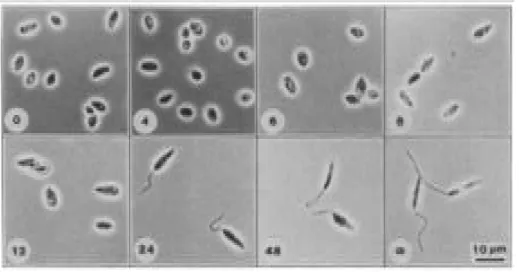
Figure 1 : Évolution morphologique du parasite Leishmania major en culture. La
transformation de l'amastigote vers le promastigote se fait en environ 24h (Schneider
et al., 1992). ... 1
Figure 2: Cycle parasitaire de Leishmania ... 4
Figure 3: Molécules de surface du promastigote Leishmania ... 5
Figure 4 : Structure du LPG de Leishmania ... 6
Figure 5: Clivage de la Gal-3 Visualisation sur gel d’acrylamide coloré au Bleu de Coomassie, d’une expérience de clivage de la Gal-3 ... 7
Figure 6 : Représentation schématisée de la réponse immunitaire selon la théorie du soi, non-soi ... 10
Figure 7 : Schématisation simplifiée de la réponse immunitaire selon la théorie du danger ... 11
Figure 8: Les différentes cellules immunitaires de l'épiderme et du derme humain ... 19
Figure 9: Représentation imagée de la théorie du cheval de Troie impliquant le neutrophile et le macrophage lors de l'infection avec Leishmania ... 23
Figure 10: Les souris C57BL/6 et BALB/c ont des réactions immunitaires différentes face au parasite Leishmania; la maladie progresse aussi différemment chez ce deux souches de souris ... 25
Figure 11: Photographie en microscopie électronique d'un marquage spécifique du glycocalyx ... 29
Figure 12: N-glycosylation et O-glycosylation chez les eucaryotes ... 29
Figure 13: Le contrôle de la glycosylation chez les mammifères ... 30
Figure 14: Les 3 familles de galectines et leurs membres respectifs ... 31
Figure 15: Rôles connus de différentes galectines au niveau des cellules immunitaires .... 33
Figure 16: Test d'hémaglutination des globules rouges effectué comme contrôle qualité des galectines produites dans notre laboratoire ... 42
Figure 17: Résultats obtenus suite à l'infection expérimentale de souris Gal-3 KO et WT avec le parasite Leishmania ... 47
Figure 18: Mesure de la quantité de neutrophiles dans le coussinet plantaire et dans le noeud lymphatique le drainant. Mesures effectuées à 12h et 24h post infection chez les souris Gal-3 KO et WT. ... 48
Figure 19: Recrutement cellulaire mesuré après 6h de stimulation lors d'une expérience de la poche d'air en utilisant L. major LV39 ou le LPS comme stimulant ... 49
Figure 20: Lors d'une expérience de la poche d'air, la seule injection de Gal-3 est suffisante pour induire un recrutement significatif après 6h ... 49
Figure 21: Comparaison des résultats obtenus lors d'une expérience de la poche d'air obtenus suite à l'injection de L. major LV39 et L. major Friedlin ... 50
Figure 22: Mesure, par Western blot, de la libération de Gal-3 de taille complète (anticorps Mac-2) par les macrophages (A) et les fibroblastes (B) suite à une stimulation in vitro de 3h par le parasite Leishmania ... 51 Figure 23: Mesure, par Western blot, du clivage de la Gal-3 par L. major Friedlin et L.
major LV39 dans des conditions d'incubation avec ou sans sérum (détection de la
forme non clivée de la Gal-3 avec l’anticorps Mac-2) ... 51 Figure 24: Détection, par Western blot, du clivage de la Gal-3 par L. major Friedlin et L.
major LV39 dans des conditions d'incubation avec ou sans sérum (détection du CRD
de la Gal-3 avec l'anticorps anti-CRD) ... 52 Figure 25: Contrôles utilisés pour les expériences de clivage de la Gal-3. ... 52 Figure 26: Détection, par Western blot, du clivage de la Gal-3 par L. major LV39 dans un
milieu d'incubation constitué de lavages de poches d’air murines (détection de la forme entière de la Gal-3 avec l'anticorps Mac-2) ... 54 Figure 27: Détection, par Western blot, du clivage de la Gal-3 par L. major LV39 dans un
milieu d'incubation constitué de lavage de poches d’air murines (détection du CRD de la Gal-3 avec l'anticorps anti-CRD) ... 55 Figure 28: Comptes à l'hémacimètre des leucocytes ayant migré dans la poche d'air suite à
une stimulation de 6h ... 56 Figure 29: Implication de la Gal-7 dans la migration des leucocytes dans la poche d'air
après une stimulation de 6h ... 57 Figure 30: Implication de la Gal-9 dans la migration des leucocytes dans la poche d'air
après une stimulation de 6h ... 58 Figure 31: Implication de la Gal-7 lors du suivi de la réponse inflammatoire locale des
souris infectées dans le coussinet plantaire avec L. major Friedlin ou L. major LV39 (suivi hebdomadaire de la différence de volume entre la patte infectée et la patte non-infectée) ... 59 Figure 32: Implication de la Gal-9 lors du suivi de la réponse inflammatoire locale des
souris infectées dans le coussinet plantaire avec L. major Friedlin et L. major LV39 (suivi hebdomadaire de la différence de volume entre la patte infectée et la patte non-infectée) ... 60 Figure 33: Illustration du pourcentage de LPG comportant le nombre de répétitions de
Galβ1-3 spécifié sus ses chaines latérales... 62 Figure 34: Schématisation du lien de la Gal-3 au LPG de L. major, puis de son clivage par
la GP63 du parasite ... 66 Figure 35: Schématisation du clivage de la Gal-3 par L. major puis de la taille des bandes détectées par Western blot en employant l'anticorps anti-CRD... ...67
Liste des abréviations
ATP : Adénosine-5'-triphosphateCD : Clusters de différenciation CLA : Cutaneous lymphocyte antigen
CMH : Complexe majeur d'histocompatibilité CRD : Domaine de reconnaissance des sucres
DAMPs : « Damage associated molecular pattern » Patrons moléculaires associés au danger
Gal : Galectine
GP63 : Leishmaniolysine
GPI : Glycosylphosphatidylinositol
HMGB1 : High-mobility group protein B1
SHP : Heat Shock Protein, protéine de shock thermique
KO : « Knock-out »; se dit d’un gène inactivé par génie génétique LPG : Lypophosphoglycan
LPS : Lipopolysaccharide
MAC : Complexe d’attaque membranaire du complément
Mac-2 : Désigne un Anticorps monoclonal reconnaissant la partie N-terminale de la galectine-3 humaine
NK : Natural Killer (se dit d’un type de lymphocyte)
PAMPs : « Pathogen-associated molecular patterns » Patron moléculaire associé aux pathogènes
PPG : Protéophosphoglycan
PRR : Pattern recognition receptors, récepteur des PAMPs TCR : Récepteur des cellules T
Th : Lymphocyte T helper (lymphocyte T auxiliaire) TLR : Toll like receptor
Introduction
Généralités sur le parasite Leishmania
Leishmania, le parasite
Leishmania est un genre de la famille des Trypanosomatidae. Ce parasite eucaryote est l’agent causal de la leishmaniose et est transmis grâce à un insecte : la mouche des sables. Deux morphologies distinctes caractérisent ce parasite : la forme extracellulaire du parasite, nommée promastigote, est mobile et reconnaissable par un parasite de forme allongée possédant un long flagelle et est retrouvée dans l'intestin de l'insecte tandis que la forme intracellulaire du parasite, nommée amastigote est non mobile et est quant à elle reconnaissable par sa plus petite dimension et par sa forme arrondie et elle est retrouvée à l'intérieur des macrophages de l’hôte infecté (Peters & Killick-Kendrick 1987) (Schneider et al. 1992) (Figure 1).
Figure 1 : Évolution morphologique du parasite Leishmania major en culture. La transformation de l'amastigote vers le promastigote se fait en environ 24h.
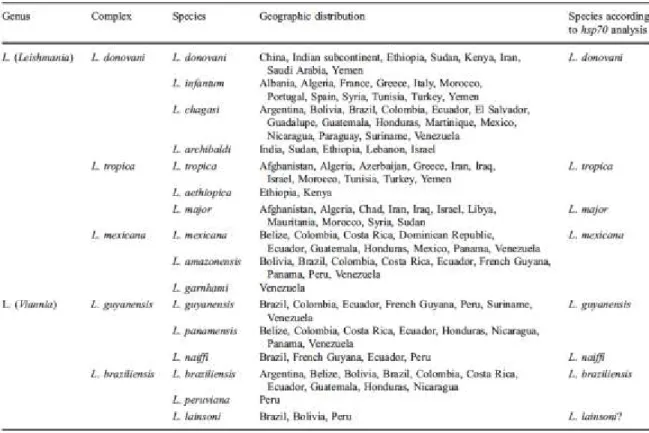
Différentes espèces du parasite vont causer différentes manifestations de la maladie. Trois types de leishmanioses sont connus; soit la forme viscérale, cutanée et mucocutanée. La classification taxonomique du parasite à été possible par l’emploi de différentes méthodes telles l’électrophorèse d’isoenzymes (Gardener et al. 1974), l’étude des caractéristiques de
croissance (Laison & Shaw 1972), la microscopie électronique (Gardener et al. 1977) ainsi que par l’utilisation d’anticorps monoclonaux (Pratt & J. R. David 1981). Une nouvelle classification basée sur la séquence du gène de la heat shock protein 70 a cependant été proposée récemment (Tableau 1) (Antinori et al. 2011). En tenant compte de ces résultats, les espèces L. tropica, L. major, L. mexicana, L. guyanensis, et L. naiffi causent la forme cutanée de la maladie, les espèces L. infantum et L. donovani causent la forme viscérale, tandis que L. brazillensis cause la leishmaniose mucocutanée.
Tableau 1: Classification des différentes espèces de Leishmania en fonction du séquençage du gène HSP70
Leishmania, la maladie
La forme cutanée de la maladie est la plus répandue avec environ 1.5 million de nouveaux cas répertoriés annuellement comparé à environ 500 000 cas pour la leishmaniose viscérale (Tiuman et al. 2011). Au total, environ 350 millions de personnes sont à risque de contracter la leishmaniose au cours de leur vie. Comme bien des maladies parasitaires, la leishmaniose est transmise par un insecte vecteur : la mouche des sables.
Leishmaniose cutanée
La distribution géographique de la leishmaniose cutanée est concentrée principalement dans les pays d’Amérique du Sud, d’Afrique du Nord et au Moyen Orient. La majorité des infections sont cependant rencontrées dans les pays suivants: Brésil, Pérou, Algérie, Syrie, Arabie Saoudite, Afghanistan et Pakistan,(Reithinger et al. 2007). La plupart des infections cutanées restent probablement asymptomatiques (Murray et al. 2005). Dans le cas où des symptômes apparaissent, le premier signe clinique est l’apparition d’un érythème à l’endroit où l’hôte a été piqué par la mouche des sables. La maladie se développera ensuite sur une période allant de deux semaines à six mois. Les rougeurs évolueront vers l’apparition d’une papule, puis d’un ulcère. L’infection va se résorber sans besoins d’interventions externes (sans traitement). La disparition des symptômes va se faire en un temps variable selon l’espèce de Leishmania dont il est question. Dans le cas de L. major, cela va se faire une période variable allant de quelques mois à quelques années (Alrajhi et al. 2002). Le fait de guérir de la maladie va procurer à l’individu une protection contre les prochaines infections. Dépendamment des cas, cette protection peut ou pas être spécifique à l’espèce de Leishmania qui est responsable de la primo-infection (passé en revue dans (Review & Reithinger 2008)).
Leishmaniose viscérale
La grande majorité des cas rapportés de leishmaniose viscérale proviennent des pays suivants : Bangladesh, Brésil, Éthiopie, Inde, Népal, Soudan (Reithinger 2008). Contrairement à la leishmaniose cutanée, la leishmaniose viscérale peut être fatale si non traitée. Les symptômes la caractérisant sont principalement la fièvre, l’hépatosplénomégalie (augmentation du volume du foie et de la rate), la lymphadénopathie (augmentation de volume des ganglions lymphatiques), la pancytopénie (réduction importante du nombre de globules blancs, de globules rouges et de plaquettes dans le sang), la perte de poids, une faiblesse généralisée et éventuellement, la mort (Reithinger, 2008).
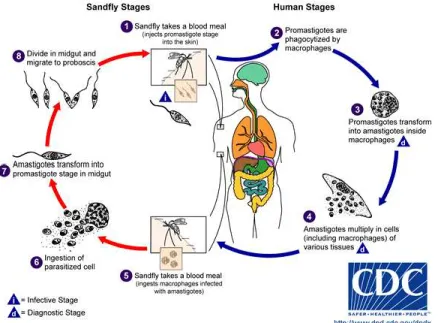
Leishmania, le cycle parasitaire
Deux stades principaux caractérisent le cycle parasitaire du parasite Leishmania (Figure 2). Une première partie de ce cycle se déroule chez le vecteur, la mouche des sables. Cet insecte appartient à la famille des Psychodidae. Deux principaux genres sont connus pour participer à la transmission du parasite soit le genre Phlebotomus rencontré dans les zones arides de l’Eurasie tandis que le genre Lutzomyia est surtout rencontré en Amérique du Sud (Killick-Kendrick 1990). Lorsque l’insecte pique un hôte infecté, il ingère des cellules infectées par des amastigotes.
Figure 2: Cycle parasitaire de Leishmania.
Ceux-ci vont par la suite se transformer en promastigotes et vont coloniser l’appareil digestif de l’insecte. Cette transformation est principalement induite par des changements environnementaux. Les parasites passent d’un environnement d’une température de 37°C qui pauvre en nutriment (le phagolysosome) vers un environnement d’une température de 26°C très riche en nutriments (le tube digestif de l’insecte) (Hart et al. 1981). Par la suite, ces promastigotes évoluent de la forme procyclique vers la forme métacyclique; cette dernière morphologie étant reconnue comme étant la forme infectieuse du parasite, dû à sa résistance au complément (Puentes & R. Da Silva 1990), qui est transmise à un hôte mammifère lors du prochain repas sanguin. Il est aussi possible de reproduire le phénomène de métacyclogénèse in vitro. Le fait de laisser les parasites de 6 à 10 jours leur
permet d’atteindre une phase de croissance stationnaire, obtenant alors un pourcentage significatif de parasites sous la forme métacyclique (Sacks, 1989). Finalement, à l’entrée du parasite dans les macrophages de l’hôte mammifère, la transformation des promastigotes en amastigotes est principalement médiée par une augmentation de température ainsi qu’une baisse de pH (Bates, 1993; Zilberstein and Shapira, 1994).
Leishmania, les molécules de surface
Les promastigotes du parasite Leishmania sont recouverts d’un glycocalix assez dense. La plupart des molécules le composant sont ancrées à la membrane cellulaire du parasite par le glycosylphosphatidylinositol (GPI). Des molécules de GPI sont aussi retrouvées seules; sans être reliées à une protéine (Figure 3) (McConville and Ferguson, 1993) (Ilgoutz and McConville, 2001).
Figure 3: Molécules de surface du promastigote Leishmania
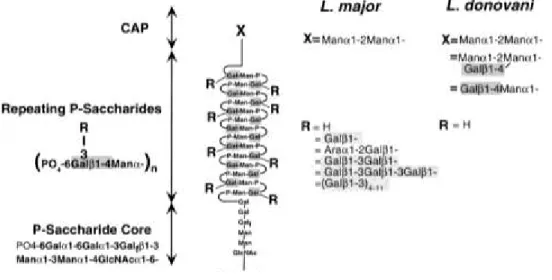
Le lipophosphoglycan (LPG) est une molécule de surface majeure du parasite Leishmania. Cette molécule structurelle est entre autres importante dans la survie du parasite face aux différents stress qu’il rencontre au cours de son existence (A. Descoteaux & Salvatore J Turco 2002) (Puentes & R. Da Silva 1990). La structure du LPG peut être divisée en quatre sections principales : i) l’ancrage à la membrane se faisant par le phosphatidylinositol (PI), ii) un cœur de polysaccharides, iii) des répétitions de polysaccharides et iv) une coiffe.
La partie qui nous intéresse le plus pour cette étude est la section composée de phosphosaccharides. La partie centrale de cette section est conservée à travers les différentes espèces de Leishmania. Les chaines latérales (R) sont cependant variables selon l’espèce rencontrée. En effet, lorsque l’on fait la comparaison des chaines latérales de L.
major et de L. donovani, on constate que celles de L. donovani sont uniquement composées
d’atomes d’hydrogène tandis que celles de L. major peuvent être composées de différents saccharides dont des répétitions de galactose reliées par des liaisons β1-3. C’est par cette différence structurelle qu’il est possible d’expliquer le fait que la Gal-3 lie L. major, mais pas L. donovani (la Gal-3 ayant une affinité pour les polymères de galactose liés entre eux par des liens β1-3; plus de détails à ce sujet dans la section glycobiologie). Il est possible d’en faire la preuve par l’usage d’une souche modifiée génétiquement de L. major Friedlin, nommée « Spock ». Cette souche n’est pas en mesure de lier la Gal-3 car, tout comme L.
donovani, elle est déficiente en β-galactosyltransférase, rendant ses les chaines latérales
identiques à celles de L. donovani (Figure 4) (Pelletier & S. Sato 2002).
Figure 4 : Structure du LPG de Leishmania.
Un détail important à propos des espèces de Leishmania utilisées dans le cadre de cette étude est ici à souligner. Deux souches de L. major ont été utilisées : L. major LV39 et L. major Friedlin. Ces deux souches causent la forme cutanée de la maladie. La caractéristique principale qui les distingue est le nombre de répétitions de Galactoseβ1-3 composant les chaines latérales de leur LPG (Tableau 2) (Dobson et al. 2010).
Tableau 2: Comparaison du nombre de répétitions de Galβ1-3 retrouvées sur les chaines latérales du LPS de différentes souches et espèces de Leishmania.
Souche de Leishmania
% du LPG contenant le nombre de répétitions du nombre d’unités de
(Galβ1-3) mentionné Moyenne du nombre de
0 1 2 3 4 5 6 7 8 9 répétitions
L. major Friedlin 21 59 11 1 0 0 0 0 0 0 1.2
L. major LV39 7 16 22 16 17 11 7 3 2 0 3.3
L. donovani 100 0 0 0 0 0 0 0 0 0 0
Une des protéines les plus abondantes retrouvées à la surface du parasite est la leishmaniolysine, aussi appelée GP63. Cette protéine est une zinc protéase (Novozhilova & N. V. Bovin 2010). Elle est exprimée chez les promastigotes ainsi que chez les amastigotes, mais en plus faible quantité chez ces derniers (Medina-Acosta et al. 1989). Cette protéase est connue pour hydrolyser des protéines de la matrice extracellulaire (P. B. Joshi et al. 2002) ainsi que certaines protéines du système immunitaire, dont des composants du système du complément (Brittingham et al. 1999). Des résultats obtenus précédemment dans notre laboratoire tendent à démontrer que la GP63 pourrait être à l’origine du clivage de la Gal-3 par les parasites Leishmania (Figure 5) (Pelletier et al. 2003).
Figure 5: Clivage de la Gal-3
Visualisation sur gel d’acrylamide coloré au Bleu de Coomassie d’une expérience de clivage de la Gal-3.
Il est cependant à noter que la Gal-3 doit premièrement lier le parasite via son LPG pour ensuite être clivée par la GP63. C’est pourquoi L. donovani, bien qu’il possède une GP63 fonctionnelle, ne clive pas la Gal-3.
Le protéophosphoglycan (PPG) est aussi présent à la surface des parasites et est impliqué dans la protection contre les enzymes protéolytiques (Secundino & Kimblin 2010). Le protéophosphoglycan est en fait un terme regroupant les protéines contenant une partie
phosphoglycan liée de manière covalente via les résidus sérines de la partie protéique. Le phosphoglycan du protéophosphoglycan de L. major, comporte, tout comme celui du LPG, les répétitions de polysaccharides Gal-Man-PO(4), une coiffe ainsi qu’un nombre important de répétitions de Galactoseβ1-3. C’est aussi grâce à ces dernières que le PPG est aussi être un ligand potentiel pour la Gal-3.
Réponse immunitaire
Le système immunitaire humain est composé de deux axes principaux : l’immunité innée et l’immunité adaptative. L’immunité innée va réagir aux infections en peu de temps et de manière non spécifique tandis que l’immunité acquise prend plus de temps à réagir, mais est plus spécifique et possède une capacité mémoire de manière à contrer plus efficacement une infection subséquente impliquant un pathogène ayant des caractéristiques immunologiques similaires. Deux modèles de pensée sont proposés afin d’expliquer le fonctionnement général du système immunitaire. Le modèle classique du soi et du non-soi proposé par Charles A Janeway expliquait initialement que le système immunitaire reconnait comme immunogène ce qui ne provient pas du soi (Medzhitov & Janeway 2002). Ce modèle s’est cependant adapté au fil du temps en ajoutant des éléments supplémentaires à ce principe de base (discutés plus loin). La théorie proposée par Polly Matzinger est quant à elle basée sur la dangerosité d’un évènement. Un évènement dangereux pour l’organisme est détecté par le fait qu’il entraine le bris de tissus ou de cellules. Ce bris occasionne la libération de molécules intracellulaires (comme HMGB1 ou l’ATP). Ces composés sont aussi appelés alarmines du fait qu’elles sont les responsables de l’activation de la réaction immunitaire (Matzinger 2002). La glycosylation fait aussi partie intégrale de différents mécanismes associés au système immunitaire. En effet, chez les vertébrés, les séquences de sucres ajoutés aux protéines se terminent souvent par un acide sialique. Cette glycosylation est essentielle pour différents processus de communication intercellulaire (Axford 2001). Des patrons de glycosylation spécifiques peuvent aussi être retrouvés chez certains pathogènes. Les lectines de type C, les galectines ainsi que les siglecs ont été décrits dans la littérature comme étant des PRRs (Dam & Brewer 2010). Les lectines de type C peuvent avoir différentes spécificités; par exemple, DC-SIGN reconnaît les glycans se terminant par
des résidus mannose et/ou fucose tandis que la « Macrophage galactose lectin » reconnaît les glycans se terminant par un résidu galactose ou N-acétylgalactosamine. Les siglecs sont quant à eux impliqués à la fois dans la reconnaissance des PAMPs présents sur certains virus, bactéries ou parasites, mais aussi de certaines molécules de l’hôte; facilitant ainsi a communication intercellulaire (Crocker et al. 2007). Finalement, certaines galectines ont démontré une capacité à lier des pathogènes; qu’ils soient d’origine bactérienne (Mey et al. 1996), virale (Lee 2007, St-Pierre et al. 2011, Ouellet et al. 2005) ou eucaryote (Pelletier, et al. 2003; Pelletier & S. Sato 2002). Ce lien à un pathogène peut cependant bénéficier davantage au pathogène en question qu’à son hôte (Mercier et al. 2008).
Réponse immunitaire : La théorie du soi, non-soi
Selon la dernière version de cette théorie, le système immunitaire est en mesure de réagir à 3 types d’évènements (Figure 6) (Medzhitov & Janeway 2002). La présence d’un « non-soi microbien » basée sur la reconnaissance de patrons moléculaires uniques aux microorganismes (ex. reconnaissance du LPS par le TLR-4). Ces molécules uniques sont appelées PAMPs pour Pathogen Associated Molecular Patterns. Ces patrons moléculaires sont donc reconnus par le système immunitaire via les PRRs pour Pattern recognition
Receptors. Parce que les PAMPs sont retrouvés à la fois sur les microorganismes
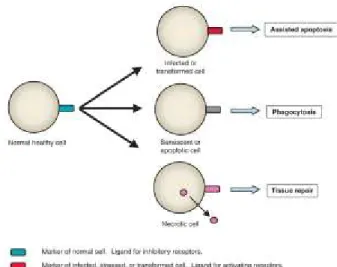
pathogènes et sur ceux qui ne le sont pas, ce système de reconnaissance ne peut cependant pas être la seule explication du fonctionnement de notre système immunitaire. En effet, le fait que notre corps soit en mesure de tolérer des microorganismes à certains endroits particuliers (système digestif, muqueuses) invalide le fait que les PAMPs soient la seule explication au fonctionnement de notre système immunitaire. Pour complémenter le système des PAMPS, le système immunitaire mise aussi sur la reconnaissance d’un « soi manquant » détecté par l’absence de marqueurs du soi normal (ex. l’activation des cellules NK par l’absence de CMH-1 à la surface d’une cellule). Finalement, la présence d’un « soi anormal » détectable par la présence de marqueurs induits lors d’une situation particulière (ex. l’exposition de molécules de phosphatidyl sérine par une cellule infectée par un virus va mener à l’élimination de cette cellule par phagocytose) est le troisième mécanisme d’activation de la réponse immunitaire (Medzhitov & Janeway 2002).
Figure 6 : Représentation schématisée de la réponse immunitaire selon la théorie du soi, non-soi.
Réponse immunitaire : La théorie du danger
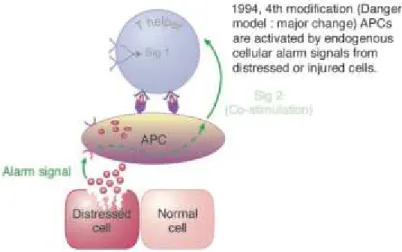
Bien que le modèle du soi, non-soi était présent dans le monde de la science depuis des nombreuses années, quelques questions étaient toujours ambigües. Par exemple, comment le système immunitaire fait pour s’adapter aux changements du soi lors de la puberté ou de la maternité? Comment les fétus sont tolérés ? Pourquoi il est nécessaire d’ajouter des adjuvants pour rendre les préparations vaccinales efficaces ? N’étant pas satisfaite des modèles précédents, c’est vers le milieu des années 90 que Polly Matzinger élabora une nouvelle théorie expliquant le fonctionnement du système immunitaire, la théorie du danger (Figure 7). Cette théorie se base sur le principe que le système immunitaire va réagir non pas aux molécules étrangères, mais plutôt au danger, signalé par des molécules de l’hôte nommées alarmines. Ces signaux de danger peuvent être d’origines variées, mais la clef est qu’ils sont produits selon deux possibilités : soit sécrétés par des cellules immunitaires stimulées ou libérés par la nécrose ou la mort non physiologique d’une cellule. Des molécules telles, les Heat-Shock proteins (HSP), la protéine HMGB1 (impliquée dans l’organisation de l’ADN) (Klune et al. 2008), les protéines S100 liant le calcium et retrouvées chez les cellules d’origine myéloïde (Foell et al. 2007) ainsi que l’ATP qui est une molécule intracellulaire connue comme coenzyme de bien des réactions
métaboliques (Kouzaki et al. 2011) sont des exemples d’alarmines souvent cités. Ces molécules vont, lorsqu’elles se retrouvent dans le milieu extracellulaire, activer les cellules présentatrices d’antigènes se trouvant à proximité. Pour être qualifiées d’alarmines, les molécules candidates doivent répondre a certains critères (Bianchi 2007). Elles doivent être (i) libérées dans l’environnement suite à la mort cellulaire non programmée de la cellule. (ii) Les cellules du système immunitaire peuvent être en mesure de sécréter ces molécules sans avoir à mourir. (iii) Ces molécules doivent être en mesure de recruter et d’activer les cellules immunitaires exprimant le récepteur correspondant afin de déclencher la réponse immunitaire. (iv) Les alarmines doivent finalement être en mesure de restaurer un état d’homéostasie en favorisant la reconstruction des tissus pouvant avoir été endommagés directement ou indirectement par la réponse immunitaire.
Figure 7 : Schématisation simplifiée de la réponse immunitaire selon la théorie du danger.
Toujours selon ce modèle, les cellules présentatrices d’antigène sont à la fois en mesure de reconnaitre les signaux endogènes et exogènes. Sont cités en exemple la reconnaissance de HSP70 et du LPS par le TRL-4 (Fang et al. 2011) ainsi que le lien du HSP60 et de différentes lipoprotéines microbiennes par le TLR-2 (Zanin-Zhorov & Cahalon 2006).
Le neutrophile
Chez l’homme, les neutrophiles sont les leucocytes les plus abondants du sang. Ce sont aussi un des premiers types cellulaires à être recrutés au site d’infection. En plus de leurs
nombreuses granules intracellulaires, les neutrophiles ont la capacité de phagocyter les pathogènes. Il est de plus en plus documenté que les neutrophiles jouent un rôle important dans l’activation et dans la régulation de la réponse immunitaire innée et adaptative (Mantovani et al. 2011). Les neutrophiles possèdent un arsenal de défense assez impressionnant. La plupart des molécules constituant cet arsenal se trouvent dans des granules retrouvées à l’intérieur du neutrophile. La dégranulation des neutrophiles se fait en plusieurs étapes ; ce ne sont pas tous les types de granules qui sont libérées en même temps. La cinétique de dégranulation est la suivante : (1) vésicules sécrétoires, (2) granules gélatinases, (3) granules spécifiques et (4) granules azurophiles (Sengelov 1993) (Sengelov et al. 1995)
Neutrophiles et immunité : les granules du neutrophile
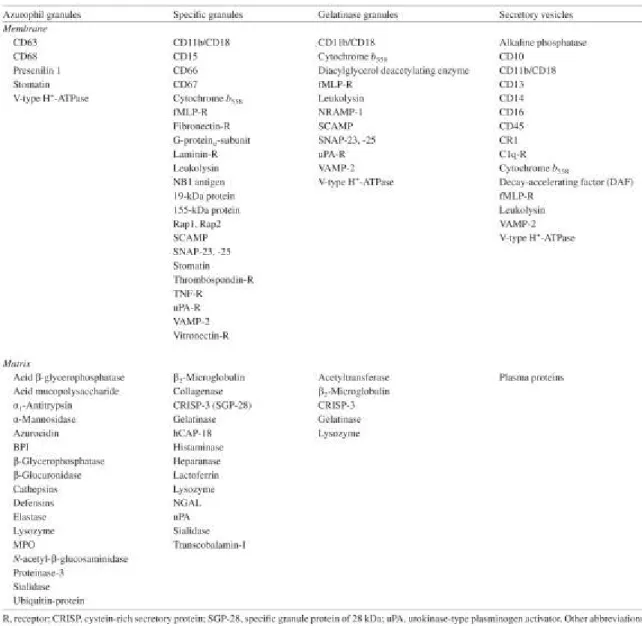
Tableau 3: Les granules et vésicules sécrétoires du neutrophile
Vésicules sécrétoires
Ces vésicules sont les premiers à être libérés. Une stimulation avec le fMLP ou avec l’ionomycine déclenche rapidement la libération d’albumine, un marqueur utilisé pour les vésicules sécrétoires. Aussi, contrairement à la libération des autres types de granules, la libération des vésicules sécrétoire est très peu affectée par la chélation du calcium extracellulaire. (Sengelov 1993b). La libération de ces vésicules provoque la libération de différentes molécules qui seront ensuite incorporées dans la membrane cellulaire. Parmi ces molécules, on retrouve la β2-integrin CD11b/CD18 (Mac-1) (Sengeløv et al. 1993) le récepteur 1 du complément (CR1) (Sengelov & Lars Kjeldsen 1994) ainsi que le récepteur du fMLP (Sengelov & Lars Kjeldsen 1994). À cela s’accompagne aussi l’exposition des molécules de L-sélectines, molécules impliquées dans le processus de migration des neutophiles (Bainton et al. 1994).
Granules gélatinases
Le contenu des granules gélatinases ne possède pas d’activité peroxydase. Leur contenu est plutôt composé d’enzymes capables de dégrader la matrice extracellulaire telles la gélatinase et l’héparinase (Mollinedo et al. 1997). Ces enzymes ont un rôle important à jouer dans la migration et l’extravasion des neutrophiles. Elles peuvent cependant aussi avoir des effets néfastes sur la santé lorsque leur niveau est mal contrôlé ; des maladies telles l’asthme et la fibrose kystique peuvent en résulter (Cederqvist et al. 2001). Les granules gélatinases diffèrent des granules spécifiques par le fait qu’elles ne contiennent pas de lactoferrine. En effet, les granules spécifiques (qui contiennent de la lactoferrine) contiennent aussi de la gélatinase (Segal 2005).
Granules spécifiques
Comme mentionné précédemment, les granules spécifiques contiennent de la lactoferrine ; un agent antimicrobien qui va lier et séquestrer le fer ; exerçant ainsi des propriétés antimicrobiennes pouvant agir sur un large spectre de bactéries à Gram-négatif ou positif (Oram & Reiter 1968). Ces granules contiennent aussi la majorité du lysozyme, une enzyme ayant une activité antibactérienne par sa capacité à hydrolyser le peptidoglycan (Baggiolini et al. 1969).
Granules azurophiles
Les granules azurophiles sont les dernières à êtres libérées dans le processus de dégranulation du neutrophile. Elles se distinguent des autres granules par leur activité peroxydase provenant de la myélopéroxidase une enzyme catalysant la production d’acide hypochloreux à partir de peroxyde d’hydrogène et de chlore. Cet acide hypochloreux ainsi que différents autres sous-produits de la réaction chimique vont attaquer les membranes des microorganismes par des réactions d’oxydoréduction (Klebanoff 2005). Il est aussi retrouvé dans ces granules l’enzyme élastase, qui, en plus pouvoir dégrader l’élastine (une composante de la matrice extracellulaire) (Sinha et al. 1987), possède la capacité de cliver la partie N-terminale de la Gal-3 (Nieminen et al. 2005). Finalement, les granules azurophiles contiennent aussi une variété de peptides antimicrobiens, dont les cathepsines et les défensines (Ganz 2003).
Neutrophiles et immunité : recrutement cellulaire
Lors d’une réaction inflammatoire, une série de mécanismes impliquant entre autres de nombreuses lectines, amènent les neutrophiles au site inflammatoire (Parham 2003). Cela commence par l’activation de l’endothélium vasculaire situé à proximité du site d’intérêt. Des médiateurs inflammatoires tels le leucotriène LTB4, le fragment C5a du complément ou l’histamine contribuent à l’exposition rapide de la P-sélectine, qui était préalablement
conservée dans les corpuscules de Weibel-Palade, à la surface des cellules endothéliales activées. Des molécules pro-inflammatoires telles le LPS et le TNF-α peuvent, quant à elles, provoquer l’expression de la E-sélectine à la surface des cellules endothéliales. Alors que l’expression de la P-sélectine peut se faire en quelques minutes, celle de la E-sélectine demande plutôt quelques heures. Ces sélectines vont lier des glycoprotéines contenant des structures osidiques de type sialyl-Lewis situées sur les neutrophiles et vont permettre une adhérence réversible des neutrophiles aux parois de l’endothélium. Cela permet donc aux neutrophiles de rouler sur la surface de l’endothélium vasculaire
La seconde étape de la migration des neutrophiles vers le site d’inflammation est l’adhésion ferme. Cela se fait par l’interaction entre les intégrines LFA-1 et Mac-1 du neutrophile avec les molécules d’adhérence cellulaire de l’endothélium tel ICAM-1. Lors du processus de roulement, les médiateurs inflammatoires tel l’IL-8 présents permettent de modifier la conformation des molécules LFA-1 et Mac-1 du neutrophile, augmentant ainsi la force de l’adhérence des neutrophiles aux parois endothéliales ; le roulement est ainsi stoppé pour laisser place à l’adhésion ferme.
S’en suit ainsi l’étape de migration du neutrophile à travers la paroi du vaisseau sanguin. En plus des molécules LFA-1 et Mac-1, la protéine CD31, une immunoglobuline exprimée sur les leucocytes ainsi que sur la paroi de l’endothélium vasculaire joue un rôle important dans ce processus. Le processus par lequel le neutrophile passe entre deux cellules endothéliales se nomme diapédèse. Le neutrophile peut alors atteindre la matrice extracellulaire, puis, grâce à un gradient de molécules chimiotactiques comme l’IL-8, le neutrophile se rend jusqu’au foyer du site inflammatoire.
Parce qu’elle a la capacité de lier les neutrophiles ainsi que les cellules endothéliales, il possible de croire que la Gal-3 ait un rôle à jouer dans ce processus de migration cellulaire (Nieminen et al. 2008).
Les chimiokines sont des cytokines ayant des propriétés chimioattractantes. Elles jouent un rôle primordial dans le processus de migration directionnelle des neutrophiles précédemment expliqué. Elles sont sécrétées par différents types cellulaires suite à une lésion cellulaire ou suite à la reconnaissance d’un pathogène. Il existe deux familles
principales de chimiokines qui sont classées selon la position de certains résidus cystéine (C) au sein de leur séquence d’acides aminés. Les chimiokines ayant deux cystéines consécutives (CC) sont connues pour attirer les monocytes ainsi que les lymphocytes T effecteurs et mémoire. Les chimiokines possédant deux cystéines intercalées par un acide aminé différent (CXC) sont quant à elles connues pour attirer les neutrophiles et les lymphocytes T naïfs.
Le macrophage
Selon la différenciation cellulaire qu’ils ont subit et selon leur localisation, les macrophages sont des cellules assez diversifiées. Les macrophages peuvent être des cellules résidentes des tissus (ex. dans la peau) ou bien être issus de la différenciation de monocytes ayant migré vers un site inflammatoire (Taylor et al. 2005). Leurs rôles au sein du système immunitaire peuvent aussi être assez diversifiés; ils sont entre autres connus pour leurs capacités phagocytaires, pour leur capacité à agir comme cellule présentatrice d’antigène ainsi que pour leurs rôles dans la modulation de la réponse immunitaire (Pozzi et al. 2005). Les antigènes rencontrés par les macrophages ainsi que les lymphocytes T se trouvant dans leur environnement vont influencer l’hétérogénéité ainsi que la maturation des macrophages (Van Ginderachter et al. 2006). Les macrophages qui répondent aux stimuli immunitaires par la sécrétion de molécules pro-inflammatoires et/ou microbicides sont considérés comme étant activés selon la voie classique (M1). Ces macrophages proviennent d’un environnement riche en cytokines provenant de cellules Th1 telles l’INF-γ et/ou contenant des PAMPs et/ou des DAMPs. Ce type de macrophage peut être reconnu par sa production élevée d’IL-12 (Verreck et al. 2004), par sa capacité proliférative et cytotoxique accrue ainsi que par sa production de cytokines pro-inflammatoires telles le TNF-α, l’IL-6 et l’IL-1 (Bonnotte et al. 2001). Le second type d’activation des macrophages est appelé activation alternative (M2). Ces macrophages ont plutôt des propriétés anti-inflammatoires qu’ils ont acquises suite à leur exposition à des cytokines Th2 telles l’IL4 et l’IL-13 ainsi qu’à d’autres molécules anti-inflammatoires telles l’IL-10, le TGF-β ainsi que les glucocorticoïdes (Goerdt & Orfanos 1999) (Verreck et al. 2004).
La phagocytose
Les neutrophiles ainsi que les macrophages sont considérés comme des phagocytes professionnels. Ils peuvent phagocyter tant les cellules mortes, les débris cellulaires que les pathogènes. La phagocytose est le processus par lequel l’élément à phagocyter pénètre à l’intérieur de la cellule par une vacuole provenant de l’invagination de la membrane cellulaire. Ce processus est possible grâce à la présence de différents récepteurs qui peuvent être séparés en deux classes : les récepteurs de type Fc tels FcγRIIA (CD32) et Fc γRIIIB (CD16) ainsi que les récepteurs du complément tels CR1 (CD35) et CR3 (les intégrines CD11b/CD18) (Witko-Sarsat & Rieu 2000). Suite à la reconnaissance d’une molécule à phagocyter, la cellule va réorganiser son cytosquelette afin de former des pseudopodes et une vacuole nommée phagosome. À ce moment, le phagosome ne contient aucune molécule microbicide, mais sa maturation va lui permettre de les acquérir. Le processus de maturation du phagosome commence par sa fusion avec les endosomes de tri (pH≈6.0). Cela permet l’acquisition de molécules essentielles au guidage du phagosome précoce ainsi formé à travers le cytoplasme. Par un processus encore mal caractérisé, le vésicule va fusionner avec les endosomes tardifs (pH≈5.5) afin de former le phagosome tardif. Ce phagosome tardif va finalement fusionner avec le lysosome (pH<5.5) qui contient des enzymes hydrolytiques comme la cathepsin D (O. V Vieira et al. 2002), formant ainsi le phagolysosome. Un second modèle fonctionnel nommé « Kiss and run » a aussi été proposé (Desjardins 1995). Bien que ce modèle suive les mêmes étapes que le modèle précédent, la différence principale se situe au niveau du processus de fusion entre le phagosome qui mature et les endosomes. Au lieu de proposer une fusion complète des deux vésicules, le modèle « Kiss and run » propose une fusion partielle et transitoire des membranes des deux vésicules qui serait suffisante pour leur permettre d’échanger leur contenu intravésiculaire.
Peau et immunité
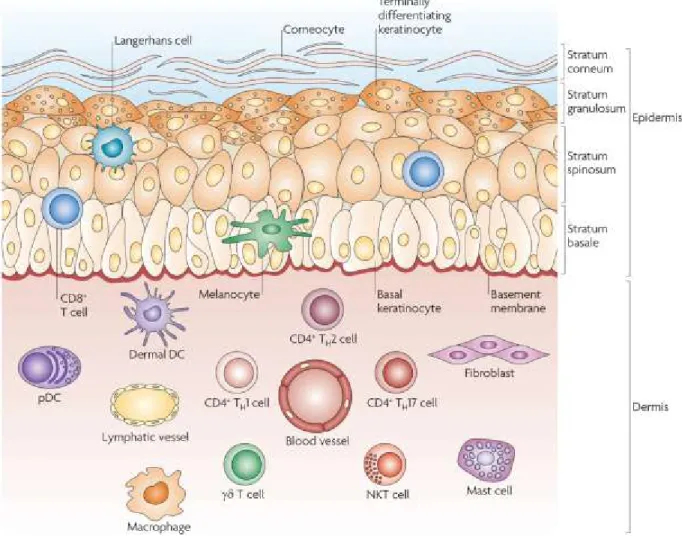
En plus de former une barrière physique et chimique contre le milieu extérieur, la peau possède aussi des défenses immunitaires biologiques très élaborées. Sont présentes dans les
différentes couches de la peau un nombre important de cellules dont certaines sont des cellules immunitaires spécialisées comme les lymphocytes T, les cellules dendritiques ainsi que les macrophages alors que d’autres comme les fibroblastes et les kératinocytes ont des rôles structuraux et immunologiques (Figure 8). Est aussi présent dans la peau un réseau lymphatique permettant aux cellules de migrer vers les organes lymphoïdes secondaires drainant cette région (Nestle et al. 2009).
Peau et immunité : Les kératinocytes
Les kératinocytes sont présents dans l’épiderme et sont une des premières cellules immunitaires à rencontrer les pathogènes. Ils sont en mesure de reconnaître les agents microbiens grâce aux nombreux TLRs qu’ils possèdent. Sont présents à leur surface les TLR1 (lipoprotéines bactériennes), TLR2 (lipoprotéines bactériennes), TLR4 (LPS), TLR5 (flagelline) et TLR6 (lipoprotéines bactériennes). Les TRL3 (ARN viral double brin) et TLR9 (ARN viral simple brin) sont aussi présents dans leurs endosomes (Lebre et al. 2007). Les kératinocytes sont reconnus pour diriger la réponse immunitaire vers une réponse de type Th1 (Miller & Modlin 2007). Cette généralité à été confirmée comme pouvant s’appliquer à la leishmaniose où des cytokines telles l’IL-12 étaient exprimées par les kératinocyes suite à une infection avec L. major (Ehrchen et al. 2010). Suite à l’activation de ces récepteurs, les kératinocytes peuvent réagir en sécrétant des molécules antimicrobiennes comme les β-défensines, la calprotectine ainsi que le lysozyme (Bando et al. 2007) (N. Hayashi et al. 2007). Les kératinocytes activés sont aussi connus pour produire des cytokines telles CXCL1 ainsi que CXCL8 (Albanesi et al. 2005). Il a finalement été publié que les kératinocytes peuvent agir comme cellules présentatrices d’antigènes grâce au fait qu’ils possèdent des CMH à leur surface.
Peau et immunité : Les cellules dendritiques
Les cellules de Langerhans sont localisées dans l’épiderme et sont les premières cellules dendritiques qui vont rencontrer les molécules antigéniques provenant du milieu extérieur. Étant une cellule présentatrice d’antigène professionnelle, son rôle principal est de capturer et de présenter les antigènes aux cellules T (Romani et al. 2006). Elles semblent aussi induire la différenciation des lymphocytes T vers le type Th2 (Klechevsky et al. 2008). Des cellules dendritiques sont aussi présentes au niveau du derme et ce sont ces cellules qui vont migrer vers les organes lymphoïdes secondaires adjacents à leur localisation. Une étude pouvant remettre en doute le rôle des cellules de Langerhans a démontré que seules les cellules dendritiques (pas les cellules de Langerhans) peuvent activer la prolifération des lymphocytes T auxquels elles présentent des antigènes. Cette étude a cependant été
faite dans le cadre de recherches sur les réactions d’hypersensibilité utilisant le Platelet-activating factor (PAF) comme stimulant (Fukunaga 2008). Il n’est pas impossible que l’emploi d’un stimulant différent ait donné des résultats différents. Des monocytes sont aussi présents au niveau du derme et ils peuvent maturer en cellules dendritiques ou en cellules de Langerhans (López-Bravo & Ardavín 2008). Il est finalement connu que les cellules dendritiques de la peau peuvent participer plus directement à la réponse immunitaire par la production de cytokines (Guttman-Yassky et al. 2007).
Peau et immunité : Les macrophages
Les macrophages présents dans la peau sont surtout retrouvés dans le derme, mais ils sont aussi connus pour leur capacité à migrer vers les nœuds lymphatiques drainant leur emplacement d’origine (Furth & Nibbering 1985).
Peau et immunité : Les lymphocytes T
Un nombre impressionnant de lymphocytes T se retrouve dans la peau. Environ 1E6 lymphocytes T par cm2 est présent dans une peau normale ; pour un grand total d’environ 2E10 lymphocytes T, ce qui est environ deux fois supérieur à ce qui est retrouvé dans la circulation sanguine (Schaerli et al. 2006). Les lymphocytes T peuvent être retrouvés à la fois dans l’épiderme (souvent près des cellules de Langerhans) (Foster & Yokozeki 1990) ainsi que dans le derme (Bos & Kapsenberg 1993). Des lymphocytes T CD4+ et CD8+ sont retrouvés en proportions équivalentes et la majorité d’entre eux sont des cellules mémoire. Elles ont aussi la particularité d’exprimer un antigène spécifique aux lymphocytes T de la peau : le « cutaneous lymphocyte-associated antigen » (CLA) (Nestle et al. 2009). Des cellules moins communes telles les lymphocytes T γδ (Shiohara & Moriya 1990) et Natural Killer (NKT) sont aussi retrouvées dans la peau.
Leishmania et immunité
Le parasite Leishmania entre dans la circulation sanguine sous la forme promastigote. Les promastigotes sont ensuite phagocytés pas les macrophages. Cette phagocytose est souvent facilitée par l’opsonisation des parasites avec la molécule C3b. La métalloprotéase de surface du parasite GP63 va ensuite cliver le C3b en C3i afin de favoriser la phagocytose du parasite au lieu de mener vers la voie lytique du complément (Hermoso et al. 1991). Il a aussi été démontré que les parasites sous leur forme stationnaire de culture in vitro ; forme correspondante au type de parasites injectés par la mouche des sables aussi appelée promastigotes métacycliques, sont plus résistants au système du complément que les parasites sous leur forme exponentielle de croissance en culture in vitro (Puentes & R. Da Silva 1990); forme correspondant aux parasites retrouvés dans les premières étapes de développement chez la mouche des sables et aussi appelée promastigotes procycliques. Ni les promastigotes, ni les amastigotes ne sont en mesure d’inhiber la fusion du phagosomes avec les endosomes tardifs ou avec les lysosomes (Courret et al. 2002). Il a cependant été démontré que le LPG peut inhiber de manière transitoire la maturation du phagosome afin de permettre aux promastigotes de se transformer en amastigotes qui sont plus résistants aux hydrolases (Dermine et al. 2000). L’emploi de parasites L. major déficients en LPG a confirmé ce résultat par l’obtention d’un taux de survie parasitaire inférieur à celui des parasites de type sauvage (Späth et al. 2000). Cette observation n’est cependant pas valide pour toutes les espèces de Leishmania. Les parasites de l’espèce L. mexicana déficients en LPG possèdent le même taux de survie face à la phagocytose par les macrophages lors que comparé avec les parasites de type sauvage (Ilg et al. 2001). En plus de modifier la régulation de la compartimentalisation intracellulaire, le parasite Leishmania est aussi en mesure de modifier certains patrons de signalisation intracellulaire de ça cellule hôte. Le meilleur exemple de cette modification est l’inhibition spécifique de la production d’IL-12. Cela se produit par l’inhibition de la signalisation de la voie JAK/STAT menant habituellement à l’activation du promoteur de l’IL-12p40 (McDowell MA and Sacks 1999). L. donovani, mais pas L. major affecte aussi la production d’IL-12p70 par les cellules dendritiques (Mcdowell et al. 2002). D’un point de vue symptomatique, cette découverte semble logique du fait que les cellules dendritiques sont activées dans le cas
d’une infection avec L. major ; participant ainsi à restreindre l’infection à une zone locale, ce qui n’est cependant pas le cas lors d’une infection avec L. donovani.
Neutrophiles et Leishmania
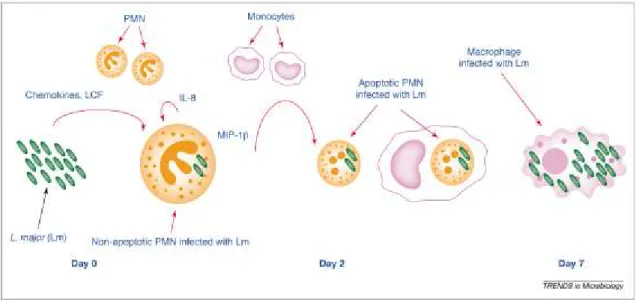
Comme mentionné précédemment, les neutrophiles sont parmi les premières cellules à être mobilisées au site d’infection suite à l’injection de parasites Leishmania. Ces neutrophiles seront en mesure de phagocyter et de tuer un fort pourcentage des parasites présents au site d’infection. Il a cependant aussi été démontré qu’un faible pourcentage des parasites internalisés à l’intérieur des neutrophiles peut survivre plusieurs heures (Helmut Laufs et al. 2002). Cela a mené à l’élaboration de l’hypothèse du cheval de Troie (Figure 9). Selon ce modèle, l’infection et la survie de parasites à l’intérieur des neutrophiles auraient comme effet d’occasionner un délai dans l’apoptose spontanée de ceux-ci. Ce délai permet aux monocytes recrutés au site d’infection de se transformer en macrophages qui vont pouvoir phagocyter les neutrophiles alors devenus apoptotiques. Cela va alors permettre aux parasites toujours présents à l’intérieur des neutrophiles de pouvoir infecter plus efficacement le macrophage qui ne s’attend pas a faire face à une cellule pathogène (Tamás Laskay et al. 2003).
Figure 9: Représentation imagée de la théorie du cheval de Troie impliquant le neutrophile et le macrophage lors de l'infection avec Leishmania.
Cependant, il faudrait mettre un certain bémol sur cette théorie. Les résultats obtenus au sein de notre laboratoire tendent plutôt à démontrer l’importance des neutrophiles dans la réponse immunitaire contre le parasite Leishmania. Nous avons en effet observé que les neutrophiles avaient une excellente capacité à phagocyter et à tuer les parasites. De plus, l’utilisation, pré-infection, d’anticorps afin d’éradiquer les neutrophiles à eu comme effet d’aggraver l’infection par l’augmentation de la charge parasitaire mesurée après 24h (Bhaumik et al. 2012).
Les neutrophiles sont plus que de simples éboueurs entrant en apoptose après avoir fait leur travail de nettoyage (phagocytose des parasites). Ils participent aussi à l’organisation de la réponse immunitaire. Il a entre autres été démonté que les neutrophiles sécrètent de l’IL-8 suite à leur exposition à des parasites in vitro (G Van Zandbergen et al. 2002). Ils sont donc susceptibles de participer à une cascade d’amplification de leur propre recrutement au site d’infection. Des cytokines comme l’IL-17 (Lopez Kostka et al. 2009) et le TNF-α (Q. Vieira et al. 1996) jouent aussi un rôle dans le recrutement des neutrophiles et le contrôle du développement de la maladie à plus long terme.
Macrophages et Leishmania
Les macrophages sont la principale cellule hôte des amastigotes. Selon leur type d’activation, ils seront plus ou moins permissifs à l’infection. Alors que les macrophages activés selon la voie classique (M1) sont en mesure de tuer efficacement les parasites, les macrophages activés selon la voie alternative sont permissifs à l’infection (Martinez et al. 2009) (Hölscher 2006) (B. H. W. Murray & Nathan 1999). Ce sont les intermédiaires oxygénés et nitrogénés réactifs qui participent à l’élimination des parasites (B. H. W. Murray & Nathan 1999). Les parasites du genre Leishmania ont développé des différentes tactiques afin de pouvoir survivre dans leur hôte, celles-ci passent principalement par l’inhibition ainsi que la modulation des gènes de leurs cellules hôtes : les macrophages (Buates & Matlashewski 2001; M. C. De Almeida et al. 2003; Hawn et al. 2002). Cette évasion de la réponse immune sera discutée plus en détail dans la section « Évasion de la réponse immunitaire » débutant à la page 26.
Lymphocytes T et Leishmania
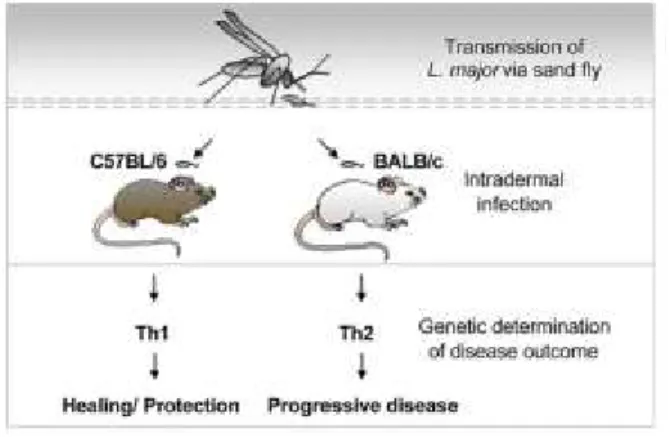
Figure 10: Les souris C57BL/6 et BALB/c ont des réactions immunitaires différentes face au parasite Leishmania; la maladie progresse aussi différemment chez ces deux souches de souris.
Du point de vue de l’immunité médiée par les lymphocytes T auxiliaires, les expériences d’infection avec L. major réalisées chez différentes souches de souris (Figure 10) ont permis de mettre en évidence l’importance d’une réponse immunitaire de type Th1 dans le contrôle de la charge parasitaire et éventuellement dans la guérison de la maladie (P Launois et al. 1997) (Himmelrich et al. 2000). Ce résultat est logique parce que la réponse immunitaire de type Th1 est connue pour être particulièrement efficace contre les pathogènes intracellulaires. Dans le cas du parasite Leishmania, cela se fait premièrement par la production d’IFN-γ par les lymphocytes T auxiliaires Th1 ainsi que par les lymphocytes cytotoxiques présents au site d’infection. L’IFN-γ est alors en mesure d’activer les macrophages afin qu’ils puissent produire des intermédiaires oxygénés et nitrogénés réactifs utilisés afin d’éliminer les parasites préalablement phagocytés (Liew et al. 1990) (Evans & Thai 1993). La susceptibilité aux infections avec L. major peut donc être associée à une réponse de type Th-2 impliquant la production de cytokines telles l’IL-4, IL-12, IL-10, toutes des signes de réponse Th-2 et antagonistes à la réponse Th-1 contribuant à l’inhibition de l’activation des macrophages (David Sacks & Noben-Trauth 2002). Bien que le modèle d’infection murin soit assez représentatif de ce qui se passe chez l’humain, la polarisation de la réponse immunitaire n’est cependant pas aussi franche chez l’humain. L’analyse de la réponse immunitaire par l’étude de la réponse cytokinaire des
lymphocytes T périphériques humains a démontré une réponse immunitaire mixte Th-1 et Th-2 (Barral et al. 2001). Comme pour le modèle murin, il a cependant été démontré chez l’homme que la résolution de la leishmaniose cutanée était liée à une réponse immunitaire de type Th-1 médiée par l’IFN-γ (Rogers et al. 2002).
Évasion de la réponse immunitaire
Les parasites protozoaires ont évolué afin de pouvoir évader la réponse immunitaire de leur hôte et ainsi pouvoir provoquer des infections chroniques chez celui-ci. Le parasite
Leishmania ne fait pas exception et a lui aussi quelques moyens à sa disposition afin
d’avoir un cycle infectieux le plus efficace possible (David Sacks & Sher 2002).
Le LPG du parasite à un grand rôle à jouer dans ces processus d’évasion du système immunitaire de l’hôte. Il a précédemment été démontré que les promastigotes sous leur forme métacyclique étaient plus résistants au système du complément que les promastigotes sous leur forme procyclique (Puentes & R. Da Silva 1990). Ce phénomène corrèle avec le fait que les chaines latérales du LPG sont plus longues lorsque le parasite est sous ça forme métacyclique (McConville et al. 1992) ; cela pourrait empêcher la formation du complexe d’attaque membranaire (MAC) par les protéines du complément.
Bien que le parasite Leishmania n’engendre pas de remodelage significatif du phagosome, (Courret et al. 2002) il semble qu’il soit à l’origine d’une inhibition transitoire de la maturation de celui-ci (Dermine et al. 2000). Cela permettrait au parasite de se différencier de sa forme promastigote à sa forme amastigote ; cette dernière étant plus résistante au pH acide ainsi qu’aux différentes enzymes hydrolytiques présentes dans le phagolysosome.
Le parasite Leishmania est aussi capable d’influencer la production de cytokines de son hôte à son avantage. Il est en mesure de diminuer l’efficacité de la réponse immunitaire par l’inhibition de la transcription du gène de l’IL-12p40 (McDowell & DL Sacks 1999). Sachant que l’IL-12 induit la production d’IFN-γ, une molécule particulièrement
importante dans le développement d’une immunité efficace contre les pathogènes intracellulaire, on comprend mieux pourquoi cette inhibition est favorable au parasite.
Traitements et prévention de la leishmaniose
Leishmania étant un eucaryote unicellulaire, les infections qu’il cause sont assez difficiles à
traiter du fait qu’il est phylogénétiquement plus proche des cellules humaines (comparé aux procaryotes, comme les bactéries par exemple). Les drogues traditionnellement utilisées pour le traitement de la leishmaniose sont l’antimoine pentavalent (dont le développement remonte à 1945), la pentamidine et l’amphotéricine B. Probablement due aux retombées financières peu encourageantes, le développement et l’étude de ces molécules ne semble pas être une priorité pour la plupart des entreprises pharmaceutiques. C’est probablement aussi ce qui explique le peu de nouvelles avancées dans ce champ d’étude ainsi que le fait que les mécanismes d’action des drogues préalablement mentionnées ne sont pas totalement identifiés. Il semble cependant que l‘antimoine pentavalent agit sur le métabolisme des purines, molécules essentielles composant l’ADN et l’ARN (Frézard et al. 2009). L’amphotéricine B fonctionne quant à elle par la formation d’un pore membranaire chez le parasite; déstabilisant ainsi le contenu ionique de son cytoplasme pour finalement mener à sa mort (Baginski & Czub 2009). Même si ces drogues sont acceptablement efficaces, elles ont de nombreux défauts dont la voie d’administration (intraveineuse), la durée du traitement (souvent de plusieurs semaines) ainsi que les effets secondaires néfastes pour l’organisme (fatigue, problèmes rénaux, cardiaques et pancréatiques) (Herwaldt 1999). Il est aussi important de noter que des cas de résistance à ces différentes molécules ont été observés en clinique (Croft et al. 2006) (Ouellette et al. 2004).
D’un point de vue vaccinal, aucune solution vraiment efficace n’a encore été mise au point. Sachant qu’une infection cutanée guérie confère à son hôte une protection contre les infections subséquentes du même type, il est possible de croire qu’une préparation vaccinale efficace pourrait avoir les mêmes effets protecteurs. Différentes stratégies vaccinales sont présentement au stade de développement et se basent sur différents
principes. Certains croient que l’induction d’une forte réponse immunitaire contre la salive de l’insecte pourrait engendrée une forte réponse immunitaire au site de la piqûre et ainsi contraindre le parasite. D’autres croient plutôt à une approche vaccinale plus classique composée de parasites tués et d’ADN du parasite; cette dernière composante servant à l’activation des lymphocytes T CD8+, cellules importantes dans la résistance aux infections causées par des parasites intracellulaires (Okwor & Uzonna 2009) (P. M. Kaye & Aebischer 2011).
La prévention n’est pas non plus à négliger. C’est un moyen efficace et peu couteux de ralentir la propagation de la maladie. Cette prévention passe essentiellement par le contrôle du vecteur (la mouche des sables) ainsi que des réservoirs animaux. Le contrôle du vecteur est entre autres effectué par l’usage d’insecticides dans les endroits les plus à risque (Clive R Davies et al. 2003) ainsi que par l’emploi de filets recouvrant le lit et la personne qui s’y trouve. Une étude réalisée au Népal a prouvé l’efficacité de cette dernière méthode en démontrant une réduction des incidences de leishmaniose viscérale de 70% (Bern et al. 2000). Pour ce qui est du contrôle des réservoirs animaux, une étude réalisée au Brésil à démontré des résultats prometteurs qui ont cependant été remis en perspective dans une seconde étude (Ashford et al. 1998) (Gavgani et al. 2002). Des études utilisant des insecticides directement sur les chiens ont aussi été réalisées. Bien qu’elles ont démontré des résultats intéressants, il est à se questionner au niveau de la mise en application quotidienne de ce genre de mesures préventives.
Glycobiologie
Chez les mammifères, les sucres recouvrent la surface de toutes les cellules (Figure 11). Parce qu’ils sont les éléments situés principalement aux extrémités des cellules, ces sucres formant le glycocalyx sont impliqués dans plusieurs mécanismes cellulaires.
Figure 11: Photographie en microscopie électronique d'un marquage spécifique du glycocalyx.
C’est dans l’appareil de Golgi qu’a lieu le processus de glycosylation des protéines. Cela se produit grâce à de nombreuses enzymes ; entre autres les glycosyltransférases qui catalysent le transfert des saccarides de la molécule donneuse vers la protéine à glycosyler ainsi que les glycosidases qui catalysent l’hydrolyse des liens glycosidiques au sein d’une structure de sucres complexes (Van Kooyk & Gabriel a Rabinovich 2008) (Ohtsubo & Marth 2006). Il existe deux types de glycosylation chez les mammifères ; leur différenciation est principalement basée sur la séquence protéique sur laquelle la glycosylation est amorcée.
Les sucres N-liés sont reconnaissables par leur séquence riche en résidus mannose et par le fait qu’ils sont liés à la protéine par le biais d’un résidu asparagine présent dans la séquence ASN-X-Ser/Thr où le X représente un acide aminé quelconque. Les sucres O-liés sont quant à eux reliés à des résidus sérine, thréonine ou hydroxylysine sans regard aux séquences adjacentes (Figure 12).
La formation des glycans cellulaires peut être régulée de multiples façons (Figure 13) (Ohtsubo & Marth 2006) que ce soit par : (i) le contrôle de la transcription des gènes des glycosyltransférases et des glycosidases, (ii) la synthèse et le transport des nucléotides de sucres qui agissent comme donneurs de sucres, (iii) la modulation par phosphorylation des enzymes impliquées dans les processus de glycosylation, (iv) la compétition enzyme/substrat à l’intérieur du Golgi, (v) la modulation du transport intracellulaire des enzymes, (vi) la protéolyse des enzymes pouvant avoir lieu dans le Golgi ainsi que par (vii) le renouvèlement des glycoprotéines par endocytose à la surface cellulaire.
Galectines
Les galectines sont des protéines retrouvées chez un large spectre d’êtres vivants en passant des vertébrés (poissons, oiseaux, amphibiens et animaux) jusqu’aux protistes (champignons) sans oublier les invertébrés comme les insectes et les vers (D. N. W. Cooper & Barondes 1999). Quinze galectines sont jusqu’ici connues chez les animaux (Figure 14). D’un point de vue structural, elles possèdent toutes un domaine de reconnaissance des sucres (CRD). Selon leur structure, les galectines sont classées en 3 familles distinctes. Les galectines 1, 2, 7, 11, 13 et 14 sont classées dans la famille des prototypes, car elles ne sont composées que d’un seul ou de deux CRD sans aucune autre structure protéique. La Gal-3 est quant à elle le seul membre de la famille des galectines chimériques. Elle a la particularité de posséder une queue protéique N-terminale lui permettant, suite à la liaison à son ligand, de former des oligomères. Finalement, les galectines 6, 8, 9 et 12 sont classés dans la famille des tandems répétés, car elles sont composées de 2 CRD (pas toujours identiques) liés ensemble par une région peptidique (Nieminen et al. 2007).
Figure 14: Les 3 familles de galectines et leurs membres respectifs.
Les galectines sont des lectines c’est-à-dire qu’elles sont des protéines ayant une affinité pour les sucres. Les galectines ont une affinité spécifique pour les saccarides de type