La dynamique du zooplancton au lac Saint-Jean
Texte intégral
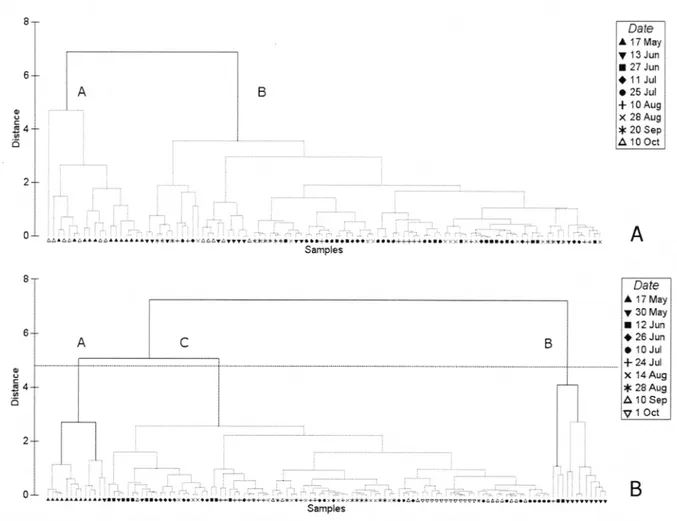
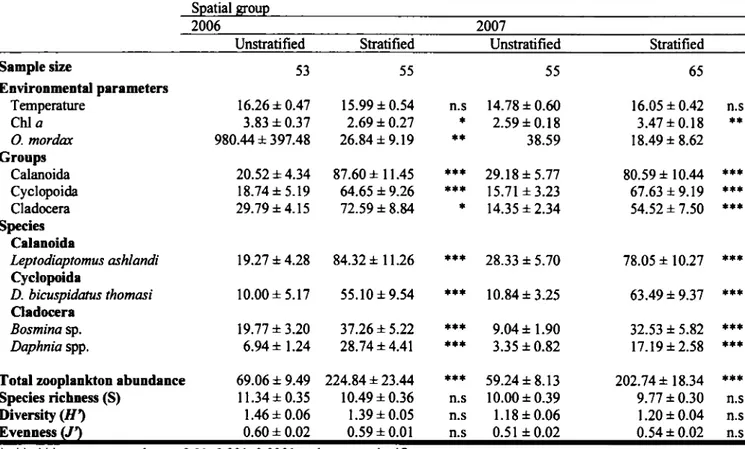
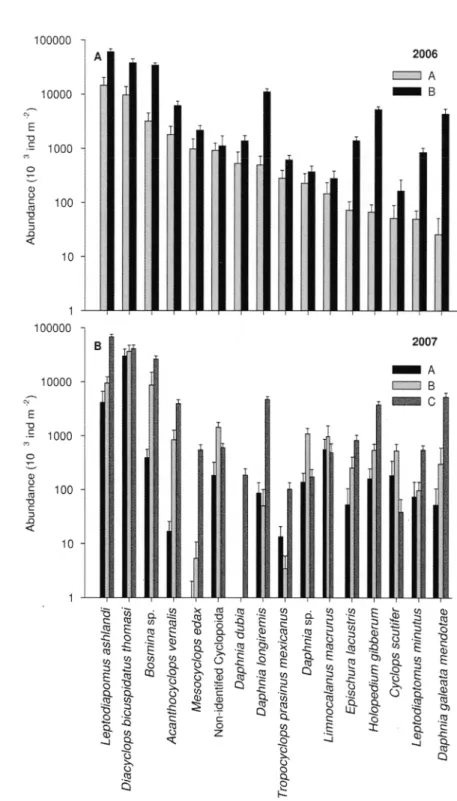
Figure
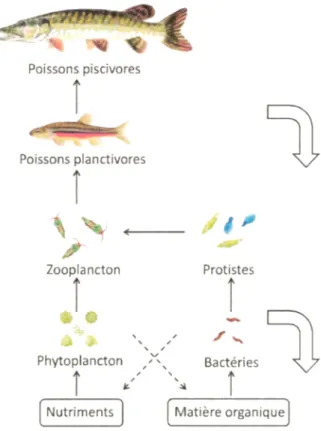
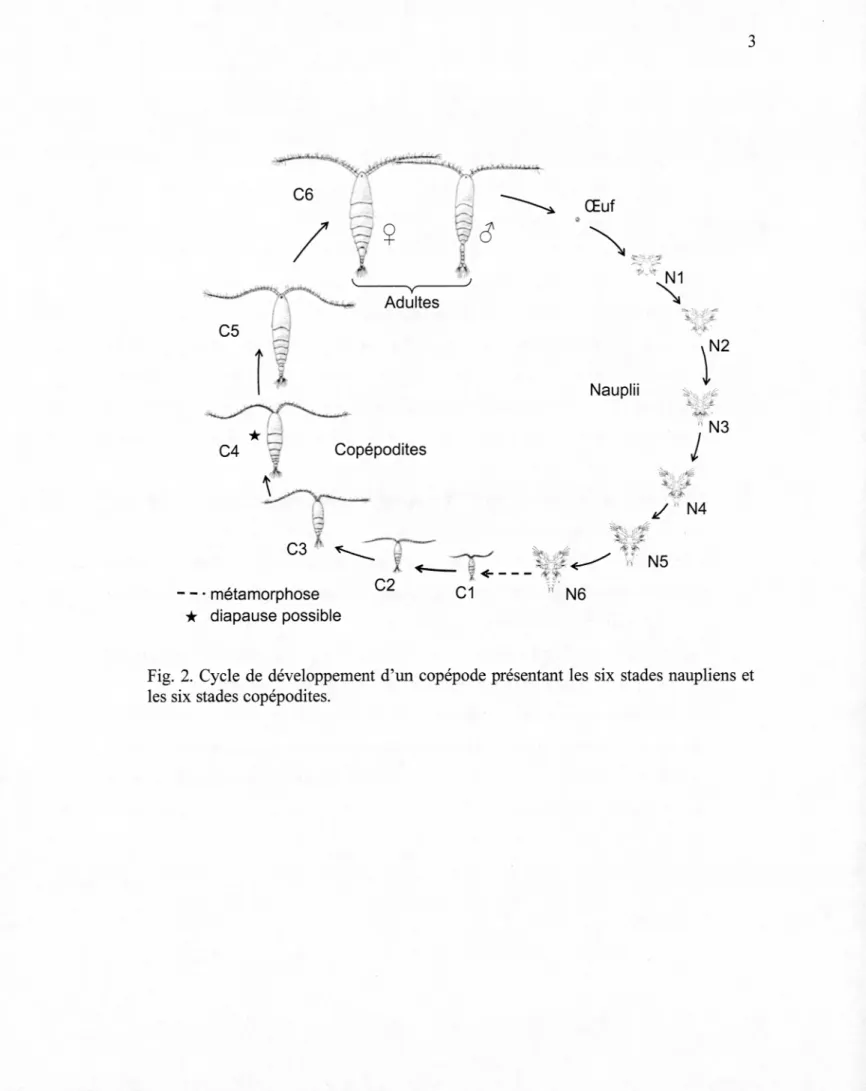

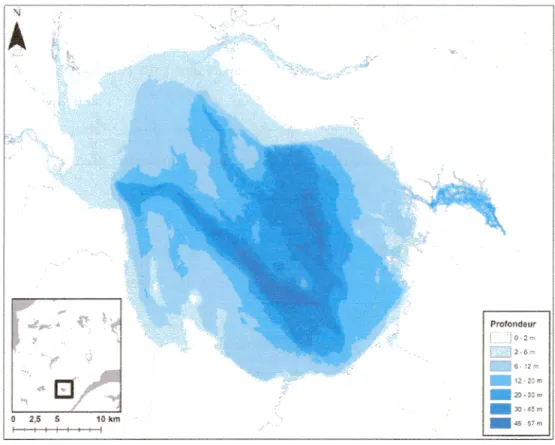
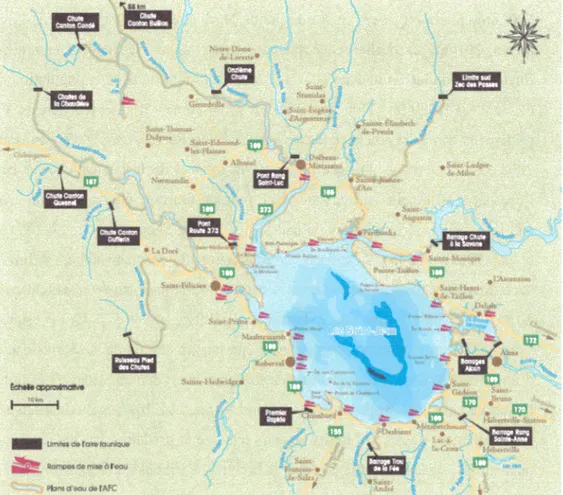


Documents relatifs
détroit d ' Hudson , ces informations deviennent cruciales car elles permettent de retracer avec précision la dynamique de la calotte de glace des Laurentides.. les
Dans chacune des stations d’échantillonnage appartenant aux types de peuplements associés au pin gris et à l’épinette noire, 45 charbons de bois, dont 25 proviennent de
Après la mise en œuvre des remblais, le contrôle de la qualité du compactage peut être réalisé au pénétromètre dynamique à énergie variable PANDA (*) dans le cadre
Comme les informations fournies par la bande 5 seule sont suffisantes pour étudier la répartition spatiale des solides en suspension et que l'ob- tention de ces
Dans le cadre de la planification de ses activités pour la saison 2011-2012, la Forêt modèle du Lac-Saint- Jean (FMLSJ) a réalisé des inventaires ayant pour objectif de caractériser
Pour la population de 22 municipalités du Saguenay–Lac-Saint-Jean, le transport interurbain par autocar est le seul mode de transport collectif, si l’on excepte bien sûr le
Notamment dans le tiers des raisons portant sur l’abondance du doré, par exemple « ça a déjà été mieux » ou encore « je réussis à capturer du doré à chaque sortie,
Ce guide est le premier réalisé sur l’identification du zooplancton et des invertébrés présents dans les rivières artificielles de l’installation expérimentale
