ETUDE DE LA SYMBIOSE ACTESfORHIZIENNE CHEZ L'AULNE RUGUEUX ET L'AULNE CRISPE COLONISANT LES SITES PERTURBES PAR L'INDUSTRIE
PETROLIERE ALBERTAINE
par
Cyntia Bissonnette
memoire presente au Departement de biologie en vue de l'obtention du grade de maitre 6s sciences (M.Sc.)
FACULTE DES SCIENCES UNIVERSITE DE SHERBROOKE
ill
Library and Archives Canada Published Heritage Branch 395 Wellington Street Ottawa ON K1A0N4 Canada Bibliothdque et Archives Canada Direction du Patrimoine de l'6dition 395, rue Wellington Ottawa ON K1A 0N4 CanadaYour file Votre r6f6rence ISBN: 978-0-494-83689-7 Our file Notre r6f6rence ISBN: 978-0-494-83689-7
NOTICE:
The author has granted a non
exclusive license allowing Library and Archives Canada to reproduce, publish, archive, preserve, conserve, communicate to the public by
telecommunication or on the Internet, loan, distrbute and sell theses
worldwide, for commercial or non commercial purposes, in microform, paper, electronic and/or any other formats.
AVIS:
L'auteur a accords une licence non exclusive permettant £ la Biblioth&que et Archives Canada de reproduire, publier, archiver, sauvegarder, conserver, transmettre au public par telecommunication ou par I'lnternet, pr§ter, distribuer et vendre des thdses partout dans le monde, d des fins commerciales ou autres, sur support microforme, papier, 6lectronique et/ou autres formats.
The author retains copyright ownership and moral rights in this thesis. Neither the thesis nor substantial extracts from it may be printed or otherwise reproduced without the author's permission.
L'auteur conserve la propri6t6 du droit d'auteur et des droits moraux qui protege cette these. Ni la th&se ni des extraits substantiels de celle-ci ne doivent §tre imprim&s ou autrement reproduits sans son autorisation.
In compliance with the Canadian Privacy Act some supporting forms may have been removed from this thesis.
While these forms may be included in the document page count, their removal does not represent any loss of content from the thesis.
Conform6ment d la loi canadienne sur la protection de la vie priv6e, quelques formulaires secondares ont 6t§ enlev6s de cette th6se.
Bien que ces formulaires aient inclus dans la pagination, il n'y aura aucun contenu manquant.
Le 20 janvier 2012
le jury a accepte le memoire de Madame Cyntia Bissonnette dans sa version finale.
Membres du jury
Professeur Sebastien Roy Directeur de recherche Departemenf de biologie
Professeure Carole Beaulieu Membre
Departement de biologie
Professeur Ryszard Brzezinski President rapporteur Departement de biologie
SOMMAIRE
Sous la foret boreale albertaine, dans l'Ouest canadien, repose la deuxieme plus grande reserve de petrole mondiale. Son exploitation detruit l'ecosysteme, en plus de generer de grandes quantites de residus phytotoxiques, dont les principaux sont le sable residuel (TS) et le coke. Selon la loi provinciale, avant la fin de 1'exploitation, les terres doivent etre restituees dans leur etat original. Cependant, la rehabilitation des terres demeure un defi significatif compte tenu du volume de residus qui seront generes par l'industrie petroliere en Alberta.
Les plantes actinorhiziennes sont des vegetaux tres resistants, qui colonisent souvent des habitats hostiles : depots volcaniques, dunes de sable, sols erodes a la suite d'un glissement de terrain, sols contamines. La plupart de ces milieux sont pauvres en azote, en consequence, l'etablissement de la symbiose avec un actinomycete fixateur d'azote atmospherique, Frankia spp., leur confere un avantage indeniable. Parmi les plantes actinorhiziennes, l'aulne actinorhize revet un interet particulier. A maintes reprises, il a prouve son utilite dans le cadre de diverses pratiques agroforestieres ou de rehabilitation de sites contamines par des metaux lourds, des residus chimiques et des hydrocarbures. Des 47 especes d'aulnes distribuees mondialement, 7 sont indigenes du Canada, dont l'aulne rugueux et l'aulne crispe. Puisque ces deux especes ont deja et6 utilisees a grande echelle au Canada pour rehabiliter des terres perturbees, leur emploi dans le cadre de la revegetalisation des sites albertains est pertinent.
Ce projet de recherche en microbiologic environnementale avait comme objectif d'evaluer la capacite des aulnes symbiotiques a coloniser directement les principaux residus de l'industrie petroliere albertaine. II tentait 6galement d'evaluer l'impact qu'auraient ces residus sur la symbiose de l'aulne crisps et de l'aulne rugueux avec Frankia. L'hypothSse de recherche stipule done que l'aulne crisps et l'aulne rugueux peuvent croitre directement en TS et en
coke, et que la symbiose avec la bacterie du genre Frankia ameliore le developpement. des aulnes croissant dans ces residus.
Afin d'investiguer la question, des aulnes crispes et rugueux ont ete cultives en conditions gnotobiotiques, en absence ou en presence de Frankia, dans du TS ou du coke. Par la suite, ils ont ete recoltes, et leur developpement a 6t6 lvalue. Que ce soit en presence de TS ou de coke,
Frankia a etabli la symbiose avec l'aulne crispe et l'aulne rugueux. Cette symbiose a permis
aux aulnes de survivre en substrats residuels et a assure un meilleur developpement de ceux-ci. Dans le cas du coke, une etude terrain a confirm^ ces r^sultats. En conclusion, l'aulne rugueux et l'aulne crispe detiennent les qualites necessaires pour participer a la revegetalisation des sites perturbes par l'industrie petroliere albertaine, et la presence de la symbiose actinorhizienne ameliore significativement le developpement de ces vegetaux en presence de stress environnemental.
Mots-cles: Frankia, Aulne, Symbiose actinorhizienne, Revegetalisation, Sables bitumineux, Coke
REMERCIEMENTS
J'aimerais tout d'abord remercier mon directeur de recherche, le professeur Sebastien Roy, pour m'avoir accueillie dans son laboratoire et m'avoir guidee tout au long de mon cheminement. Ses precieux conseils et son soutien lors des embuches rencontrees ont ete grandement apprecies. Je tiens 6galement a offrir mes remerciements a mes conseillers de maitrise, les docteurs Carole Beaulieu et Ryszard Brzezinski, pour leurs idees et leurs commentaires constructifs au sujet de mon projet.
Merci a mes collegues de laboratoire pour leur soutien technique et leur avis scientifique; leur aide a ete precieuse. Je ne peux non plus passer sous silence la participation de plusieurs amis et collogues ecologistes, dont Bill Parsons, qui m'ont eclairee a travers les meandres des domaines avec lesquels j'etais moins familiere. Je tiens aussi a souligner la contribution du professeur Robert Bradley, qui m'a g^nereusement prete son chromatographe en phase gazeuse, ainsi que celle des autres laboratoires ayant prete 1'equipement m'ayant aidee a mener mon projet a terme. Merci egalement a mon amie Maggie Fontaine pour avoir revise les sections redigees en fran9ais.
Je me dois enfin de mentionner le support financier du Fond de la Recherche en Sante du Quebec, par le biais d'une bourse de maitrise en recherche.
Finalement, merci a ma famille et k tous les amis qui ont eu foi en mes capacites. Un merci tout special h Alexandre Henry qui, en plus d'avoir participe h 1'edition des photos et la revision du manuscrit, m'a accompagnee et soutenue tout au long de mon parcours universitaire.
TABLE DES MATIERES
SOMMAIRE I
REMERCIEMENTS ; III
TABLE DES MATIERES IV
LISTE DES ABREVIATIONS VIII
LISTE DES TABLEAUX X
LISTE DES FIGURES XI
INTRODUCTION 1
1. Problematique etudiee : les sites perturbes par Pindustrie petroliere albertaine 1
1.1. L'exploitation des sables bitumineux 1
1.2. Les sites perturbes et les efforts de rehabilitation 3
2. Les organismes etudies 5
2.1. Frankia spp 6
2.2. Les plantes actinorhiziennes 12
2.3. La symbiose entre l'aulne et Frankia 16
2.3.1 Le processus de nodulation 16
2.3.2 Les facteurs influensant la nodulation 19
2.3.3 Les avantages procures par la symbiose actinorhizienne 21
3. Les aulnes : leur utilisation ant^rieure et actuelle 23
CHAPITRE 1
SYMBIOSIS WITH FRANKIA SP. BENEFITS THE ESTABLISHMENT OF ALNUS
VIRIDIS SSP. CRISPA AND ALNUS INCANA SSP. RUGOSA IN TAILING SANDS
FROM THE CANADIAN OIL SANDS INDUSTRY 27
1. Reference de l'article et contribution de l'auteur 27
2. Originalite du travail et contribution k la science 27
3. Resume de l'article en franfais 28
4. Manuscrit de l'article 29
4.1. Abstract 29
4.2. Introduction 30
4.3. Material and methods 32
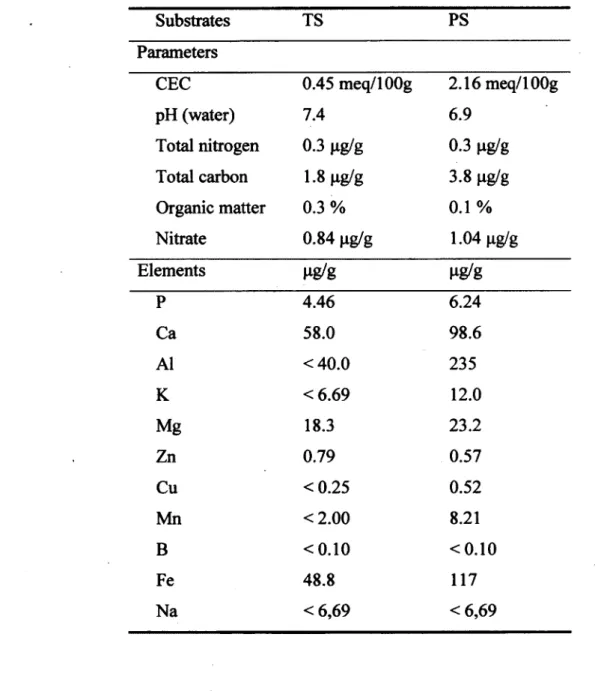
4.3.1. Growth substrate 32
4.3.2. Seedling treatment and experimental setup 34
4.3.2.1. Seed germination 34
4.3.2.2. Plant culture 34
4.3.3. Harvest and samples processing 36
4.3.4. Tissue and substrate analyses 36
4.3.4.1. pH and electrical conductivity 36
4.3.4.2. Naphthenic acids analyses in plant tissue 37
4.3.5. Statistical analyses 38
4.4. Results 38
4.4.1. Frankia colonization of alders ...38
4.4.2. Biomass production 38 4.4.1. Plant morphology 41 4.4.2. NA scan . 43 4.5. Discussion 47 4.6. Acknowledgements . 51 4.7. References 52 CHAPITRE 2
ACTINORHIZAL AND ECTOMYCORfflZAL SYMBIOSES ENHANCE ALDER GROWTH IN THE PRESENCE OF WEATHERED COKE FROM THE CANADIAN
OIL SANDS INDUSTRY 58
1. Reference de l'article et contribution de l'auteur 58
2. Originalite du travail et contribution a la science 58
3. Resume de l'article en frangais 59
4. Manuscrit de l'article 60
4.1. Abstract 60
4.2. Introduction 61
4.3. Material and methods 63
4.3.1. Growth substrate 63
4.3.2.1. Seed germination 65
4.3.2.2. Plant culture 65
4.3.3. Harvest and sample processing 66
4.3.4. Field trial 67
4.3.5. Statistical analyses 67
4.4. Results 68
4.4.1. Alder growth and actinorhizal symbiosis development 68
4.4.2. Plant morphology 70 4.4.3. Field experiment 73 4.5. Discussion 75 4.6. Acknowledgements 79 4.7. References 79 CONCLUSION 84 BIBLIOGRAPHIE 87 vii
LISTE DES ABBREVIATIONS
% pourcentage
°C degre Celcius
16S ARN ribosomal (vitesse de sedimentation de 16 Svedberg) ADN acide desoxyribonucl&que
AN acide(s) naphtenique(s) ARN acide ribonucleique ATP adenosine triphosphate
C cytosine
cm centimetre
EDTA acide ethylene diamine tetra acetique
G guanine
g gramme
g/1 gramme par litre
ha hectare (10 000 m2) kg kilogramme km kilometre h heure 1 litre M molaire mg milligramme min minute ml millilitre mm millimetre mM millimolaire pH potentiel hydrogene
ppm partie par million (mg/1) sp. espece (lorsque non definie)
spp. especes (ensemble des)
ssp. sous-espece
TS sables residuels (tailing sand)
(j.1 microlitre
^ig microgramme
LISTE DES TABLEAUX
Tableau 1. Liste des families vegetales actinorhizees par les souches de Frankia selon le
groupe d'infection auquel elles appartiennent 9
LISTE DES FIGURES
Figure 1. Principaux residus generes par l'industrie p&roliere aibertaine 4
Figure 2. Arbre phylogenetique illustrant la position de Frankia parmi les actinomycetes 8
Figure 3. Apparence de Frankia ssp. ACN14a en culture 10
Figure 4. Distribution globale de toutes les especes actinorhiziennes combinees 14
Figure 5. Distribution mondiale des plantes actinorhiziennes de la famille des Betulaceae.... 15
Figure 6. Infection d'une racine par Frankia, mecanisme intracellulaire 17
Figure 7. Exemples typiques de nodules d'Alnus viridis ssp. crispa 18
INTRODUCTION
1. Problematique etudiee : les sites perturb£s par l'industrie petroliere albertaine
Sous la foret boreale albertaine, dans l'Ouest canadien, est enfouie la deuxieme plus grande reserve de petrole mondiale. Les 175 milliards de barils extractibles que represente cette ressource ne reposent toutefois pas sous forme liquide tel le petrole d'Arabie Saoudite (Moorhouse et al, 2010). Les gisements sont plutot constitues d'un melange de grains de quartz, d'argile, de bitume, d'eau et de mineraux-traces ; ce sont les sables bitumineux (Rivard
et al., 2010). Ces sables bitumineux, tres denses et visqueux, sont principalement repartis en
trois gisements (Athabasca, Cold Lake et Peace River), et couvrent une superficie totale d'environ 140 000 km2, c'est-a-dire 20 % du territoire albertain (Toman et al, 2010).
Un projet pilote d'extraction du bitume en Alberta a ete mis en place des 1944, et l'exploitation commerciale de la ressource a debute en 1967. II faudra neanmoins attendre 1'implantation de nouvelles politiques gouvernementales en 1997, ainsi que la hausse des prix du petrole, pour que l'exploitation prenne r^ellement son envoi (Woynillowicz et al, 2005). La raison est simple : le bitume albertain est couteux et complexe a extraire (Engelhardt et Todirescu, 2005). De plus, son exploitation implique de lourdes consequences environnementales.
1.1 L'exploitation des sables bitumineux
Selon la profondeur des gisements, deux techniques sont utilisees pour recuperer le bitume des sables bitumineux. Les gisements localises a plus de 100 m de la surface sont exploites in situ
(sur place). Cette technique implique de creuser de multiples puits dans le depot, puis de chauffer les sables bitumineux a l'aide de vapeur d'eau afin de diminuer leur viscosite, pour ensuite les pomper vers la surface. Les sables bitumineux sont ensuite achemines par pipeline vers l'usine d'extraction (Moorhouse et al., 2010). Les gisements localises a moins de 100 m de la surface sont exploites par extraction miniere (Woynillowicz et al., 2005). La foret bordale est tout d'abord couple, puis le sol de recouvrement est retire, creant ainsi une mine a ciel ouvert. Les sables bitumineux sont excaves, concasses et melanges a de l'eau pour ensuite etre transports par pipeline a l'usine d'extraction (Toman et al., 2010).
La prochaine etape consiste a separer le bitume des autres composantes des sables bitumineux par le biais du procede d'extraction a la soude chaude de Clark (Clark, 1932). Brievement, les sables bitumineux sont chauffes a approximativement 85 °C, additionnes d'hydroxyde de sodium et malaxes, ce qui detache le bitume des particules solides et lui permet de flotter a la surface, ou il est recueilli par ruissellement. Ce procede permet d'obtenir du bitume brut qui doit etre fluidifie, c'est-a-dire additionne d'hydrocarbures plus legers, avant d'etre raffine (Alboudwarej et al., 2006).
Le processus de raffinage est constitue de trois etapes principales: la conversion thermique, la distillation et l'hydrotraitement. Le raffinage debute toujours par une premiere distillation permettant de retirer le naphta du bitume. Par la suite, le bitume est achemind a 1'unite de cokefaction, ou il est chauffe a 500 °C afin de fractionner les hydrocarbures lourds en molecules plus courtes, et done plus volatiles. II en r6sulte une fraction solide, le coke vert, constitu^ du carbone residuel, et une fraction gazeuse. Cette fraction sera gen£ralement distill6e et hydrotrait^e, e'est a dire chauffee sous pression en presence d'hydrogene. L'hydrotraitement stabilise les hydrocarbures en les saturant en hydrogene, en plus de retirer les impuretes telles que le soufre et les metaux lourds.
A
la fin du processus de raffinage, les principaux produits generes sont l'huile brute synthetique et le coke vert. L'huile est vendue sur le marche p^trolier, tandis que le coke vert, encore combustible, est converti en cokecalcine lorsqu'il est utilise pour chauffer les fournaises de la raffinerie (OSDC, 2008; Engelhardt et Todirescu, 2005).
1.2 Les sites perturb£s et les efforts de rehabilitation
L'expioitation petroltere en Alberta des perturbations environnementales majeures sur les sites d'extraction: l'excavation miniere scarifie le paysage, tandis que les nombreuses routes et pipelines qui sillonnent la foret boreale fragmentent les ecosystemes (Bordetsky, 2007). Le gouvernement albertain a cependant pris des mesures afin de proteger les terres. En effet, selon la section 137(1) du « Environmental Protection and Enhancement Act » (Government of Alberta, 2010), une fois l'expioitation petroliere terminee sur les sites, ceux-ci doivent etre restitu^s dans leur etat de productivity initiale. Cette exigence, apparemment aisee a satisfaire, est complexifiee par la presence de residus d' exploitation.
Les principaux dechets generes par l'expioitation des sables bitumineux sont les sables residuels (TS). En effet, 1'extraction a la soude chaude produit quantity de boue liquide composee d'eau alcaline, de sable et d'argile (Holroyd et Simieritsch, 2009). Cette boue est entreposee dans des bassins de decantation afin de permettre aux particules solides de sedimenter; l'eau est par la suite retiree pour etre reutilisee. Eventuellement, les bassins de decantation, ayant atteint le maximum de leur capacite, sont recouverts de sol de recouvrement et prets a etre revegetalis^s (Macyk et Drozdowski, 2008).
Le TS est un r^sidu sablonneux tres fin (Figure 1), alcalin, pauvre en nutriments, riche en sels et en acides naphteniques, tous des elements del^teres a la croissance des plantes. Les sols alcalins et ceux riches en sel sont connus pour leur induction du stress chez la plante (Gao et
2005). Specifiquement, les deux principaux sels presents dans le TS, le chlorure de sodium et le sulfate de sodium, entralnent une diminution de la croissance et une baisse de la quantite d'azote tissulaire (Anthraper et DuBois, 2003; Aydi et al., 2008; Santos et al, 2002; Redfield et Zwiazek, 2002); le chlorure de sodium est le plus dommageable des deux. Finalement, le TS contient des acides naphteniques, constituants naturels du petrole qui sont bien reconnus pour leur phytotoxicite (Clemente and Fedorak, 2005). lis entrainent des perturbations physiologiques ainsi qu'une diminution de la croissance des plantes (Armstrong et al., 2008).
Figure 1. Principaux residus g^neres par 1'industrie p&roliere albertaine.
A
gauche: TS, a droite: coke.Un second residu majeur g£nere par l'exploitation des sables bitumineux est le coke calcine, c'est-a-dire la matiere solide incombustible resultant de l'incineration du coke vert. Le coke est compost de billes de couleur noire et de granulom&rie variable (Figure 1). Sur le terrain, il est parfois accumule dans des fosses, puis couvert de sol de recouvrement (observations terrain). Un rapport exhaustif de The American Petroleum Institue Petroleum HPV Testing Group (2007) semble indiquer que le coke n'est pas un residu toxique. Neanmoins, selon ce meme report, le coke contient des quantites importantes de soufre, nickel et vanadium, ainsi
que des hydrocarbures aromatiques polycycliques (HAP), connus pour etre phytotoxiques (Alkio et al, 2005). Jusqu'a present, peu ou pas de travaux ont ete realises au sujet de la revegetalisation du coke.
De nombreuses strategies de revegetalisation ont ete mises sur pied par le gouvernement albertain et les compagnies petrolieres; un rapport exhaustif a ce sujet a ete public par Macyk et Drozdowski en 2008. De plus, plusieurs Etudes indiquent que la phytoremediation des residus contamines par les hydrocarbures est possible (Godwin et Thorpe, 2000; Gerhardt et
al, 2009). Malgre tout, un seul certificat a ete decern^ jusqu'a present par le gouvernement,
pour attester que Gateway Hill, d'une superficie de 1 km2, est retournee a son etat initial
(Lemphers et al, 2010). A l'heure actuelle, 686 km2 de foret boreale sont perturbes (Lemphers et al, 2010), et la superficie des bassins de decantation avoisine les 130 km2 (Woynillowicz et al, 2004; Holroyd et Simieritsch, 2009). Aucune des options de rehabilitation des terres mises
en place ne peut permettre de traiter le volume de residus qui seront generes d'ici la fin de Sexploitation des sables bitumineux. Des methodes de revegetalisation viables au niveau technique, economique et environnemental doivent etre mises au point (Quagraine et al, 2005).
2. Les organismes Studies
Plusieurs indices semblent suggerer que l'aulne actinorhize pourrait permettre la revegetalisation efficace des sites perturbes par l'industrie petroliere albertaine. Ce projet de recherche en microbiologic environnementale tente d'evaluer la capacite des aulnes symbiotiques a coloniser les residus de l'industrie petroliere albertaine. II s'efforce egalement d'estimer l'impact qu'auraient ces residus sur la symbiose entre deux especes d'aulnes indigenes du Canada, et une souche de Frankia isoiee au Canada egalement.
2.1. Frankia spp.
L'existence de symbioses racinaires actinorhiziennes est connue depuis presque 200 ans ; des 1829, les scientifiques ont soulev6 la possibility de la presence de microorganismes vivant dans les protuberances localisees sur les racines de diverses plantes (Schwenke et Caru, 2001). La description de ces microorganismes a ete efFectuee en 1866 dans un memoire presente par Woronin (Normand 2006). Vingt ans plus tard, un nom generique a ete propose par Brunchorst afin d'identifier les bacteries presentes dans les nodules des aulnes et des Elaeagnaceae: il s'agit de Frankia, en l'honneur de A.B. Frank, le microbiologiste ayant popularise l'usage du terme "symbiose" (Chaia et al., 2010; Lechevalier et Lechevalier, 1990). Malgre les efforts deployes, a cette epoque, l'etude de ce groupe de microorganismes s'averait plutot fastidieuse car limitee a l'etat symbiotique de la bacterie (Chaia et al., 2010). II aura fallu attendre pres de 100 ans pour obtenir une premiere isolation reproductible en culture pure a partir de nodules de Comptonia peregrina, grace au groupe de John Torrey (Callaham
et al., 1978). L'annee 1978 marque done l'eclosion de la recherche au sujet de Frankia, le
nombre d'isolats cultiv^s se multipliant alors de fa9on exponentielle (Chaia et al., 2010, Callaham et al., 1978).
Le genre Frankia a ete classifie parmi les actinomycetes par Krebber en 1932. II est aujourd'hui £galement associe au sous-ordre des Frankineae, tout comme son plus proche parent phylogenique, Acidothermus, une bacterie acidophile degradant la cellulose (Figure 2) (Benson et Dawson 2007). Bien que la description du genre Frankia et la creation de la famille Frankiaceae aient ete acceptees a 1'unanimity tel que decrit par Lechevalier et Lechevalier (1989), la definition d'especes de Frankia s'est av£ree plus complexe et controversee. En effet, les techniques classiques sont difficilement applicables a Frankia, entre autres en raison de son temps de generation eleve (15 a 48 h, voire davantage) (Benson et Silvester, 1993). Lalonde et son equipe avaient tout de meme propose en 1988, dans l'ouvrage "Nitrogen fixation : hundred years after" la creation de deux especes, F. elaeagni et F. alni,
cette derniere etant subdivide en deux sous-especes, pommerii et vandijkii (Benson et Silvester, 1993). Cette suggestion etait basee sur des etudes biochimiques et physiologiques effectuees sur des souches de Frankia isol6es de 4 families de plantes actinorhiziennes differentes. Peu apres, Fernandez et al. (1989) a compart 43 souches par le biais de l'homologie de sequences d'ADN et a ainsi identifie 9 especes genomiques distinctes. Plusieurs autres etudes a ce sujet se sont succedees au fil des annees, sans qu'aucune ne soit exempte de contradictions lorsque comparee h la litterature. II a done ete etabli que jusqu'a ce qu'on comprenne mieux les caracteristiques definissant une esp£ce chez Frankia, aucun nom d'espece ne serait utilise. Un code pour designer les souches a plut6t ete mis en place par Lechevalier (1983). Selon celui-ci, 1'etiquette pour chaque souche est composee de 3 lettres designant le laboratoire d'origine et de 2 chiffres correspondant au genre de la plante-hote, suivis d'un code unique de 6 a 8 chiffres.
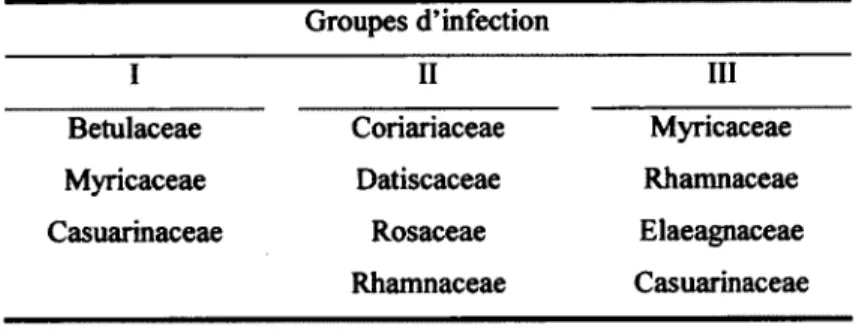
Grace aux Etudes phylogenetiques basees sur le gene codant pour l'ARN ribosomal 16S, 4 groupes de souches ont tout de meme ete definis. Les trois premiers groupes correspondent approximativement a la specificite d'infection des hotes (Tableau 1), tandis que le 4e groupe
regroupe des actinomycetes de type Frankia qui, selon les connaissances actuelles, n'induisent pas la formation de nodules, et ne fixent pas l'azote atmospherique (Wall, 2000). Ces techniques avaient cependant revele une faible diversity phylogenetique chez le genre Frankia (Lechevalier, 1994). Or, de nouveaux outils de biologie mol6culaire semblent reveler le contraire et permettent de mieux ^valuer la richesse propre a ce genre (Bern6che-D' Amours et
al., 2011). Pour completer le tout, malgr£ sa croissance lente et agglomeree en faisant un
microorganisme difficile k etudier, de nouvelles techniques en microplaques permettent maintenant d'effectuer une caracterisation biochimique exhaustive des diff&rentes souches (Belanger et al. 201 la).
JflNUdMlDWAMN J Streptomycin### Mtmmonotporintm 002 jMkracocctcM* JSfripftMporanptoM* jActfMpotaporftMM JAethomyetm— JG/yeomyctm* • Pftptoufcicdrfioiii Actll9O0M^P^pOfl0CGM — MtmyMlMMt G^rmvcNHH* ~™ fltfctotocfvrfsoM# ^Ic&taRcreMiOM# »! Auteotecttntftt 100
Figure 2. Arbre phylogenetique illustrant la position de Frankia spp. parmi les actinomycetes. Tire de Zhi et al 2009, avec permission.
Tableau 1. Liste des families vegetales actinorhizees par les souches de Frankia selon le groupe d'infection auquel elles appartiennent
Groupes d'infection
I II III
Betulaceae Coriariaceae Myricaceae Myricaceae Datiscaceae Rhamnaceae Casuarinaceae Rosaceae Elaeagnaceae
Rhamnaceae Casuarinaceae
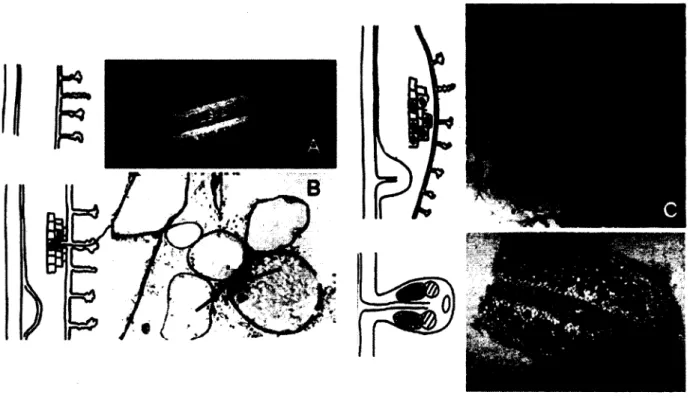
Les bacteries du genre Frankia sont des actinomycetes a Gram positif, fixateurs d'azote, ayant la capacite d'induire la nodulation chez plusieurs especes vegetales (Lechevalier et Lechevalier, 1990). Elles sont reconnues pour avoir un genome tres riche en nucleotides G+C, une paroi cellulaire de type III et un patron de phospholipides de type phosphatidylinositol (Lechevalier et Lechevalier, 1990; Benson et Silvester, 1993; Schwencke et Caru, 2001). Le temps de generation de Frankia spp. est long: de 15 a 48 h, voire davantage (Benson et Silvester, 1993). La propagation de Frankia spp. en laboratoire s'effectue generalement par le biais de cultures liquides statiques maintenues entre 25 et 30 °C (Belanger et al. 2011a; Normand 2006). Frankia prend alors l'aspect d'un mycelium floconneux generalement blanc (Figure 3 A) (Scwencke et Caru, 2001). La production de pigments de couleurs tres variees est souvent observee chez les cultures plus agees (Lechevalier et Lechevalier, 1990). Recemment, un milieu solide permettant la croissance de colonies de Frankia visibles a l'oeil nu en moins de 3 jours a ete mis au point. II favorise £galement la sporulation du microorganisme (Bassi et Benson, 2007).
En culture, on peut retrouver chez Frankia spp. trois structures typiques aux actinomycetes (Schwencke et Caru, 2001). Les hyphes vegetatifs, filamenteux, septes et faiblement branches, permettent la croissance de la bacterie (Figure 3B). Lorsque soumise a des conditions
limitantes en azote, Frankia produit egalement des structures spheriques, les vesicules, qui permettent la fixation d'azote atmospherique (Figure 3B). A la fin de la courbe de croissance, ou lorsque les conditions sont defavorables, une troisieme conformation apparalt chez certaines souches: les sporanges (Figure 3C). Ceux-ci contiennent les sporangiospores, qui jouent un role dans la dissemination de la bacterie. II existe finalement une quatrieme structure, relativement meconnue et nomm^e hyphes de reproduction toruleux (Figure 3D). Tout comme les spores, ces hyphes sembleraient impliques dans la propagation de Frankia (Diem et Dommergues, 1985).
Figure 3. Apparence de Frankia ssp. ACN14a en culture. A. Culture liquide en BAPS. B. Hyphes et vesicules (V) observes en microscopie interferentielle, grossissement 60 X. C. Sporange et hyphes colons au Live/Dead, observes en microscopie a epifluorescence, grossissement 100 X. D. Hyphe de reproduction toruleux colors au Live/Dead, observe en microscopie a Epifluorescence, grossissement 100 X.
La distribution mondiale du genre Frankia est vaste : on le retrouve sur tous les continents, exception faite de l'Antarctique (Benson et Dawson, 2007; Chaia et al., 2010). En plus de vivre en symbiose avec les plantes actinorhiziennes, plusieurs souches de Frankia peuvent survivre et proliferer en mode saprophyte dans le sol, meme lorsque qu'aucune plante-hote n'est presente sur les sites (Huss-Danell et Frej, 1986; Smolander et Sundman, 1987; Markham et Chanway, 1996; Maunuksela et al, 1999; Chaia et al., 2010). II semblerait done que si la degradation des tissus nodulaires contribue au maintien de la population de Frankia dans les sols (Benson et Silvester, 1993), divers moyens permettent par la suite sa dispersion. II a ete demontre que l'eau, les invertebres, les oiseaux et memes les mammiferes peuvent efficacement contribuer au transport de Frankia (Arveby et Huss-Danell, 1988; Reddell and Spain 1991; Paschke and Dawson, 1993; Chaia et al., 2010). La propriete des sols a induire la nodulation en absence de plantes-hotes pourrait aussi etre attribuee a la faculte de Frankia de persister dans ces sols pendant de nombreuses annees. Alnus incana a ete nodule par des souches vieilles de 30 ans lorsque plante dans des sediments tires d'un lac (Huss-Danell et al., 1997), tandis que Discaria trinervis l'a ete par des souches subsistant dans des sediments vieux de 50 ans (Chaia et al., 2005).
Une des caracteristiques les plus remarquables de Frankia spp. est sa capacite a fixer le diazote, non seulement en symbiose, mais egalement en mode saprophyte (Normand, 2006). Selon les connaissances actuelles, seuls les procaryotes sont capables de fixer 1' azote atmospherique, et seulement 6 des 53 phyla du domaine des bacteries incluent des diazotrophes (Martinez-Romero, 2006). La conversion de l'azote atmospherique en ammoniac est catalysee par la nitrogenase, un complexe enzymatique tres conserve parmi les microorganismes fixateurs d'azote (Franche, 2009); les genes nif codent pour les diverses composantes du complexe (Huss-Danell, 1997). La nitrogenase classique est composee de deux metalloprot&nes, la premiere complexee au fer, la seconde complexee au fer et au molybdene (Martinez-Romero, 2006). Ces proteines sont associees k un cofacteur fer-molybdene, qui constitue le site actif du complexe enzymatique (Kim et Rees, 1992). En plus de la nitrogenase classique, ou nitrogenase au molybdene, deux autres types de nitrogenases
sont presents chez la plupart des microorganismes: la nitrogenase au vanadium, et la nitrogenase exclusivement au fer. Ces nitrogenases alternatives permettent de s' adapter aux conditions environnementales limitantes en molybdene (Martinez-Romero, 2006).
La presence d'oxygene constitue un autre facteur environnemental pouvant affecter la fixation d'azote, car la nitrogenase est denaturee par l'oxyg&ne. Cette sensibilite represente un probleme pour les microorganismes aerobies, puisque leur metabolisme energetique repose majoritairement sur l'oxygene et que la fixation d'azote requiert beaucoup d'energie. II existe une variete de strategies pour resoudre le dilemme de l'oxygene (Silvester et al., 2008). Pour sa part, Frankia est pourvue d'une structure protegeant sa nitrogenase de l'oxygene: la vesicule (Figure 3B). Les vesicules sont constitutes de plusieurs couches de lipides (jusqu'a 95) composees principalement d'hopanoides, qui abritent un groupe de cellules exprimant la nitrogenase (Harriott et al, 1991; Silvester et al, 2008). La couche de lipides agit comme une barriere empechant la diffusion de l'oxygene, et Frankia peut en reguler l'epaisseur et la composition selon la concentration en oxygene du milieu (Berry et al., 1993; Wall, 2000). Par ailleurs, en guise de protection supplementaire, on peut retrouver a proximite de la nitrogenase une concentration tres elevee d'hemoglobine (Silvester et al., 2008). Finalement, a l'instar de plusieurs diazotrophes, Frankia peut egalement beneficier de la protection d'une plante-hote (Benson et Silvester, 1993; Franche, 2009). Les plantes ayant la capacite d'entrer en symbiose avec Frankia sont nommees plantes actinorhiziennes.
2.2. Les plantes actinorhiziennes
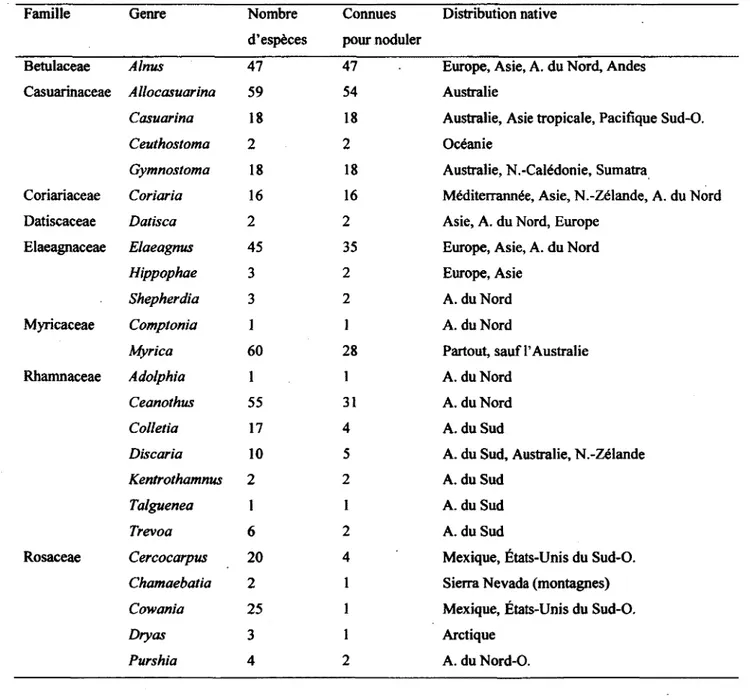
Contrairement a la symbiose rhizobienne, qui implique presque exclusivement une seule famille, Leguminosae, il existe pres de 250 espdces connues de plantes, r^parties en 25 genres et 8 families, capables d'entrer en symbiose avec la bacterie saprophyte Frankia (Tableau 2) (Huss-Danell, 1997; Gualtieri et Bisseling, 2000; Schwencke et Caru, 2001; Dawson, 2008).
De ce nombre, toutes sont des arbres et arbustes, exception faite de Datisca, qui possede des racines ligneuses, mais des parties aeriennes herbacees (Benson et Dawson, 2007).
Tableau 2. Les plantes actinorhiziennes et leur distribution, traduit de Dawson 2008
Famille Genre Nombre d'esp^ces
Connues pour noduler
Distribution native
Betulaceae Alnus 47 47 Europe, Asie, A. du Nord, Andes Casuarinaceae Allocasuarina 59 54 Australie
Casuarina 18 18 Australie, Asie tropicale, Pacifique Sud-O.
Ceuthostoma 2 2 Oc^anie
Gymnostoma 18 18 Australie, N.-Cal6donie, Sumatra
Coriariaceae Coriaria 16 16 M6diterrann6e, Asie, N.-Z61ande, A. du Nord Datiscaceae Datisca 2 2 Asie, A. du Nord, Europe
Elaeagnaceae Elaeagnus 45 35 Europe, Asie, A. du Nord
Hippophae 3 2 Europe, Asie
Shepherdia 3 2 A. du Nord Myricaceae Comptonia 1 1 A. du Nord
Myrica 60 28 Partout, sauf 1'Australie Rhamnaceae Adolphia 1 1 A. du Nord
Ceanothus 55 31 A. du Nord
Colletia 17 4 A. du Sud
Discaria 10 5 A. du Sud, Australie, N.-Z61ande
Kentrothamnus 2 2 A. du Sud
Talguenea 1 1 A. du Sud
Trevoa 6 2 A. du Sud
Rosaceae Cercocarpus 20 4 Mexique, Etats-Unis du Sud-O.
Chamaebatia 2 1 Sierra Nevada (montagnes)
Cowania 25 1 Mexique, Etats-Unis du Sud-O.
Dryas 3 1 Arctique
La niche ecologique des diverses especes de plantes actinorhiziennes est constitute de lieux ou le sol est pauvre en azote. L'azote disponible dans les sols est un facteur tres limitant pour la croissance des plantes en general, mais grace a leur symbionte, les plantes actinorhiziennes peuvent coloniser de tels habitats (Martinez-Romero, 2006; Franche et al., 2009). Ces habitats ont parfois subi des perturbations : c'est le cas des depots volcaniques, des sols mis a nu apres le retrait de glaciers, des dunes de sable, des sols erodes a la suite d'un glissement de terrain. L'azote est egalement limitant dans des ecosystemes non perturbes: les prairies, les marais et les zones chaudes et arides en sont de bons exemples (Dawson, 2008; Schwencke et Caru, 2001; Wall, 2000).
Figure 4. Distribution globale de toutes les especes actinorhiziennes combinees. Tire de Benson et Dawson 2007, avec permission.
Aussi diversifies soient-ils, on retrouve des environnements limites en azote partout k travers le monde. Ainsi, il n'est pas etonnant de retrouver des especes actinorhiziennes sur tous les continents, exception faite de 1' Antarctique (Figure 4). La plupart des plantes actinorhiziennes croissent dans les Ecosystemes boreaux et temptres des deux hemispheres. La famille des
Casuarinaceae fait toutefois office d'exception, puisque certaines Casuarina et toutes les
Gymnostoma spp. sont endemiques de forets tropicales humides (Benson et Dawson, 2007).
Avec un taux de fixation d'azote annuel estim£ a 240-350 kg/ ha/an, les plantes actinorhiziennes font souvent office de plantes pionnieres lors de la colonisation de nouveaux habitats (Wall, 2000; Schwencke et Caru, 2001). Cette qualite de plantes pionnieres est exploits k diverses fins industrielles et environnementales : plantes omementales, production de bois, agroforesterie, prevention de la desertification, restauration de berges et de sols perturbes sont tous des usages auxquels les plantes actinorhiziennes sont destinees (Huss-Danell, 1997; Wall, 2000; Roy etal, 2007).
Figure 5. Distribution mondiale des plantes actinorhiziennes de la famille des Betulaceae. Tire de Benson et Dawson 2007, avec permission.
Dans le cadre de la rehabilitation et de la rev^getalisation des terres, un genre actinorhizien revet un interet particulier: Alnus. Bien qu'il soit le seul genre de la famille des Betulaceae h etablir la symbiose avec Frankia (Bousquet et al., 1989), toutes les especes d'aulne connues
sont actinorhizees (Huss-Danell, 1997). La grande majority des aulnes sont egalement colonises par des mycorhizes, qui facilitent l'absorption de l'eau et du phosphore (Roy et al, 2007). La vaste repartition mondiale du genre Alnus (Figure 5) traduit son adaptability a une grande variete d'ecosystemes: l'aulne colonise tout autant les bordures des marais et des rivieres que les forets, les toundras et les milieux alpins (Benson et Dawson, 2007; Roy et al, 2007). Parmi les plantes actinorhiziennes, c'est probablement celle qui resiste le mieux au vieillissement et qui conserve un taux de fixation d'azote significatif malgre l'age (Schwencke et Cani, 2001). L'aulne poss&Ie neanmoins une faible tolerance a I'ombre, en raison de la grande quantite d'energie necessaire pour soutenir la symbiose actinorhizienne (Roy et al, 2007).
2.3 La symbiose entre l'aulne et Frankia
2.3.1 Le processus de nodulation
Contrairement au processus de nodulation chez Rhizobium, qui est tres bien caracterise, plusieurs aspects au sujet du processus chez Frankia sont encore a explorer. Puisqu'il existe plusieurs similitudes entre les deux mecanismes, les connaissances au sujet de Rhizobium offrent cependant des pistes de recherche interessantes, que ce soit au sujet des messagers chimiques impliques qu'au niveau de la fonction des diverses structures (Pawlowski et Sprent, 2008).
Peu d'informations sont connues au sujet de la colonisation des racines par Frankia. Valverde et Wall ont observe que quelques jours apres avoir inocule des plants de Discaria trinervis avec Frankia, la bacterie s'etait accumulee a certains points pres des racines de la plante; ces points sont plus tard devenus les sites de nodulation (Valverde et Wall, 2003). Une
observation similaire a ete effectuee par Berry en inoculant des aulnes (Wall et Berry, 2008).
Frankia est une bacterie non-mobile, aussi, le mecanisme lui permettant de s'accumuler
ponctuellement pres des racines est difficilement explicable, mais vraisemblablement, des exsudats racinaires seraient impliques (Wall et Berry, 2008). Une fois la bacterie h proximite des racines, elle peut infecter la plante selon deux mecanismes: intercellulaire et intracellulaire. Puisqu'une meme souche de Frankia peut utiliser les deux modes selon la famille v^getale infectee, il est probable que l'organisme controlant la voie d'infection soit la plante-hote. Les plantes infectees via le mecanisme intracellulaire sont dites primitives:
Alnus, Comptonia, Casuarina, et Myrica font partie de ce groupe (Wall, 2000; Schwencke et
Caru, 2001).
Figure 6. Infection d'une racine par Frankia, mecanisme intracellulaire. A. Deformation du poil racinaire. B. Infection intracellulaire, division corticale, primordium nodulaire induit dans le pericyle. C. Prenodule. D. Nodule mature. Tire de Wall et Berry 2008, avec permission.
L'infection par le biais du mecanisme intracellulaire (Figure 6) debute lorsque Frankia entre en contact avec le poil racinaire de l'hote, qui se d^forme, produit des embranchements et secrete davantage de mucilage (Schwencke et Caru, 2001). Les hyphes bacteriens traversent alors la paroi cellulaire du poil a un locus ou celui-ci est replie et penetrant les tissus du poil jusqu'a atteindre les cellules corticales (Gualtieri et Bisseling, 2000). Durant le processus, la membrane des cellules de la plante s'invagine et une nouvelle paroi est synthetisee au fil de la progression de Frankia, formant un tube d'infection. Tandis que Frankia progresse vers Pint6rieur du cortex racinaire, les cellules du cortex rearrangent leur cytosquelette et se divisent (Wall, 2000). La bacterie infecte alors les cellules corticales en division et produit des vesicules; l'ensemble forme le prenodule, qui se traduit par une petite protuberance a la surface racinaire (Franche et al., 2009). Chez l'aulne, le prenodule apparait generalement une semaine apres l'infection (Huss-Danell, 1997). Finalement, la division cellulaire est induite dans le pericycle, et le primordium nodulaire croit vers la surface racinaire, incorporant une partie de l'inoculum du pr&iodule au passage (Wall, 2000; Chaia et al, 2010; Franche et al., 2009).
V
Figure 7. Exemples typiques de nodules d'Alnus viridis ssp. crispa.
II en resulte un lobe, avec un faisceau vasculaire central entoure de l'endoderme, des cellules corticales infectees par Frankia, et du periderme. Chez l'aulne, les premiers lobes de nodules apparaissent entre 2 et 3 semaines apr£s l'inoculation (Huss-Danell, 1997). De nouveaux lobes continuent d'emerger, pour eventuellement former un nodule coralliforme (Figure 7) (Franche, 2009). Puisque la symbiose est couteuse en energie pour l'aulne, celui-ci limite la masse de nodules produits selon ses besoins en azote grace a un mecanisme de retrocontrole nomme autoregulation (Wall et Huss-Danell, 1997).
2.3.2 Les facteurs influen^ant la nodulation
L'etendue de la nodulation observee depend du genotype de la plante, du genotype de Frankia et des facteurs environnementaux affectant les deux organismes (Chaia et al., 2010). Myrold et Huss-Danell (1994) ont demontre que lorsque des plantules de trois especes d'Alnus etaient inocules avec un sol suedois provenant de la distribution naturelle d'A. incana, l'&endue de la nodulation 6tait toujours plus importante chez A. rubra, suivi de A. incana, puis de A.
glutinosa, peu importe la methode utilisee pour preparer l'inoculum ou le type de solution
d'arrosage. Cette constatation indique que le mecanisme de specificite symbionte-hote base sur un principe signal-r^cepteur decrit par Wall (2000) existe probablement h l'echelle souche-espece. II semblerait egalement que les autres microorganismes du sol aient une influence, souvent positive, sur la symbiose entre l'aulne et Frankia. Par exemple, selon une etude par Rojas et al. (2002) impliquant Alnus rubra, la symbiose avec le champignon ectomycorhizien
Alpova diplophloeus augmente le taux de nodulation de 30 %. Plusieurs autres Etudes ont tire
des conclusions similaires, sans surprise, puisque la symbiose mycorhizienne facilite l'apport en eau et en phosphore, protege la plante de certains stress et produit des phytohormones (Roy
et al., 2007). II a egalement ete demontr^ que certaines bacteries du sol, Pseudomonas, Streptomyces, Actinoplanes, Micromonospora, et Bacillus, favorisent la nodulation chez les
plantes actinorhiziennes, entre autres chez Alnus rubra et Alnus glutinosa (Zimpfer, 2003; Knowlton et Dawson, 1983; Knowlton et al., 1980; Solans, 2007). La presence de plantes
non-hotes a proximite des plantes actinorhiziennes accroit aussi l'etendue de la nodulation (Chaia et al., 2010).
Les facteurs abiotiques ont egalement un effet sur l'etendue de la symbiose actinorhizienne. Par exemple, si les conditions du sol sont acides, il existe une correlation positive entre le nombre de nodules arbores par Alnus et le pH (Smolander et Sundman, 1987; Elo et al, 2000). Le pourcentage d'humidite du sol semble egalement un facteur important. En effet, plusieurs etudes demontrent que l'efficacite de la nodulation augmente en fonction du pourcentage d'humidite, mais que les sols constamment inondes ne demontrent aucun signe d'infectivite (Dawson, 2008). II a aussi ete observe que certaines souches de Frankia originates de milieux humides peuvent noduler A. glutinosa, mais ne fixent pas 1'azote atmospherique dans ces conditions faibles en oxygene (Van Dijk et Sluimer-Stolk, 1990). Ces constatations ne sont pas etonnantes, compte tenu que Frankia est une bacterie microaerophile, done qu'elle necessite un apport en oxygene, et que les sols gorges d'eau en sont depourvus. De nombreuses etudes demontrent d'ailleurs que les sols riches en O2 ameliorent le taux de nodulation (Dawson, 2008).
En contrepartie, il est bien connu que les sols est riches en azote, inhibent la nodulation (Kohls et Baker, 1989; Martin et al, 2003; Gentili et Huss-Danell, 2003; Gentili et al., 2006). II semblerait que cette inhibition se deroule avant meme la deformation des poils racinaires (Ceremonie et al, 1999). Au contraire, la limitation en phosphore est delet&re k la production de nodules chez plusieurs especes, dont A. glutinosa et A. incana (Gentili et al, 2006; Chaia et
al., 2010; Valdes, 2008). Fait intgressant, une concentration dlevee en phosphore permet de
compenser pour la presence d'azote et retablit la production de nodules, mais ne restitue pas la faculty de fixer l'azote (Gentili et Huss-Danell, 2003). Chez les plantes actinorhiziennes en general, la reponse a un taux de salinite important est variable. De fait, les plantes-hotes possedant naturellement des niches ecologiques riches en sels sont tres tol6rantes au stress hydrique entrain^ par une salinite elevee, de meme que le sont lews symbiontes (Chaia, 2010;
Valdes, 2008; Dawson, 2008). II est a noter que la temperature, la luminosite et le cycle des saisons, la profondeur du sol et l'altitude du terrain, ainsi que l'etape de succession vegetale sont egalement des facteurs qui modifient la capacite de nodulation chez les plantes actinorhiziennes (Chaia et al., 2010, Valdes, 2008; Dawson, 2008).
Les contaminants anthropogeniques constituent une source de stress non negligeable pour les organismes du sol. Malheureusement, aucune &ude quantitative au sujet de 1'efFet sur la nodulation n'a ete realisee au sujet des herbicides, des dechets d'usine ou de decharges, ou encore des hydrocarbures. Cependant, les effets des metaux lourds sont mieux caracteris6s. II a ete demontre que parmi les 14 m&aux testes, sur une certaine gamme de concentrations, le nickel avait une influence positive sur la nodulation, tandis que 1'aluminium, le manganese et le cadmium reduisaient le nombre de nodules (Belanger et al., 2011b). La litterature, disponible pour certains de ces metaux, valide les resultats (Wheeler et al, 2001; Dawson, 2008; Kochian et al., 2004).
2.3 J Les avantages procures par la symbiose actinorhizienne
II a souvent ete enonce que la symbiose est Fun des principaux moteurs de Involution. Elle permet aux organismes de s' adapter a des habitats hostiles et de les coloniser, tandis qu'ils n'auraient pu prosperer, ou meme survivre seuls dans de telles conditions (Fortin et al., 2008). Dans le cas de Fassociation entre Frankia et l'aulne, il serait difficile de contester que la symbiose apporte un avantage a chacun des organismes.
Le premier avantage que Frankia retire de la symbiose est une protection physique. Puisque la bacterie est a l'abri dans les cellules corticales du nodule, elle est davantage epargnee par les fluctuations des conditions environnementales, par exemple une periode de secheresse ou un
exces d'humidite. De plus, dans le nodule, Frankia est protegee des microorganismes competiteurs et, bien qu'elle y soit assez resistante, des antibiotiques qu'ils peuvent secreter (Normand, 2006; S. Roy, resultats non publics). L'hote offre egalement une barriere supplementaire face a l'oxyg&ie, qui peut denaturer la nitrog6nase (Huss-Danell, 1997). L'approvisionnement en composes carbones et en energie pour soutenir la fixation d'azote constitue cependant la plus grande contribution de la plante envers Frankia (Valverde et Huss-Danell, 2008). La cooperation entre les deux organismes a ce sujet est necessaire, car la fixation d'azote est un processus couteux en 6nergie : pour chaque molecule de N2 transformee et deux molecules d'ammoniac produites, de 20 a 30 molecules d'ATP sont consommees (Burris, 1991).
En laboratoire, l'aulne peut croitre en absence de symbionte, a condition qu'une source d'azote, tel le KNO3, soit prodiguee. Toutefois, sur le terrain, tous les aulnes observes portent des nodules (Huss-Danell, 1997; Chaia et ah, 2010). II semblerait done que l'aulne soit largement dependant de la fixation d'azote pour croitre; d'ailleurs, en symbiose, Frankia comble de 70 a 100 % des besoins en azote de l'aulne (Myrold et Huss-Danell, 2003; Hurd, 2001). La fixation d'azote permet a l'aulne actinorhize de coloniser des habitats divers, dont plusieurs hostiles pour la plupart des autres vegetaux : toundra, masses de gravier, sols mis a nu apres le retrait d'un glacier, depots d'eruption volcanique (Roy et al., 2007; Dawson et al., 2008). Enfin, un autre benefice considerable de l'association entre Frankia et Alnus est que cette symbiose ameliore la resistance de l'aulne aux agents pathogenes (Wall, 2000).
La symbiose entre Frankia et Alnus n'est pas qu'avantageuse pour les deux partenaires, elle est Egalement d'interet pour leur habitat et pour les autres organismes qui y evoluent. En effet, sous plusieurs aspects, les aulnes ameiiorent la qualite des sols qu'ils colonisent (Perinet et al., 1985). Tout d'abord, ils ameiiorent la structure et la stabilite des sols grace a leur systeme racinaire developpe (Knowlton et Dawson, 1983). Ils participent egalement a l'apport en azote des ecosystemes. Selon le livre Nitrogen fixation in plants publie en 1986 par Dixon et
Wheeler, environ 25 % de la fixation d'azote planetaire proviendraient des plantes actinorhiziennes (Dawson, 2008), et la contribution des aulnes actinorhizes a cette statistique est non negligeable. Les plantes du genre Alnus sont singulieres en ce sens qu'elles n'emmagasinent pas l'azote foliaire dans leurs organes vivaces une fois rautomne venu (Cote
et al., 1989). Grace a cette caracteristique, environ 45 % de l'azote fixe dans le cadre de la
symbiose retournent a l'ecosysteme par le biais de la chute des feuilles (Huss-Danell, 1997). De plus, une fois decomposees, les feuilles enrichissent le sol en matiere organique (Huss-Danell et Lundmark, 1988). Cette abondante chute de feuilles riches en azote explique en grande partie pourquoi les aulnes font d'excellents vegetaux-compagnons qui permettent aux plantes qui les cotoient de croitre plus rapidement (Paschke et al., 1989). L'aulne, resistant a plusieurs champignons pathogenes, limiterait meme la propagation de ceux-ci dans les plantations de coniferes intercalees (Hibbs et Cromack, 1990; Schwencke et Caru, 2001).
3. Les aulnes : leur utilisation anterieure et actuelle
Le peuple Inca serait le premier a avoir tire profit des nombreuses qualites des aulnes actinorhizes. Des 1000 ans avant J.-C, ils ont utilise l'aulne des Andes pour reduire l'erosion des sols de leurs champs localises en montagne, puis en ont tire du bois de charpenterie, de chauffage, et de l'engrais vert. Encore aujourd'hui, ces techniques sont en usage chez les indigenes d'Amerique Centrale (Dawson, 2008). Cette application des aulnes ne constitue pas un cas isold, au contraire : un peu partout a travers le monde, l'aulne fait partie integrante de diverses pratiques agroforestieres. Par exemple, en Grande-Bretagne, l'aulne rouge est plante a densite moderee dans les paturages, dont les especes herbac6es 6puisent rapidement les reserves azotees des sols. II contribue 6galement au maintien de la structure du sol, et offire un couvert aux animaux en cas d'intemp&ies (Teklehaimanot et Mmolotsi, 2007). En Inde,
Alnus nepalensis est utilise pour rev^getaliser certaines fermes abandonnees, et forme souvent
des plantations mixtes avec de la cardamome et du the (Normand, 2006). Les cultures agricoles ne sont pas les seules a profiter de la presence de l'aulne: les forets suedoises
destinees a la production de bois, appauvries par une surexploitation et un climat peu clement, beneficient de son apport en azote (Huss-Danell et Lundmark, 1988; Myrold et Huss-Danell, 2003). En Amerique du Nord, le grand potentiel agroforestier des aulnes a frequemment ete mis a profit pour augmenter la productivity de plantations d'espdces vegetales d'interet 6conomique. Une variete de circonstances ou des aulnes ont ete intercales parmi des coniferes a ete repertoriee par Hibbs et Cromack dans leur ouvrage de 1990, Actinorhizal plants in
Pacific Northwest forest (Schwencke et Cani, 2001); I'exemple le plus frequemment observe
est celui du pin de Douglas soutenu par l'aulne rouge (Gordon et Dawson, 1979; Hart et al, 1997). Par ailleurs, l'aulne rouge, dont le bois est exploitable commercialement, est souvent utilise pour rentabiliser a court terme les plantations d'essences nobles, dont la croissance est lente, mais la valeur marchande elevee. Cette association est economiquement tr6s avantageuse, puisque selon certaines etudes, le noyer noir pourrait atteindre sa maturite deux fois plus rapidement lorsque intercale avec Alnus rubra, et ainsi etre recolte a peine 40 ans apres sa plantation (Gordon et Dawson, 1979; Paschke et al., 1989; Schwencke et Caru, 2001, Huss-Danell, 1997).
En plus d'etre utiles en agroforesterie, les aulnes sont egalement d'interet dans le cadre de la rehabilitation des terres a la suite de perturbations, generalement anthropomorphiques. En Alaska, Alnus viridis a conjointement ete utilise avec Populus balsamifera et Salix alaxensis pour revdgetaliser les residus d'une mine de quartz (Densmore, 2005). En Angleterre, Alnus
incana a ete plants dans 10 sols containing par divers residus industriels, majoritairement des
cocktails de metaux lourds et de HAP (Ridgway et al, 2004). Les aulnes ont colonise les substrats avec succes, et l'analyse des nodules a permis de demontrer que chaque sol supportait une communaute de Frankia differente. Une autre etude a revele que six especes d'aulne peuvent rehabiliter des mines de charbon desaffectees, et permettent d'ameliorer la croissance du meteze japonais sur ces sites (Moffat, 2000). Des r^sultats similaires ont ete obtenus en Italie avec l'aulne italien plante sur des sites parsemes de lignite (Lumini et al, 1994). Si, en Amerique du Nord, l'aulne rouge est l'esp6ce utilisee a profusion pour ameliorer la quality des sols, en Europe, l'espdce la plus exploitee est l'aulne noir. Alnus glutinosa peut
coloniser des sites contamines par 1'exploitation de schistes bitumineux et y promouvoir la proliferation de microorganismes degradateurs d'hydrocarbures (Vares et al, 2004). II peut egalement croitre dans des sols contamines par des metaux lourds (Whitbread-Abrutat, 1997; Rosselli et al, 2003), ou des residus d'acetylene et de chlorure de polyvinyle (Oliveira et al, 2005).
Dans le cadre de la problematique gener^e par l'exploitation des sables bitumineux en Alberta, l'EPEA (Government of Alberta, 2010) stipule que les terres doivent etre revegetalisees avec des especes indigenes a la province. Sept especes et sous-especes d'aulnes sont natives du Canada: l'aulne rugueux, l'aulne des montagnes, l'aulne rouge, l'aulne blanc, l'aulne crispe, l'aulne de Sitka et l'aulne de Siberie. Parmi celles-ci, l'aulne crispe (Alnus viridis (Chaix) DC. ssp. crispa) et l'aulne rugueux (Alnus incana (L.) Moench ssp. rugosa) constituent des choix interessants pour la revegetalisation en Alberta car ils croissent deja dans cette province et ont deja demontre des aptitudes a la rehabilitation des terres au Canada. Par exemple, les deux especes ont ete utilisees pour revegetaliser des champs quebecois abandonnes dans l'optique de produire de la biomasse (Pregent et Camire, 1985). Elles ont aussi prouve qu'elles pouvaient tolerer les contaminants en colonisant d'anciennes mines de cuivre en Ontario et au Manitoba; la presence de l'aulne y a am61iore la qualite des sols et a promu la croissance de l'epinette noire (Vogel et Gower, 1998; Markham, 2005). II semblerait qu'en plus de tolerer certains m£taux lourds, l'aulne rugueux soit prometteur pour revegetaliser les sites contamines aux hydrocarbures, puisqu'il tolere bien la presence de ceux-ci (Godwin et Thorpe, 2000). Des etudes ont Egalement demontre que l'aulne crispe peut etre produit et utilise efficacement dans le cadre de revegetalisation a grande echelle : Perinet et al. ont plante 7 millions de plantules
d'Alnus crispa inocules avec des cultures pures de Frankia spp. afin de stabiliser des digues a
la Baie James au Quebec (1985). Plus r^cemment, le complexe Bonnyville Forest Nursery en Alberta a d&nontre qu'il pouvait produire des aulnes crispes en grand nombre pour soutenir la revegetalisation des terres perturbees par l'exploitations des sables bitumineux (Quoreshi et
4. Hypotheses et objectifs du projet de recherche
Sexploitation des sables bitumineux en Alberta perturbe la foret bor^ale, en plus de produire une quantite importante de residus aux proprietes phytotoxiques. Jusqu'a present, aucune des options de rehabilitation des terres mises en place ne permet de traiter le volume des principaux residus generes par l'exploitation, le TS et le coke. Deux especes vegetales indigenes de l'Alberta ont deja, sous d'autres contextes, demontre des aptitudes a la colonisation d'habitats perturbes et contamines : il s'agit de l'aulne crispe et l'aulne rugueux en symbiose avec la bacterie du genre Frankia. Ce projet de recherche en microbiologic environnementale avait pour but d'evaluer la capacite des aulnes symbiotiques a coloniser directement les principaux residus de l'industrie petroliere albertaine. II s'est egalement efforce d'estimer l'impact qu'auraient ces residus sur la symbiose de l'aulne crispe et l'aulne rugueux avec Frankia. L'hypothese de recherche stipulait done que Alnus incana (L.) Moench ssp. rugosa et Alnus viridis (Chaix) DC. ssp. crispa actinorhizes pourraient croitre directement en TS (Chapitre 1) et en coke (Chapitre 2) et que la symbiose avec la bacterie du genre
Frankia ameliorerait le d£veloppement des aulnes croissant dans ces residus. L'hypothese de
recherche a ete investiguee en quantifiant le developpement de l'aulne crispe et de l'aulne rugueux croissant en presence du TS et du coke en conditions gnotobiotiques, en presence et en absence du symbionte Frankia sp. Une etude terrain sur un site contenant du coke sur les sites miniers de la compagnie Syncrude a egalement ete effectuee pour completer 1'investigation, et des donnees de survie et de croissance ont r6colt6es.
CHAPITRE 1
SYMBIOSIS WITH FRANKIA SP. BENEFITS THE ESTABLISHMENT OF ALNUS VIRIDIS SSP. CRISPA AND ALNUS INCANA SSP. RUGOSA IN TAILING SANDS
FROM THE CANADIAN OIL SANDS INDUSTRY
1. R6f6rence de Particle et contribution de l'auteur
Les auteurs ayant contribue a 1'article « Symbiosis with Frankia sp. benefits the establishment of Alnus viridis ssp. crispa and Alnus incana ssp. rugosa in tailing sands from the Canadian oil sands industry » sont: Bissonnette, C, Fahlman, B, Peru, K.M, Khasa, D.P, Greer, C.W, Headley, J.V, Roy, S. La majeure partie des travaux a ete realisee par le premier auteur, qui a mis au point et valide la technique de separation a l'hexane, et a adapte la methode de croissance en hydroponie en conditions gnotobiotiques deja existante au laboratoire de S. Roy pour en faire une methode de croissance en sols. Toutes les manipulations et la redaction ont ete effectuees par le premier auteur, exception faite de 1'analyse des acides naphteniques dans les tissus vegetaux, qui a ete accomplie au laboratoire de J.V. Headley par K. Peru et B. Fahlman. S. Roy a supervise les travaux et a revise le manuscrit.
2. Originality du travail et contribution k la science
Maintes etudes au sujet du comportement de la symbiose entre l'aulne et Frankia en presence de divers contaminants ont deja ete effectuees, dont certaines au sujet du TS (Roy et al., 2007; Quoreshi et al., 2007; Lefran9ois et al., 2010). Cependant, selon nos connaissances, elles se deroulaient toutes sur le terrain, ou en laboratoire en conditions non controtees. L'originalite du present travail reside dans le fait que pour la premiere fois, grace aux conditions gnotobiotiques dans lesquelles les experiences se sont deroulees, il a £te possible de mesurer
l'effet de la symbiose entre deux especes d'aulne et Frankia sp. en excluant l'interference de tout autre microorganisme. La technique experimentale, employee pour evaluer l'effet du TS sur la symbiose, peut etre transposee a tout autre type de sol contamine.
3. Resume de l'article en fran^ais
Au Canada, l'exploitation des sables bitumineux perturbe la foret boreale albertaine et genere des sables residuels (TS), un residu au pH eleve, riche en sels, pauvre en nutriments, et contenant des hydrocarbures phytotoxiques. Jusqu'a present, les methodes utilisees pour revegetaliser ne sont pas suffisamment efficaces pour gerer la quantite croissante de residus engendres par 1'industrie selon les standards gouvernementaux. Les aulnes actinorhizes pourraient etre utilises avec succes dans un programme de revegetalisation direct du TS. Ce sont des plantes pionnieres, qui colonisent naturellement des habitats hostiles et qui ameliorent les proprietes des sols dans lesquels elles croissent. Afin d'evaluer la tolerance des aulnes au stress genere par le TS, le developpement de l'aulne crispe et l'aulne rugueux croissant en TS melange avec du sable sain (de 0 a 100 %), en presence ou en absence du symbionte Frankia sp., a ete determine. II a ete demontre que la survie des aulnes n'est pas alteree par la presence du TS, mais que l'etablissement de la symbiose est moins efficace quand les aulnes croissent en TS pur (100 %). Les aulnes, lorsque actinorhizes, sont tres tolerants a la presence de TS : lorsque croissant en TS pur, leur biomasse seche totale etait 15 fois plus importante que la biomasse des aulnes non-symbiotiques croissant dans les memes conditions. De plus, les aulnes symbiotiques accordent une plus grande proportion de leur biomasse aux parties a^riennes que les aulnes non-nodul6s, peu importe la proportion de TS. Cette proportion etait de 3 a 4 fois plus importante pour l'aulne rugueux et de 1,3 a 2,7 fois plus grande pour l'aulne crisps. En conclusion, une fois la symbiose etablie, les aulnes prosperent et produisent une quantite importante de biomasse, la plantation de jeunes aulnes pr6-inocules avec Frankia sp. semble etre une methode viable de revegetalisation du TS produit lors de l'exploitation des sables bitumineux en Alberta.
4. Manuscrit de l'article
4.1 Abstract
Canada's oil sands extraction disturbs the boreal forest and generates tailing sands (TS), a high pH, high salts, and low nutrient residue which contains phytotoxic hydrocarbons. Efficient methods are needed to revegetate and reclaim impacted landscapes according to governmental standards. Actinorhizal alders might be part of the solution to a successful rehabilitation program. They are early successional species which naturally colonize harsh habitats and improve soil conditions. To evaluate the extent to which alders withstand TS-generated stress, the development of mountain and river alders growing in different proportion of TS mixed with pristine sand (from 0 to 100 %) was assessed in the presence and the absence of their symbiont, Frankia sp. Alder survival was not impaired by the presence of TS, although symbiosis establishment was less efficient when alders grow in pure residue. Actinorhizal alders are very tolerant to TS: in pure TS (100 %), their whole dry biomass was 15 times superior to the dry biomass of non-nodulated alders. Moreover, symbiotic alders allocated a higher proportion of their biomass to aerial parts, regardless of TS concentration, when compared to non-nodulated alders: the proportion was 3 to 4-fold more important for river alders and 1.3 to 2.7-fold higher for mountain alders. Once symbiosis is established, alders thrive and produce an important amount of biomass. Outplanting alder saplings pre-inoculated with Frankia sp. seem to be a valuable method to rehabilitate the TS residues produced by oil sands mining in Alberta.