Axonal growth : modes, molecules and mechanisms
Texte intégral
Figure
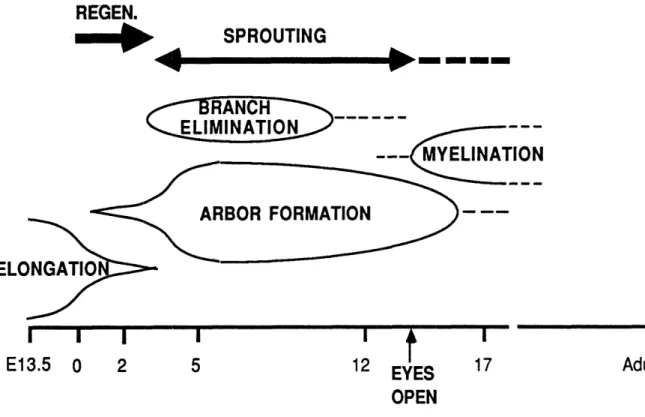
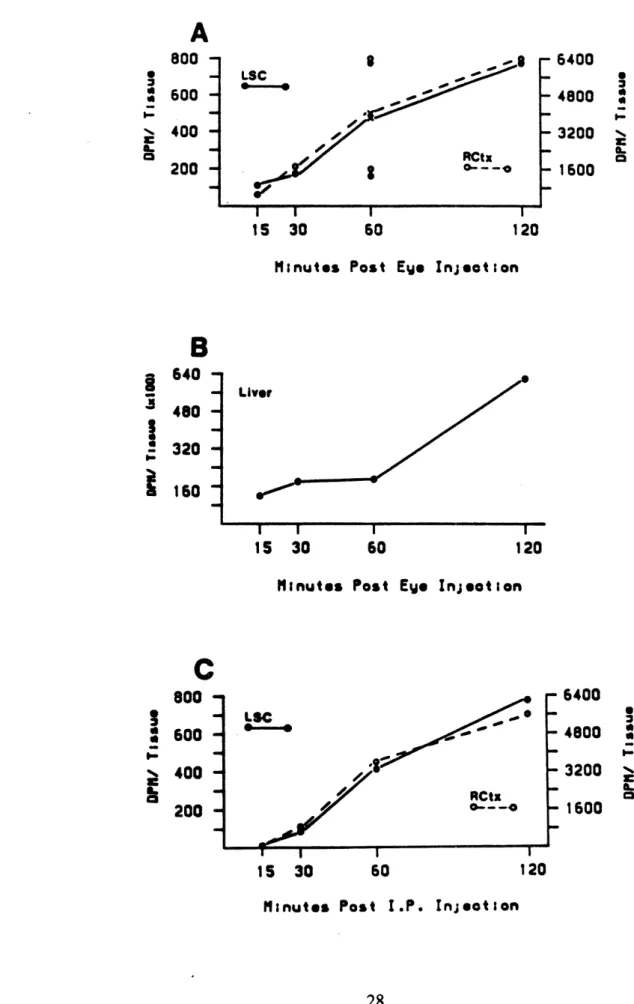
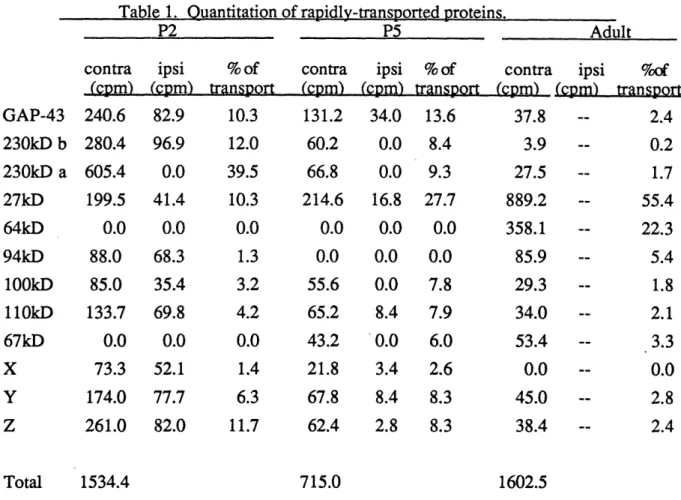
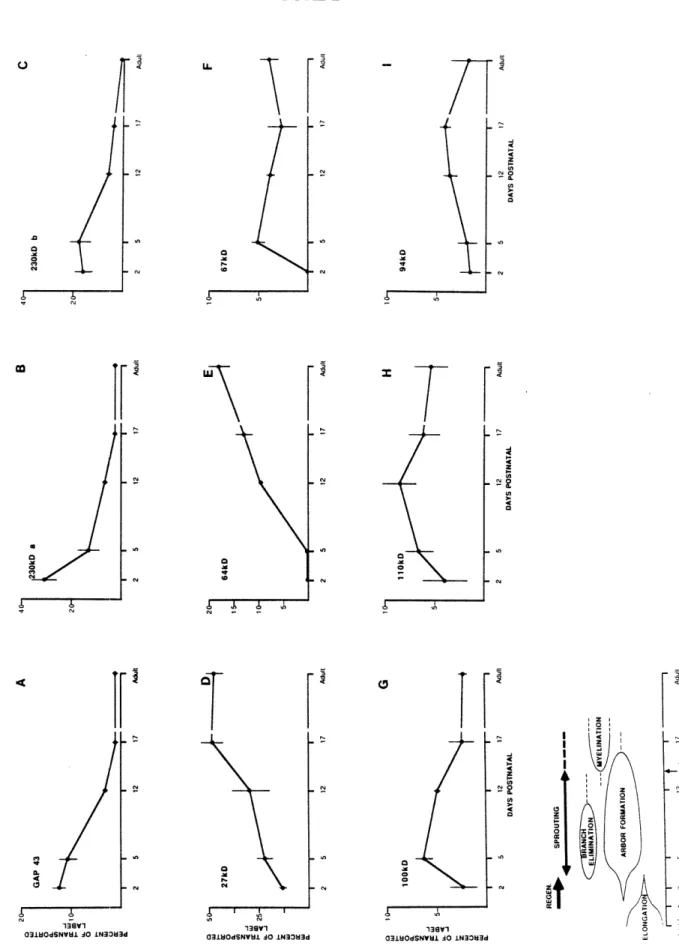
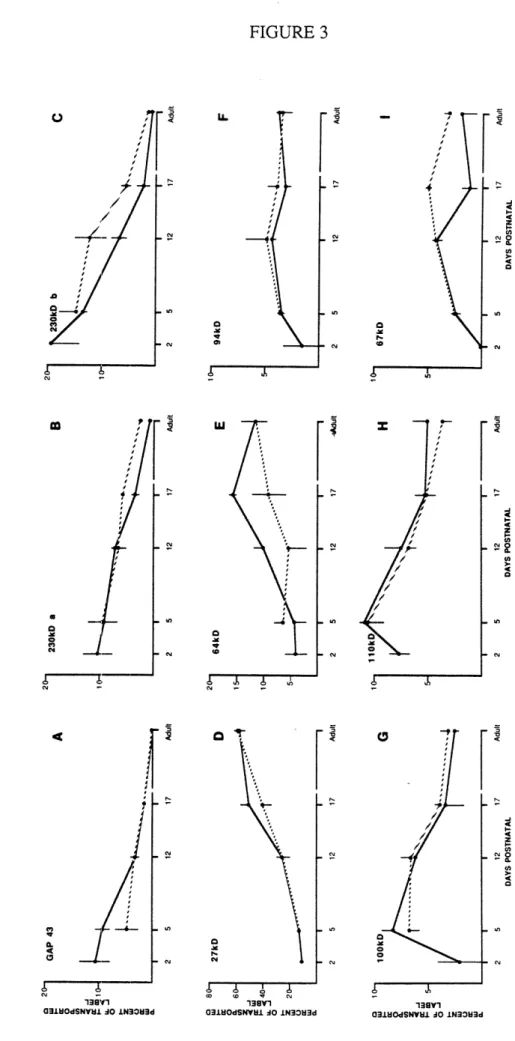
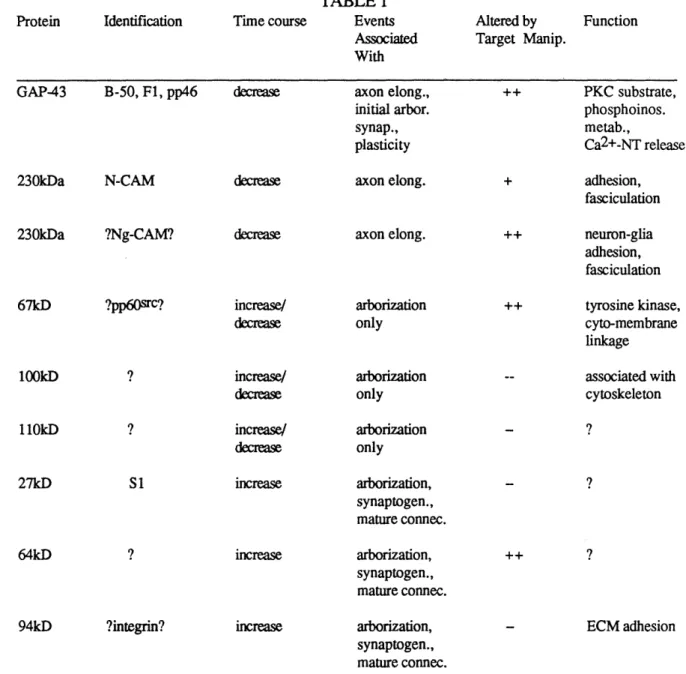
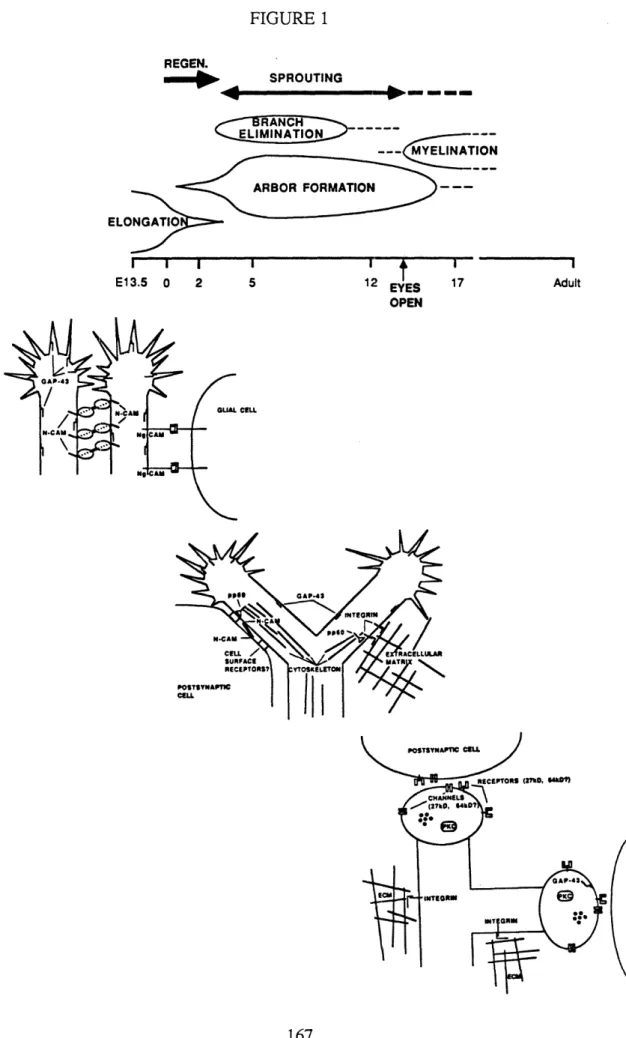
Documents relatifs
Intuitivement, cette couche de a-Si:H est trop fine et poreuse (50 Å) pour imposer des contraintes, et c’est donc au niveau de la couche de µc-Si:H:F que les contraintes
While, for many years, necrosis was considered as a strict equivalent of passive cell death and apoptosis was seen as the sole active form of cell death, nowadays the relevance
Only alternative processes such as heterogeneous nucleation on EPS and/or microbial cell surfaces or increase of saturation by metabolic activity would otherwise allow
Activation of Notch signaling was shown to induce stemness of CSCs, since Notch pathway inhibition –– achieved either by treatment with a -secretase inhibitor or by
II ne doit faire l'objet d'aucun compte rendu ou abstracted or quoted without the agreement of résumé ni d'aucune citation sans l'autorisation de the World Health
Special appreciation goes to Helen Armstrong, Genevieve Becker, Hilary Creed-Kanashiro, Felicity Savage King who were the authors of the WHO/UNICEF training courses on
Small, light monomers (gray online) represent monomers in the crystalline cluster, small dark monomers (red online) represent the attaching chain and bigger monomers (blue
Results showed that words were better reconstructed than pseudowords, evidencing a lexical benefit for words in degraded speech restoration.. We also observed an
