Demographic consequences of heavy metals and persistent organic pollutants in a vulnerable long-lived bird, the wandering albatross
39
0
0
Texte intégral
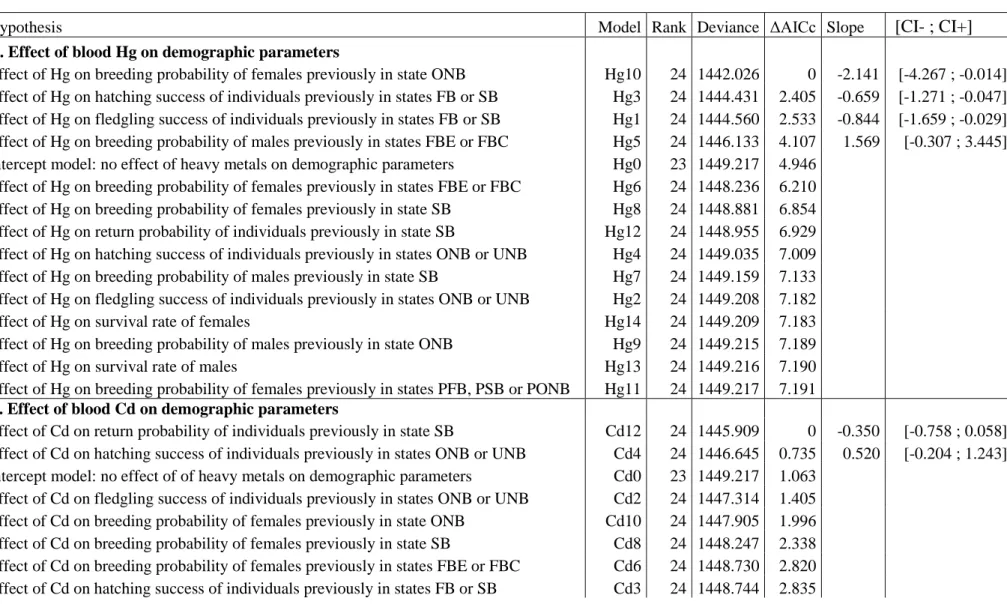
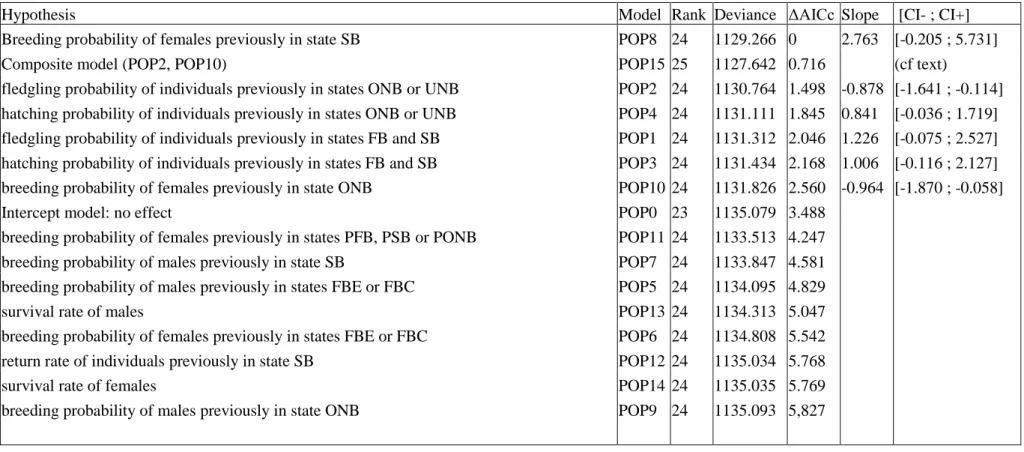
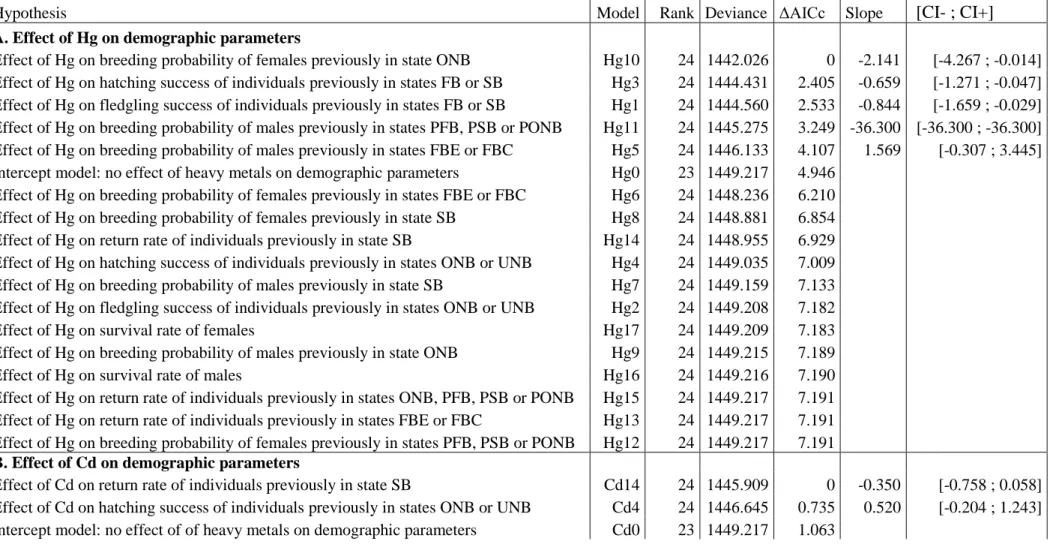
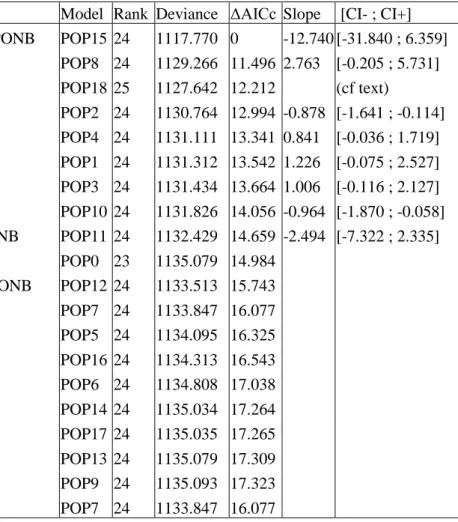
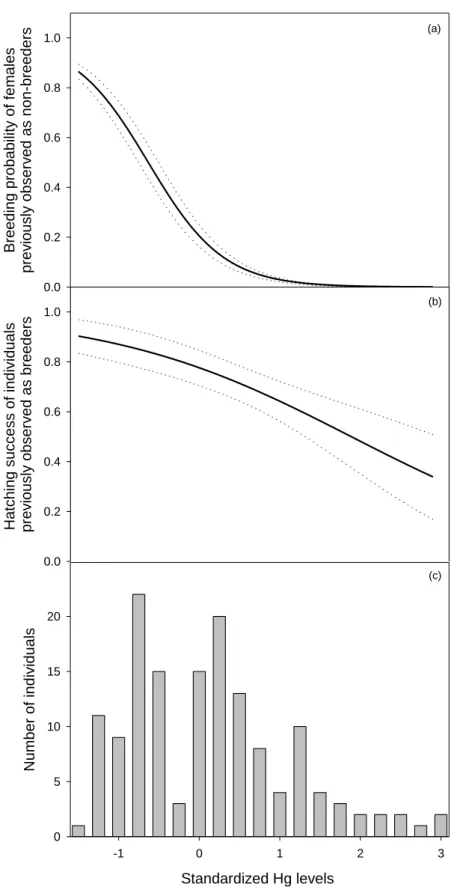
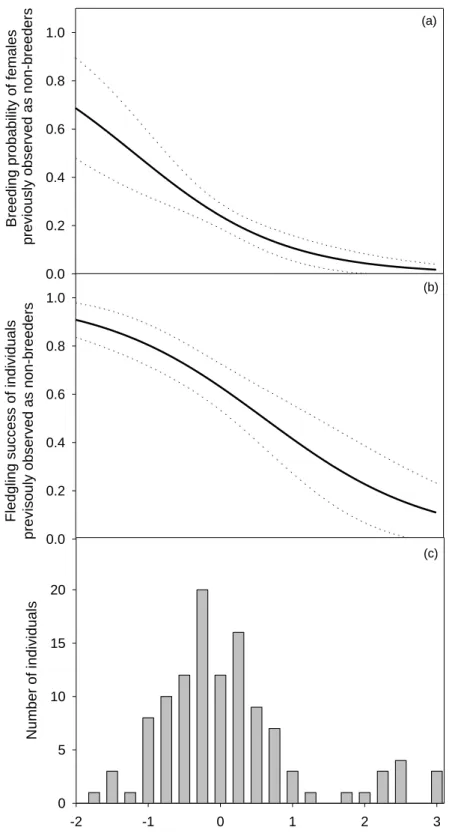
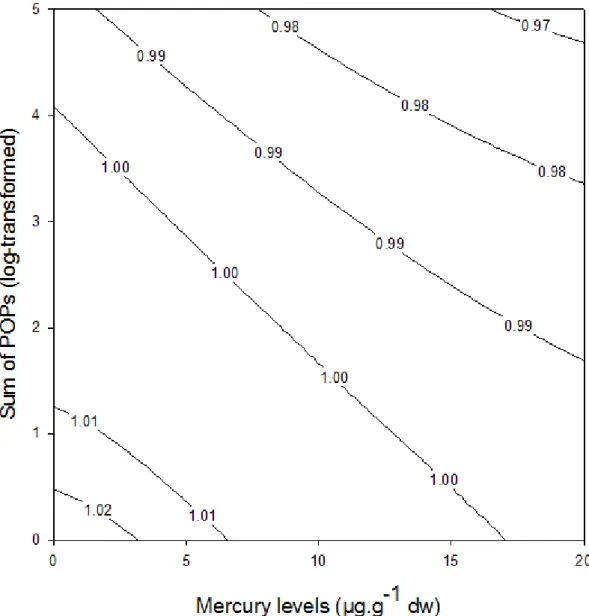
Figure
+4
Documents relatifs
