Reflexive Orienting in Spatial Neglect Is Biased towards Behaviourally Salient Stimuli
9
0
0
Texte intégral
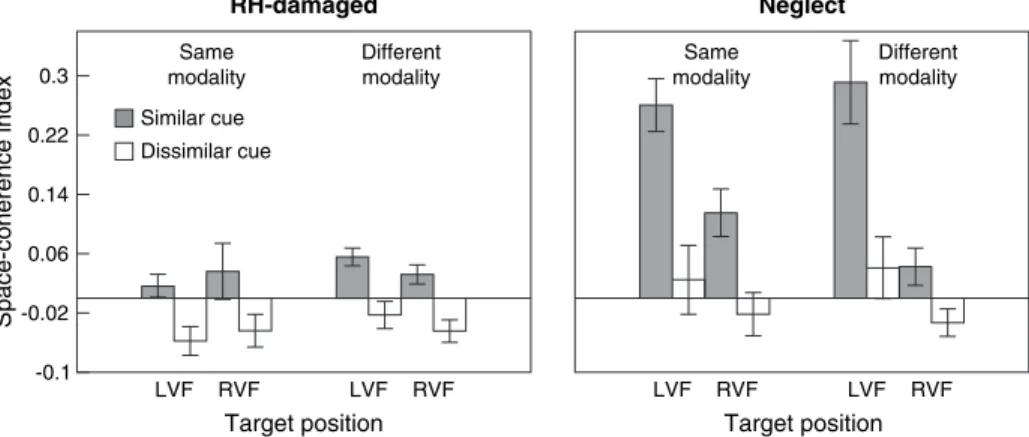
Figure
+2
Documents relatifs