Habitat structure versus food abundance: the importance of sparse vegetation for the common redstart Phoenicurus phoenicurus
11
0
0
Texte intégral
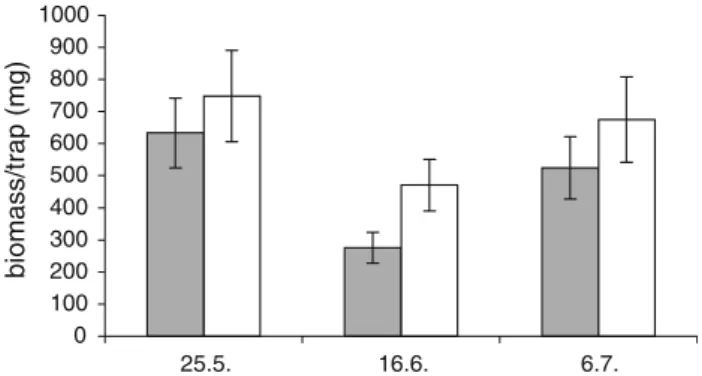
Figure

Documents relatifs