The copper-inducible ComR (YcfQ) repressor regulates expression of ComC (YcfR), which affects copper permeability of the outer membrane of Escherichia coli
11
0
0
Texte intégral
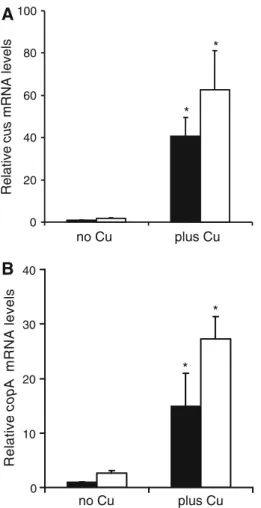
Figure
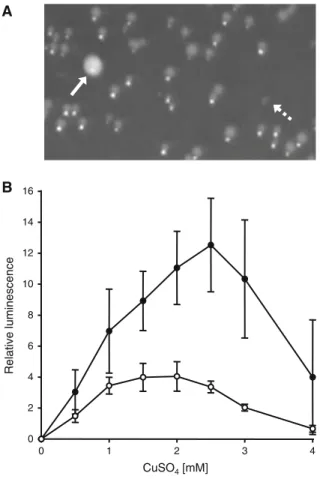
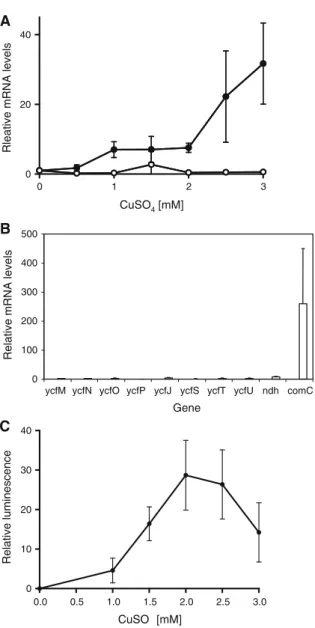
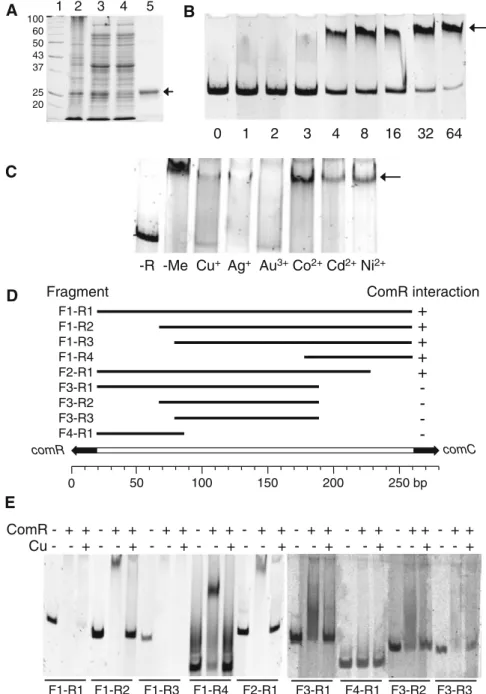
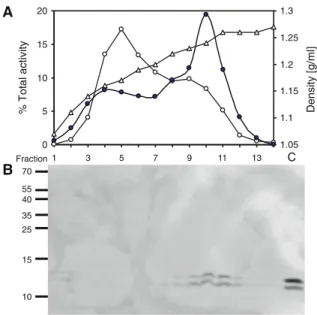
+2
Documents relatifs
