Understanding the limits of animal models as predictors of human biology: lessons learned from the sbv IMPROVER Species Translation Challenge
13
0
0
Texte intégral
Figure

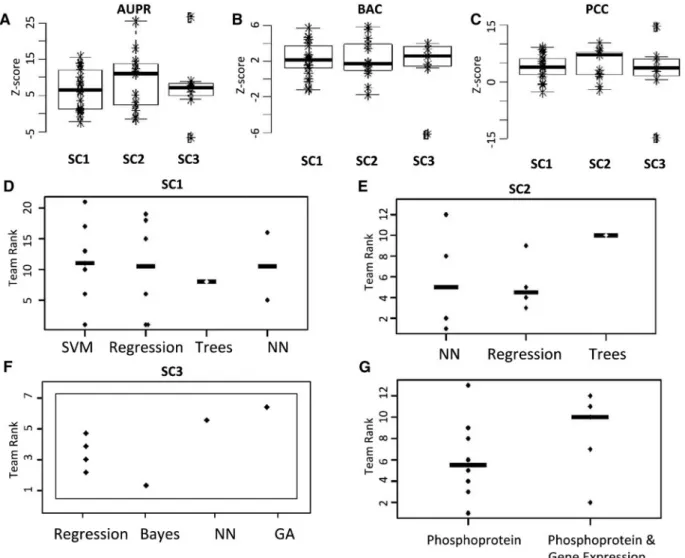
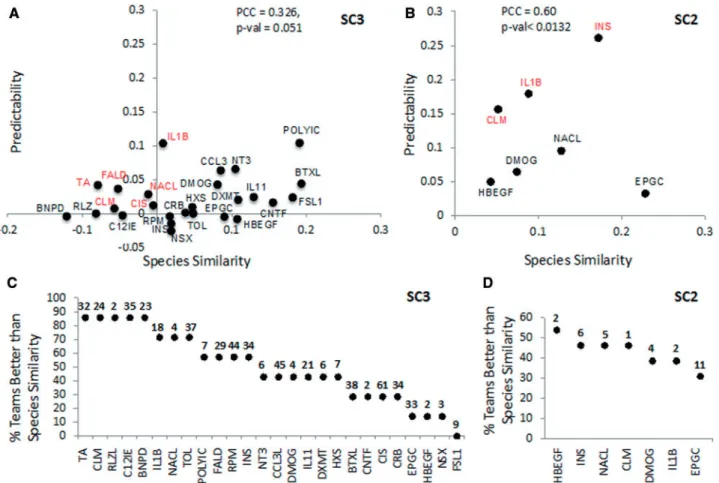
![Fig. 5. Best translated gene sets representative of different pathways. (A) Histogram of the percentage of active gene set/stimulus pairs [560 pairs from 6396 (246 gene sets 26 stimuli)] correctly predicted by N teams](https://thumb-eu.123doks.com/thumbv2/123doknet/14921403.663123/10.918.91.806.95.563/translated-representative-different-pathways-histogram-percentage-correctly-predicted.webp)
Documents relatifs